
Modern Sensor Tools and Techniques for Monitoring, Controlling, and Improving Cell Culture Processes



Abstract
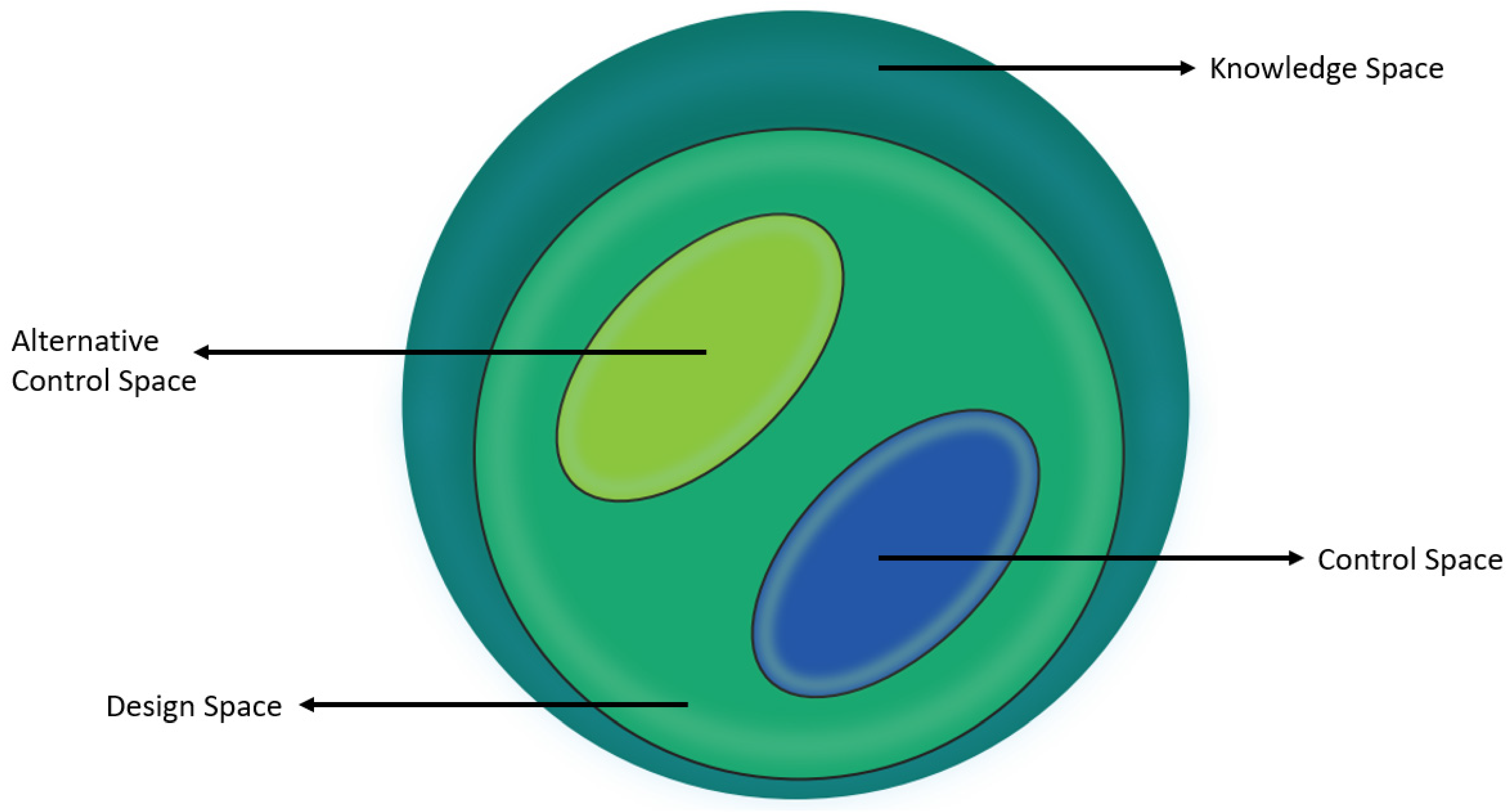
:1. Introduction
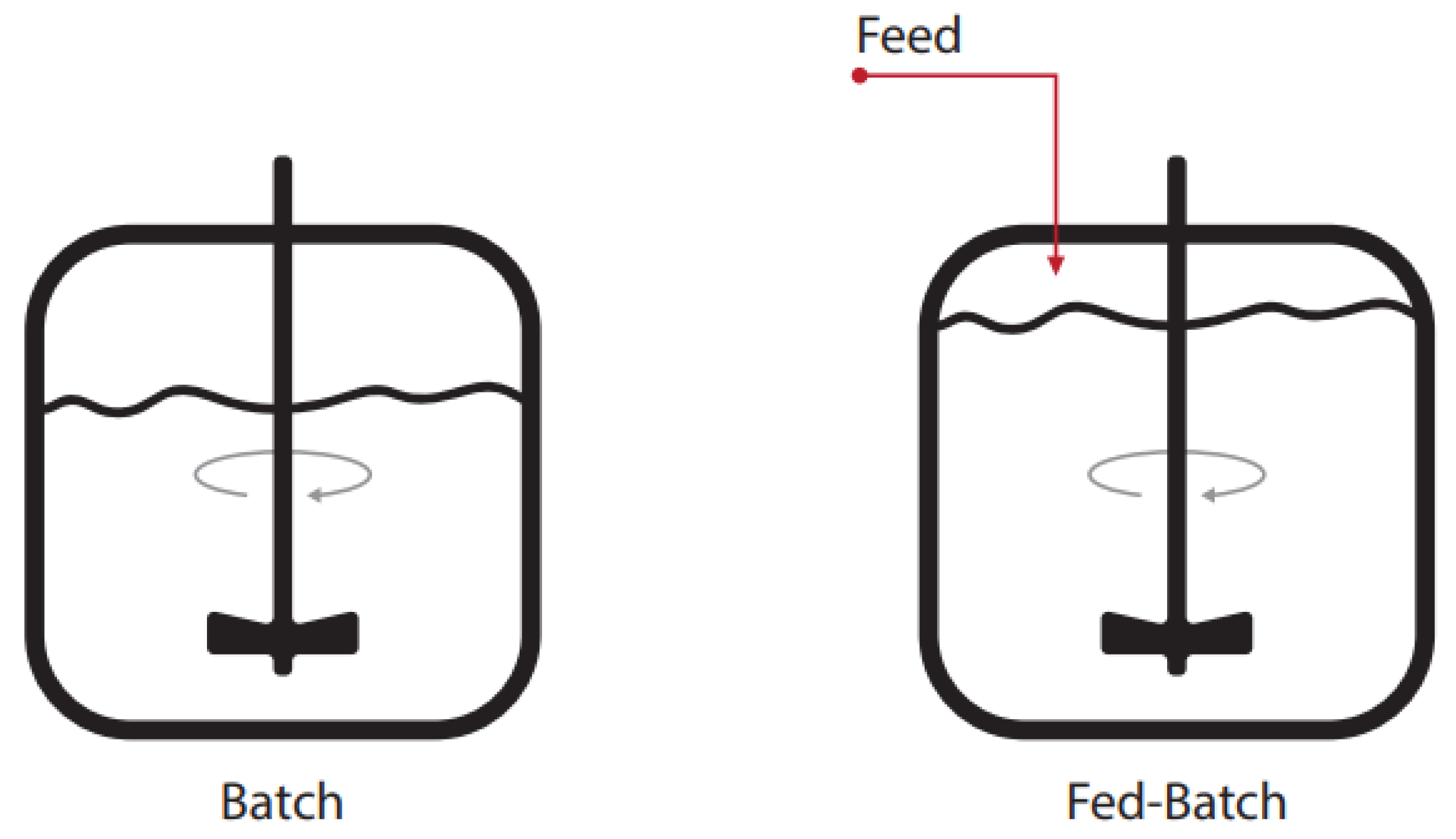
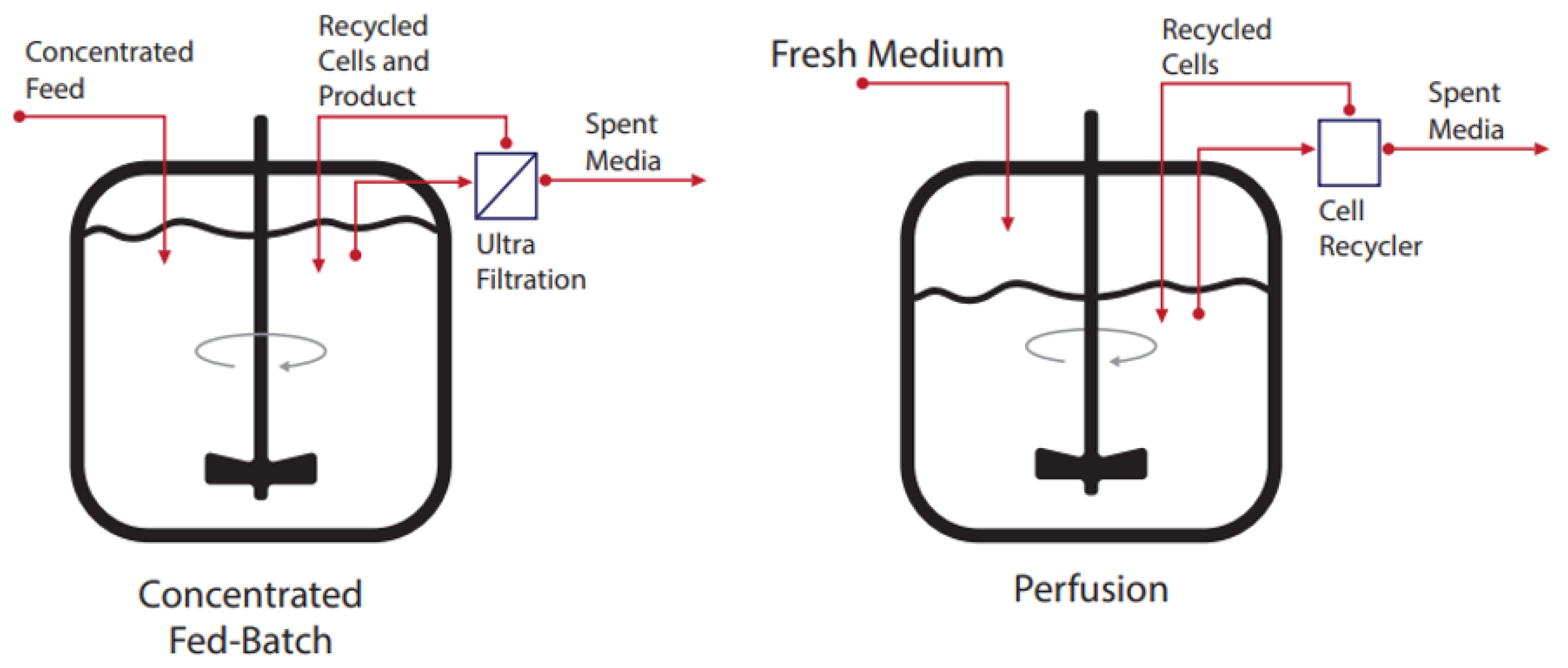
2. Bioreactor Modes of Operation
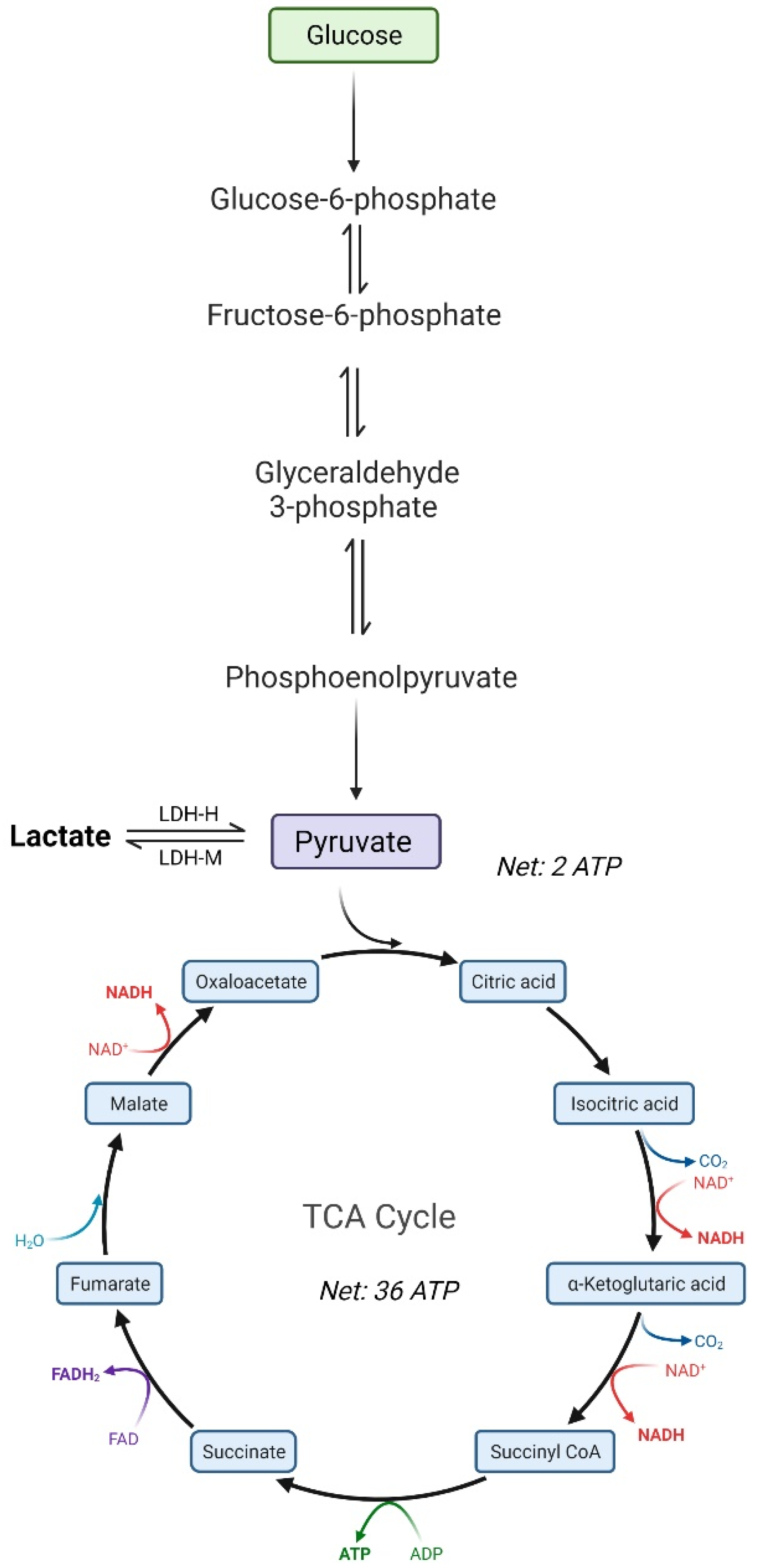
3. Mammalian Cell Metabolism in Culture
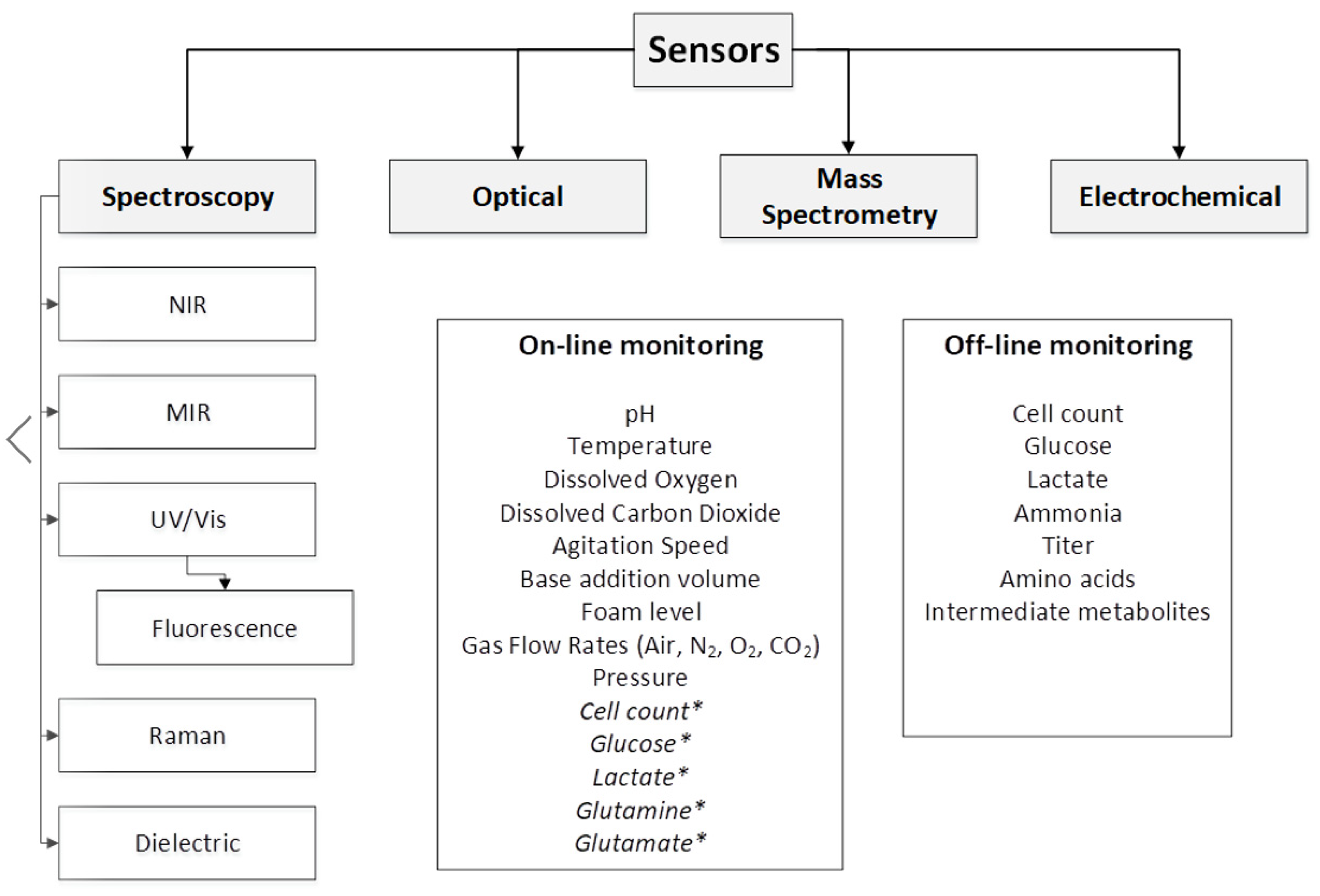
4. Sensor Types and Characteristics
5. Spectroscopy-Based Techniques
5.1. Applications of Near-Infrared (NIR) and Mid-Infrared (MIR) Spectroscopy Techniques
5.2. Applications of Ultraviolet-Visible (UV/Vis) Spectroscopy
5.3. Applications of Raman Spectroscopy
5.4. Applications of Dielectric Spectroscopy
6. Optical Sensing Techniques
7. Mass Spectrometry Techniques
8. Electrochemical Sensing Techniques
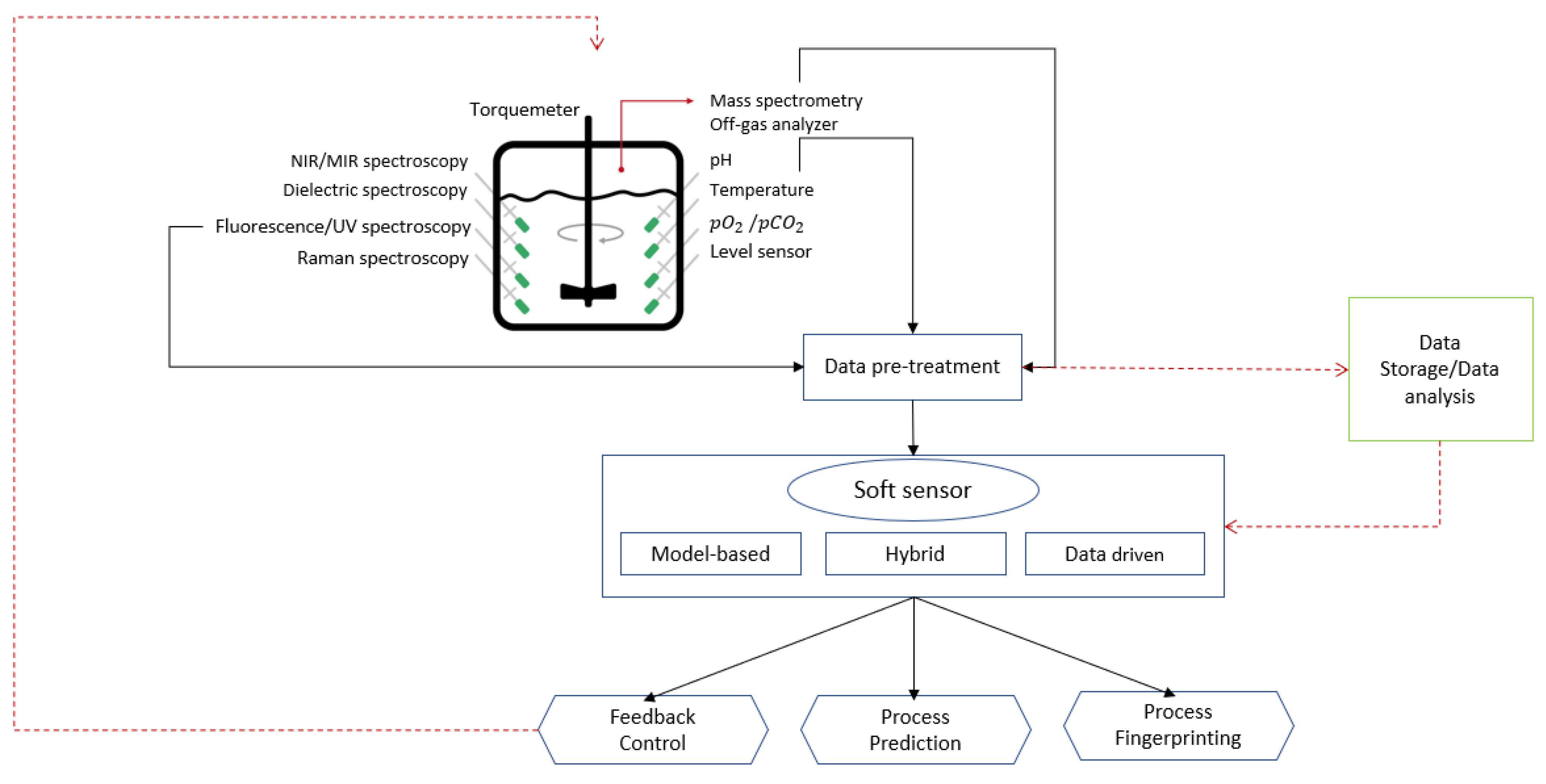
9. Soft Sensors for Cell Culture Monitoring
9.1. Data-Driven Soft Sensors
9.2. Model-Driven Soft Sensors
9.3. Hybrid Models
9.4. Applications of Soft Sensors in Bioprocessing
9.5. Soft Sensor Implementation
9.6. Soft Sensors for Bioprocess Control
10. Concluding Remarks
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Research, G.V. Biotechnology Market Size, Share & Trends Analysis Report by Technology (DNA Sequencing, Nanobiotechnology), by Application (Health, Bioinformatics), by Region, and Segment Forecasts, 2021–2028. Available online: https://www.grandviewresearch.com/industry-analysis/biotechnology-market (accessed on 24 July 2021).
- Intelligence, M. Biopharmaceuticals Market—Growth, Trends, COVID-19 Impact, and Forecasts (2021–2026). Available online: https://www.mordorintelligence.com/industry-reports/global-biopharmaceuticals-market-industry (accessed on 24 July 2021).
- Markets, R.A. Biopharmaceuticals Market by Type and Application—Global Opportunity Analysis and Industry Forecast, 2018–2025. Available online: https://www.researchandmarkets.com/reports/4612776/biopharmaceuticals-market-by-type-and-application (accessed on 24 July 2021).
- Research, G.V. Biosimilars Market Size, Share & Trends Analysis Report by Product, by Application (Oncology, Growth Hormone, Blood Disorders, Chronic & Autoimmune Disorders), by Region, and Segment Forecasts, 2018–2025. Available online: https://markets.businessinsider.com/news/stocks/biosimilars-market-size-worth-61-47-billion-by-2025-cagr-34-2-grand-view-research-inc-1027561345 (accessed on 24 July 2021).
- Intelligence, M. Biosimilars Market—Growth, Trends, COVID-19 Impact, and Forecasts (2021–2026). Available online: https://www.mordorintelligence.com/industry-reports/global-biosimilars-market-industry (accessed on 24 July 2021).
- Li, F.; Vijayasankaran, N.; Shen, A.Y.; Kiss, R.; Amanullah, A. Cell culture processes for monoclonal antibody production. mAbs 2010, 2, 466–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.M.; Hwang, Y.C.; Liu, I.J.; Lee, C.C.; Tsai, H.Z.; Li, H.J.; Wu, H.C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Lustri, J. How to Implement Process Analytical Technology in Pharmaceutical Manufacturing. Available online: https://blog.isa.org/how-to-implement-process-analytical-technology-in-pharmaceutical-manufacturing (accessed on 27 July 2021).
- Uwe Kirschner, R.E.C.; Vangenechten, R.; François, K. Process Analytical Technology: An Industry Perspective. Available online: https://www.europeanpharmaceuticalreview.com/article/3643/process-analytical-technology-pharma-industry/ (accessed on 29 July 2021).
- John, D.O.; George, L.; Reid, I. An Introduction To Process Analytical Technology. Available online: https://www.pharmaceuticalonline.com/doc/an-introduction-to-process-analytical-technology-0001 (accessed on 28 July 2021).
- Mhatre, R.; Rathore, A.S. Quality by Design: An Overview of the Basic Concepts. Qual. By Des. Biopharm. 2009, 1–8. [Google Scholar]
- Rathore, A.S.; Winkle, H. Quality by design for biopharmaceuticals. Nat. Biotechnol. 2009, 27, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Doran, P.M. Bioprocess Engineering Principles; Academic Press: London, UK, 1995. [Google Scholar]
- Shuler, M.L.; Kargı, F.; DeLisa, M. Bioprocess Engineering: Basic Concepts; Prentice Hall: Hoboken, NJ, USA, 2017. [Google Scholar]
- O’Flaherty, R.; Bergin, A.; Flampouri, E.; Mota, L.M.; Obaidi, I.; Quigley, A.; Xie, Y.; Butler, M. Mammalian cell culture for production of recombinant proteins: A review of the critical steps in their biomanufacturing. Biotechnol. Adv. 2020, 43, 107552. [Google Scholar] [CrossRef]
- Wiegmann, V.; Giaka, M.; Martinez, C.B.; Baganz, F. Towards the development of automated fed-batch cell culture processes at microscale. BioTechniques 2019, 67, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wong, H.E.; Goudar, C.T. Upstream process intensification and continuous manufacturing. Curr. Opin. Chem. Eng. 2018, 22, 191–198. [Google Scholar] [CrossRef]
- Bielser, J.-M.; Wolf, M.; Souquet, J.; Broly, H.; Morbidelli, M. Perfusion mammalian cell culture for recombinant protein manufacturing—A critical review. Biotechnol. Adv. 2018, 36, 1328–1340. [Google Scholar] [CrossRef]
- Wlaschin, K.F.; Hu, W.S. Fedbatch culture and dynamic nutrient feeding. Adv. Biochem. Eng. Biotechnol. 2006, 101, 43–74. [Google Scholar] [CrossRef]
- Yang, W.C.; Minkler, D.F.; Kshirsagar, R.; Ryll, T.; Huang, Y.M. Concentrated fed-batch cell culture increases manufacturing capacity without additional volumetric capacity. J. Biotechnol. 2016, 217, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hartley, F.; Walker, T.; Chung, V.; Morten, K. Mechanisms driving the lactate switch in Chinese hamster ovary cells. Biotechnol. Bioeng. 2018, 115, 1890–1903. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Kildegaard, H.F.; Andersen, M.R. Impact of CHO Metabolism on Cell Growth and Protein Production: An Overview of Toxic and Inhibiting Metabolites and Nutrients. Biotechnol. J. 2018, 13, 1700499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galleguillos, S.N.; Ruckerbauer, D.; Gerstl, M.P.; Borth, N.; Hanscho, M.; Zanghellini, J. What can mathematical modelling say about CHO metabolism and protein glycosylation? Comput. Struct. Biotechnol. J. 2017, 15, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Locasale, J.W.; Cantley, L.C. Metabolic flux and the regulation of mammalian cell growth. Cell Metab. 2011, 14, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Mulukutla, B.C.; Khan, S.; Lange, A.; Hu, W.-S. Glucose metabolism in mammalian cell culture: New insights for tweaking vintage pathways. Trends Biotechnol. 2010, 28, 476–484. [Google Scholar] [CrossRef]
- Mulukutla, B.C.; Gramer, M.; Hu, W.-S. On metabolic shift to lactate consumption in fed-batch culture of mammalian cells. Metab. Eng. 2012, 14, 138–149. [Google Scholar] [CrossRef]
- Ozturk, S.S.; Riley, M.R.; Palsson, B.O. Effects of ammonia and lactate on hybridoma growth, metabolism, and antibody production. Biotechnol. Bioeng. 1992, 39, 418–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagari, F.; Jordan, M.; Stettler, M.; Broly, H.; Wurm, F.M. Lactate metabolism shift in CHO cell culture: The role of mitochondrial oxidative activity. New Biotechnol. 2013, 30, 238–245. [Google Scholar] [CrossRef]
- Fan, Y.; Jimenez del Val, I.; Müller, C.; Wagtberg Sen, J.; Rasmussen, S.K.; Kontoravdi, C.; Weilguny, D.; Andersen, M.R. Amino acid and glucose metabolism in fed-batch CHO cell culture affects antibody production and glycosylation. Biotechnol. Bioeng. 2015, 112, 521–535. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.; Marison, I.W.; von Stockar, U. The importance of ammonia in mammalian cell culture. J. Biotechnol. 1996, 46, 161–185. [Google Scholar] [CrossRef]
- Martinelle, K.; Häggström, L. Effects of NH4+ and K+ on the energy metabolism in Sp2/0-Ag14 myeloma cells. Cytotechnology 1999, 29, 45–53. [Google Scholar] [CrossRef]
- Duarte, T.M.; Carinhas, N.; Barreiro, L.C.; Carrondo, M.J.T.; Alves, P.M.; Teixeira, A.P. Metabolic responses of CHO cells to limitation of key amino acids. Biotechnol. Bioeng. 2014, 111, 2095–2106. [Google Scholar] [CrossRef]
- Mohmad-Saberi, S.E.; Hashim, Y.Z.; Mel, M.; Amid, A.; Ahmad-Raus, R.; Packeer-Mohamed, V. Metabolomics profiling of extracellular metabolites in CHO-K1 cells cultured in different types of growth media. Cytotechnology 2013, 65, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Mulukutla, B.C.; Kale, J.; Kalomeris, T.; Jacobs, M.; Hiller, G.W. Identification and control of novel growth inhibitors in fed-batch cultures of Chinese hamster ovary cells. Biotechnol. Bioeng. 2017, 114, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Templeton, N.; Dean, J.; Reddy, P.; Young, J.D. Peak antibody production is associated with increased oxidative metabolism in an industrially relevant fed-batch CHO cell culture. Biotechnol. Bioeng. 2013, 110, 2013–2024. [Google Scholar] [CrossRef]
- Mulukutla, B.C.; Mitchell, J.; Geoffroy, P.; Harrington, C.; Krishnan, M.; Kalomeris, T.; Morris, C.; Zhang, L.; Pegman, P.; Hiller, G.W. Metabolic engineering of Chinese hamster ovary cells towards reduced biosynthesis and accumulation of novel growth inhibitors in fed-batch cultures. Metab. Eng. 2019, 54, 54–68. [Google Scholar] [CrossRef]
- O’Mara, P.; Farrell, A.; Bones, J.; Twomey, K. Staying alive! Sensors used for monitoring cell health in bioreactors. Talanta 2018, 176, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Hoshan, L.; Jiang, R.; Moroney, J.; Bui, A.; Zhang, X.; Hang, T.-C.; Xu, S. Effective bioreactor pH control using only sparging gases. Biotechnol. Prog. 2019, 35, e2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Z.; Lewis, A.M.; Borys, M.C.; Li, Z.J. A carbon dioxide stripping model for mammalian cell culture in manufacturing scale bioreactors. Biotechnol. Bioeng. 2017, 114, 1184–1194. [Google Scholar] [CrossRef]
- Meghrous, J.; Khramtsov, N.; Buckland, B.C.; Cox, M.M.J.; Palomares, L.A.; Srivastava, I.K. Dissolved carbon dioxide determines the productivity of a recombinant hemagglutinin component of an influenza vaccine produced by insect cells. Biotechnol. Bioeng. 2015, 112, 2267–2275. [Google Scholar] [CrossRef]
- Biechele, P.; Busse, C.; Solle, D.; Scheper, T.; Reardon, K. Sensor systems for bioprocess monitoring. Eng. Life Sci. 2015, 15, 469–488. [Google Scholar] [CrossRef]
- Zhao, L.; Fu, H.Y.; Zhou, W.; Hu, W.S. Advances in process monitoring tools for cell culture bioprocesses. Eng. Life Sci. 2015, 15, 459–468. [Google Scholar] [CrossRef]
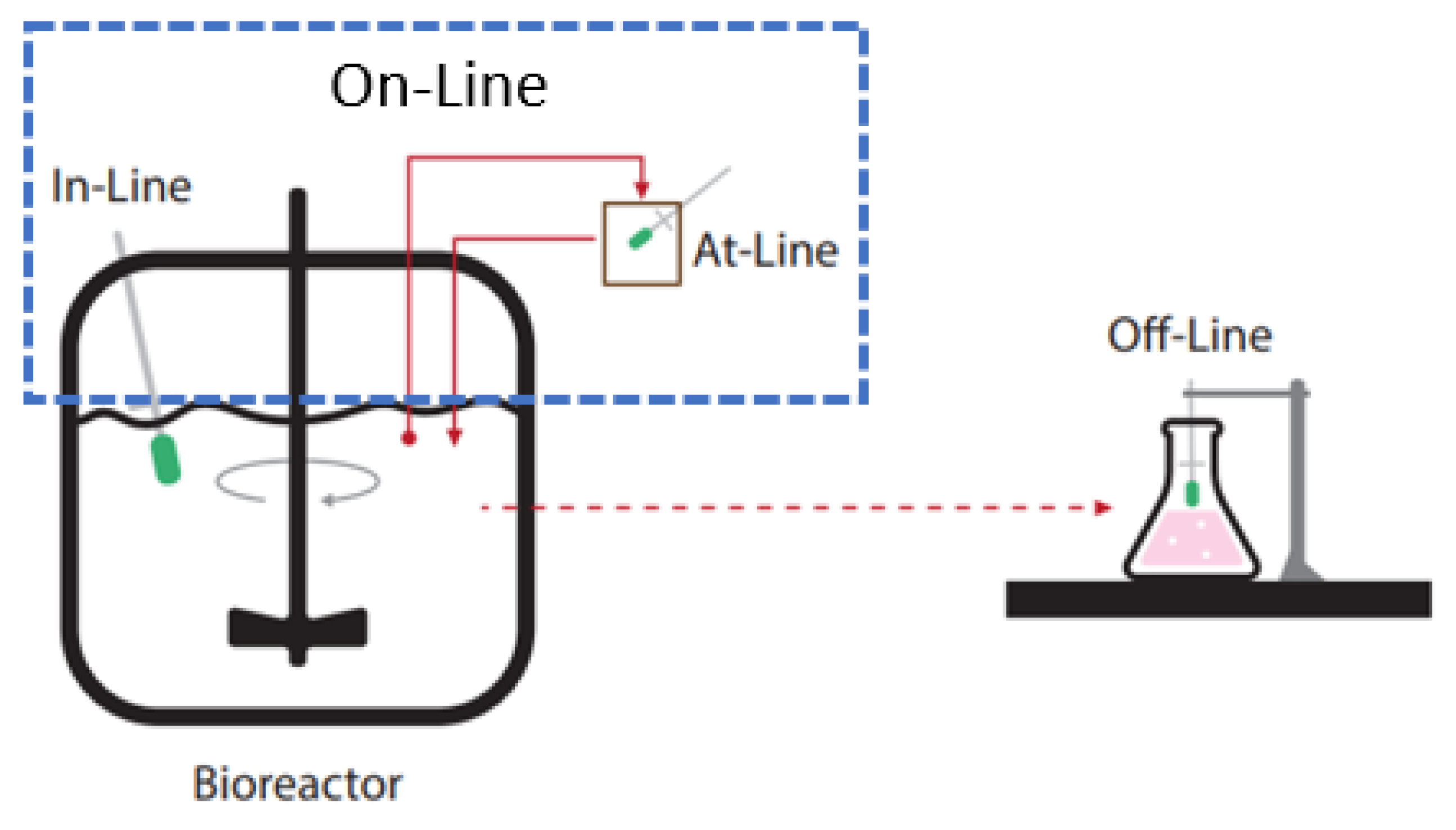
- Company, H. Bioprocess Monitoring and Control (Off-Line, At-Line, On-Line, In-Line/In-Situ). Available online: https://www.hamiltoncompany.com/process-analytics/process-analytical-technology/bioprocess-monitoring-and-control (accessed on 27 July 2021).
- Gargalo, C.L.; Udugama, I.; Pontius, K.; Lopez, P.C.; Nielsen, R.F.; Hasanzadeh, A.; Mansouri, S.S.; Bayer, C.; Junicke, H.; Gernaey, K.V. Towards smart biomanufacturing: A perspective on recent developments in industrial measurement and monitoring technologies for bio-based production processes. J. Ind. Microbiol. Biotechnol. 2020, 47, 947–964. [Google Scholar] [CrossRef]
- Hub, E. Introduction to Sensors and Transducers. Available online: https://www.electronicshub.org/sensors-and-transducers-introduction/ (accessed on 30 July 2021).
- Bhalla, N.; Jolly, P.; Formisano, N.; Estrela, P. Introduction to biosensors. Essays Biochem. 2016, 60, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kisaalita, W.S. Biosensor standards requirements. Biosens. Bioelectron. 1992, 7, 613–620. [Google Scholar] [CrossRef]
- Steinwedel, T.; Dahlmann, K.; Solle, D.; Scheper, T.; Reardon, K.F.; Lammers, F. Sensors for disposable bioreactor systems. In Single-Use Technology in Biopharmaceutical Manufacture; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2019; pp. 69–82. [Google Scholar]
- Busse, C.; Biechele, P.; de Vries, I.; Reardon, K.F.; Solle, D.; Scheper, T. Sensors for disposable bioreactors. Eng. Life Sci. 2017, 17, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Clavaud, M.; Roggo, Y.; von Daeniken, R.; Liebler, A.; Schwabe, J.O. Chemometrics and in-line near infrared spectroscopic monitoring of a biopharmaceutical Chinese hamster ovary cell culture: Prediction of multiple cultivation variables. Talanta 2013, 111, 28–38. [Google Scholar] [CrossRef]
- Sandor, M.; Rüdinger, F.; Bienert, R.; Grimm, C.; Solle, D.; Scheper, T. Comparative study of non-invasive monitoring via infrared spectroscopy for mammalian cell cultivations. J. Biotechnol. 2013, 168, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.R.; Crider, H.M.; Nite, M.E.; Garcia, R.A.; Woo, J.; Wegge, R.M. Simultaneous measurement of 19 components in serum-containing animal cell culture media by fourier transform near-infrared spectroscopy. Biotechnol. Prog. 2001, 17, 376–378. [Google Scholar] [CrossRef]
- Hakemeyer, C.; Strauss, U.; Werz, S.; Jose, G.E.; Folque, F.; Menezes, J.C. At-line NIR spectroscopy as effective PAT monitoring technique in Mab cultivations during process development and manufacturing. Talanta 2012, 90, 12–21. [Google Scholar] [CrossRef]
- Capito, F.; Zimmer, A.; Skudas, R. Mid-infrared spectroscopy-based analysis of mammalian cell culture parameters. Biotechnol. Prog. 2015, 31, 578–584. [Google Scholar] [CrossRef]
- Rosa, F.O.P.; Cunha, B.; Carmelo, J.G.; Fernandes-Platzgummer, A.; da Silva, C.L.; Calado, C.R.C. Mid-infrared spectroscopy: A groundbreaking tool for monitoring mammalian cells processes. In Proceedings of the 2017 IEEE 5th Portuguese Meeting on Bioengineering (ENBENG), Coimbra, Portugal, 16–18 February 2017; pp. 1–6. [Google Scholar]
- Hansen, S.K.; Jamali, B.; Hubbuch, J. Selective high throughput protein quantification based on UV absorption spectra. Biotechnol. Bioeng. 2013, 110, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.B.; Leme, J.; Caricati, C.P.; Tonso, A.; Fernández Núñez, E.G.; Rocha, J.C. Artificial neural network associated to UV/Vis spectroscopy for monitoring bioreactions in biopharmaceutical processes. Bioprocess Biosyst. Eng. 2015, 38, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Drieschner, T.; Ostertag, E.; Boldrini, B.; Lorenz, A.; Brecht, M.; Rebner, K. Direct optical detection of cell density and viability of mammalian cells by means of UV/VIS spectroscopy. Anal. Bioanal. Chem. 2020, 412, 3359–3371. [Google Scholar] [CrossRef]
- Leme, J.; Fernández Núñez, E.G.; de Almeida Parizotto, L.; Chagas, W.A.; dos Santos, E.S.; Tojeira Prestia Caricati, A.; de Rezende, A.G.; da Costa, B.L.V.; Ventini Monteiro, D.C.; Lopes Boldorini, V.L.; et al. A multivariate calibration procedure for UV/VIS spectrometric monitoring of BHK-21 cell metabolism and growth. Biotechnol. Prog. 2014, 30, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ebel, B.; Chauchard, F.; Guédon, E.; Marc, A. Parallel comparison of in situ Raman and NIR spectroscopies to simultaneously measure multiple variables toward real-time monitoring of CHO cell bioreactor cultures. Biochem. Eng. J. 2018, 137, 205–213. [Google Scholar] [CrossRef]
- Teixeira, A.P.; Portugal, C.A.; Carinhas, N.; Dias, J.M.; Crespo, J.P.; Alves, P.M.; Carrondo, M.J.; Oliveira, R. In situ 2D fluorometry and chemometric monitoring of mammalian cell cultures. Biotechnol. Bioeng. 2009, 102, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.P.; Duarte, T.M.; Oliveira, R.; Carrondo, M.J.T.; Alves, P.M. High-throughput analysis of animal cell cultures using two-dimensional fluorometry. J. Biotechnol. 2011, 151, 255–260. [Google Scholar] [CrossRef]
- Teixeira, A.P.; Duarte, T.M.; Carrondo, M.J.T.; Alves, P.M. Synchronous fluorescence spectroscopy as a novel tool to enable PAT applications in bioprocesses. Biotechnol. Bioeng. 2011, 108, 1852–1861. [Google Scholar] [CrossRef] [PubMed]
- Claßen, J.; Graf, A.; Aupert, F.; Solle, D.; Höhse, M.; Scheper, T. A novel LED-based 2D-fluorescence spectroscopy system for in-line bioprocess monitoring of Chinese hamster ovary cell cultivations—Part II. Eng. Life Sci. 2019, 19, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Esmonde-White, K.A.; Cuellar, M.; Uerpmann, C.; Lenain, B.; Lewis, I.R. Raman spectroscopy as a process analytical technology for pharmaceutical manufacturing and bioprocessing. Anal. Bioanal. Chem. 2017, 409, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Abu-Absi, N.R.; Kenty, B.M.; Cuellar, M.E.; Borys, M.C.; Sakhamuri, S.; Strachan, D.J.; Hausladen, M.C.; Li, Z.J. Real time monitoring of multiple parameters in mammalian cell culture bioreactors using an in-line Raman spectroscopy probe. Biotechnol. Bioeng. 2011, 108, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ford, J.; Mann, C.; Vickers, T.; Brackett, J.; Cousineau, K.; Robey, W. Raman Measurement of Glucose in Bioreactor Materials; SPIE: Bellingham, WA, USA, 1997; Volume 2976. [Google Scholar]
- Whelan, J.; Craven, S.; Glennon, B. In situ Raman spectroscopy for simultaneous monitoring of multiple process parameters in mammalian cell culture bioreactors. Biotechnol. Prog. 2012, 28, 1355–1362. [Google Scholar] [CrossRef]
- Berry, B.; Moretto, J.; Matthews, T.; Smelko, J.; Wiltberger, K. Cross-scale predictive modeling of CHO cell culture growth and metabolites using Raman spectroscopy and multivariate analysis. Biotechnol. Prog. 2015, 31, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ray, B.H.; Leister, K.J.; Ryder, A.G. Performance monitoring of a mammalian cell based bioprocess using Raman spectroscopy. Anal. Chim. Acta 2013, 796, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, T.A.; Hadley, B.C.; Hilliard, W.; Jaques, C.; Mason, C. Development of generic Raman models for a GS-KOTM CHO platform process. Biotechnol. Prog. 2018, 34, 730–737. [Google Scholar] [CrossRef]
- Santos, R.M.; Kessler, J.M.; Salou, P.; Menezes, J.C.; Peinado, A. Monitoring mAb cultivations with in-situ raman spectroscopy: The influence of spectral selectivity on calibration models and industrial use as reliable PAT tool. Biotechnol. Prog. 2018, 34, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Matthews, T.E.; Berry, B.N.; Smelko, J.; Moretto, J.; Moore, B.; Wiltberger, K. Closed loop control of lactate concentration in mammalian cell culture by Raman spectroscopy leads to improved cell density, viability, and biopharmaceutical protein production. Biotechnol. Bioeng. 2016, 113, 2416–2424. [Google Scholar] [CrossRef]
- Berry, B.N.; Dobrowsky, T.M.; Timson, R.C.; Kshirsagar, R.; Ryll, T.; Wiltberger, K. Quick generation of Raman spectroscopy based in-process glucose control to influence biopharmaceutical protein product quality during mammalian cell culture. Biotechnol. Prog. 2016, 32, 224–234. [Google Scholar] [CrossRef]
- Párta, L.; Zalai, D.; Borbély, S.; Putics, Á. Application of dielectric spectroscopy for monitoring high cell density in monoclonal antibody producing CHO cell cultivations. Bioprocess Biosyst. Eng. 2014, 37, 311–323. [Google Scholar] [CrossRef]
- Justice, C.; Brix, A.; Freimark, D.; Kraume, M.; Pfromm, P.; Eichenmueller, B.; Czermak, P. Process control in cell culture technology using dielectric spectroscopy. Biotechnol. Adv. 2011, 29, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Zeiser, A.; Bédard, C.; Voyer, R.; Jardin, B.; Tom, R.; Kamen, A.A. On-line monitoring of the progress of infection in Sf-9 insect cell cultures using relative permittivity measurements. Biotechnol. Bioeng. 1999, 63, 122–126. [Google Scholar] [CrossRef]
- Metze, S.; Blioch, S.; Matuszczyk, J.; Greller, G.; Grimm, C.; Scholz, J.; Hoehse, M. Multivariate data analysis of capacitance frequency scanning for online monitoring of viable cell concentrations in small-scale bioreactors. Anal. Bioanal. Chem. 2020, 412, 2089–2102. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.; Sanford, R.; Zhang, A. Case study: The characterization and implementation of dielectric spectroscopy (biocapacitance) for process control in a commercial GMP CHO manufacturing process. Biotechnol. Prog. 2019, 35, e2782. [Google Scholar] [CrossRef]
- Lu, F.; Toh, P.C.; Burnett, I.; Li, F.; Hudson, T.; Amanullah, A.; Li, J. Automated dynamic fed-batch process and media optimization for high productivity cell culture process development. Biotechnol. Bioeng. 2013, 110, 191–205. [Google Scholar] [CrossRef]
- Konakovsky, V.; Clemens, C.; Müller, M.M.; Bechmann, J.; Berger, M.; Schlatter, S.; Herwig, C. Metabolic Control in Mammalian Fed-Batch Cell Cultures for Reduced Lactic Acid Accumulation and Improved Process Robustness. Bioengineering 2016, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Konakovsky, V.; Yagtu, A.C.; Clemens, C.; Müller, M.M.; Berger, M.; Schlatter, S.; Herwig, C. Universal Capacitance Model for Real-Time Biomass in Cell Culture. Sensors 2015, 15, 22128–22150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casablancas, A.; Gámez, X.; Lecina, M.; Solà, C.; Cairó, J.J.; Gòdia, F. Comparison of control strategies for fed-batch culture of hybridoma cells based on on-line monitoring of oxygen uptake rate, optical cell density and glucose concentration. J. Chem. Technol. Biotechnol. 2013, 88, 1680–1689. [Google Scholar] [CrossRef]
- Goldrick, S.; Lee, K.; Spencer, C.; Holmes, W.; Kuiper, M.; Turner, R.; Farid, S.S. On-Line Control of Glucose Concentration in High-Yielding Mammalian Cell Cultures Enabled Through Oxygen Transfer Rate Measurements. Biotechnol. J. 2018, 13, e1700607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamen, A.A.; Bédard, C.; Tom, R.; Perret, S.; Jardin, B. On-line monitoring of respiration in recombinant-baculovirus infected and uninfected insect cell bioreactor cultures. Biotechnol. Bioeng. 1996, 50, 36–48. [Google Scholar] [CrossRef]
- Pappenreiter, M.; Sissolak, B.; Sommeregger, W.; Striedner, G. Oxygen Uptake Rate Soft-Sensing via Dynamic kLa Computation: Cell Volume and Metabolic Transition Prediction in Mammalian Bioprocesses. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Lam, H.; Kostov, Y. Optical instrumentation for bioprocess monitoring. Adv. Biochem. Eng. Biotechnol. 2009, 116, 125–142. [Google Scholar] [CrossRef]
- Becker, M.; Junghans, L.; Teleki, A.; Bechmann, J.; Takors, R. The Less the Better: How Suppressed Base Addition Boosts Production of Monoclonal Antibodies with Chinese Hamster Ovary Cells. Front. Bioeng. Biotechnol. 2019, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Kenty, B.M.; Li, Z.J.; Lee, S.S. Scale-up analysis for a CHO cell culture process in large-scale bioreactors. Biotechnol. Bioeng. 2009, 103, 733–746. [Google Scholar] [CrossRef]
- Doi, T.; Kajihara, H.; Chuman, Y.; Kuwae, S.; Kaminagayoshi, T.; Omasa, T. Development of a scale-up strategy for Chinese hamster ovary cell culture processes using the kLa ratio as a direct indicator of gas stripping conditions. Biotechnol. Prog. 2020, 36, e3000. [Google Scholar] [CrossRef]
- Heinzle, E. Present and potential applications of mass spectrometry for bioprocess research and control. J. Biotechnol. 1992, 25, 81–114. [Google Scholar] [CrossRef]
- Lyubarskaya, Y.; Kobayashi, K.; Swann, P. Application of mass spectrometry to facilitate advanced process controls of biopharmaceutical manufacture. Pharm. Bioprocess. 2015, 3, 313–321. [Google Scholar] [CrossRef]
- Behrendt, U.; Koch, S.; Gooch, D.D.; Steegmans, U.; Comer, M.J. Mass spectrometry: A tool for on-line monitoring of animal cell cultures. Cytotechnology 1994, 14, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Goh, H.-Y.; Sulu, M.; Alosert, H.; Lewis, G.L.; Josland, G.D.; Merriman, D.E. Applications of off-gas mass spectrometry in fed-batch mammalian cell culture. Bioprocess Biosyst. Eng. 2020, 43, 483–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floris, P.; Dorival-García, N.; Lewis, G.; Josland, G.; Merriman, D.; Bones, J. Real-time characterization of mammalian cell culture bioprocesses by magnetic sector MS. Anal. Methods 2020, 12, 5601–5612. [Google Scholar] [CrossRef]
- Rogers, R.S.; Nightlinger, N.S.; Livingston, B.; Campbell, P.; Bailey, R.; Balland, A. Development of a quantitative mass spectrometry multi-attribute method for characterization, quality control testing and disposition of biologics. mAbs 2015, 7, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Luchner, M.; Gutmann, R.; Bayer, K.; Dunkl, J.; Hansel, A.; Herbig, J.; Singer, W.; Strobl, F.; Winkler, K.; Striedner, G. Implementation of proton transfer reaction-mass spectrometry (PTR-MS) for advanced bioprocess monitoring. Biotechnol. Bioeng. 2012, 109, 3059–3069. [Google Scholar] [CrossRef] [PubMed]
- Schmidberger, T.; Gutmann, R.; Bayer, K.; Kronthaler, J.; Huber, R. Advanced online monitoring of cell culture off-gas using proton transfer reaction mass spectrometry. Biotechnol. Prog. 2014, 30, 496–504. [Google Scholar] [CrossRef]
- Rogers, R.S.; Abernathy, M.; Richardson, D.D.; Rouse, J.C.; Sperry, J.B.; Swann, P.; Wypych, J.; Yu, C.; Zang, L.; Deshpande, R. A View on the Importance of “Multi-Attribute Method” for Measuring Purity of Biopharmaceuticals and Improving Overall Control Strategy. AAPS J. 2017, 20, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Migliore, N.; Mehrman, S.J.; Cunningham, J.; Lewis, M.J.; Hu, P. High-Throughput, Automated Protein A Purification Platform with Multiattribute LC–MS Analysis for Advanced Cell Culture Process Monitoring. Anal. Chem. 2016, 88, 8673–8679. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Jimenez, R.B.; Mowery, R.; Luo, H.; Cao, M.; Agarwal, N.; Ramos, I.; Wang, X.; Wang, J. A Quadrupole Dalton-based multi-attribute method for product characterization, process development, and quality control of therapeutic proteins. mAbs 2017, 9, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, P.; Sosic, Z.; Zang, L. Monitoring glycosylation profile and protein titer in cell culture samples using ZipChip CE-MS. J. Anal. Bioanal. Tech. 2017, 8, 2. [Google Scholar] [CrossRef]
- Rogstad, S.; Faustino, A.; Ruth, A.; Keire, D.; Boyne, M.; Park, J. A Retrospective Evaluation of the Use of Mass Spectrometry in FDA Biologics License Applications. J. Am. Soc. Mass Spectrom. 2017, 28, 786–794. [Google Scholar] [CrossRef]
- Bauer, I.; Poggendorf, I.; Spichiger, S.; Spichiger-Keller, U.E.; John, G. Novel single-use sensors for online measurement of glucose. BioProcess Int. 2012, 10, 56–60. [Google Scholar]
- Spichiger, S.; Spichiger-Keller, U.E. New single-use sensors for online measurement of glucose and lactate: The answer to the PAT Initiative. In Single-Use Technology in Biopharmaceutical Manufacture; Eibl, R., Eibl, D., Eds.; Wiley: Hoboken, NJ, USA, 2010; pp. 295–299. [Google Scholar]
- Wasalathanthri, D.P.; Rehmann, M.S.; Song, Y.; Gu, Y.; Mi, L.; Shao, C.; Chemmalil, L.; Lee, J.; Ghose, S.; Borys, M.C.; et al. Technology outlook for real-time quality attribute and process parameter monitoring in biopharmaceutical development—A review. Biotechnol. Bioeng. 2020, 117, 3182–3198. [Google Scholar] [CrossRef] [PubMed]
- Claßen, J.; Aupert, F.; Reardon, K.F.; Solle, D.; Scheper, T. Spectroscopic sensors for in-line bioprocess monitoring in research and pharmaceutical industrial application. Anal. Bioanal. Chem. 2017, 409, 651–666. [Google Scholar] [CrossRef]
- Kara, S.; Mueller, J.J.; Liese, A. Online analysis methods for monitoring of bioprocesses. Chim. Oggi-Chem. Today 2011, 29, 38–41. [Google Scholar]
- Landgrebe, D.; Haake, C.; Höpfner, T.; Beutel, S.; Hitzmann, B.; Scheper, T.; Rhiel, M.; Reardon, K.F. On-line infrared spectroscopy for bioprocess monitoring. Appl. Microbiol. Biotechnol. 2010, 88, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Scarff, M.; Arnold, S.A.; Harvey, L.M.; McNeil, B. Near infrared spectroscopy for bioprocess monitoring and control: Current status and future trends. Crit. Rev. Biotechnol. 2006, 26, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Rolinger, L.; Rüdt, M.; Hubbuch, J. A critical review of recent trends, and a future perspective of optical spectroscopy as PAT in biopharmaceutical downstream processing. Anal. Bioanal. Chem. 2020, 412, 2047–2064. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, N.D.; Lopes, J.A.; Almeida, C.F.; Sarraguça, M.C.; Pinheiro, H.M. Bioreactor monitoring with spectroscopy and chemometrics: A review. Anal. Bioanal. Chem. 2012, 404, 1211–1237. [Google Scholar] [CrossRef]
- Ude, C.; Schmidt-Hager, J.; Findeis, M.; John, G.T.; Scheper, T.; Beutel, S. Application of an online-biomass sensor in an optical multisensory platform prototype for growth monitoring of biotechnical relevant microorganism and cell lines in single-use shake flasks. Sensors 2014, 14, 17390–17405. [Google Scholar] [CrossRef] [Green Version]
- Edlich, A.; Magdanz, V.; Rasch, D.; Demming, S.; Aliasghar Zadeh, S.; Segura, R.; Kähler, C.; Radespiel, R.; Büttgenbach, S.; Franco-Lara, E.; et al. Microfluidic reactor for continuous cultivation of Saccharomyces cerevisiae. Biotechnol. Prog. 2010, 26, 1259–1270. [Google Scholar] [CrossRef]
- Teixeira, A.P.; Oliveira, R.; Alves, P.M.; Carrondo, M.J.T. Advances in on-line monitoring and control of mammalian cell cultures: Supporting the PAT initiative. Biotechnol. Adv. 2009, 27, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Randek, J.; Mandenius, C.-F. On-line soft sensing in upstream bioprocessing. Crit. Rev. Biotechnol. 2018, 38, 106–121. [Google Scholar] [CrossRef]
- Ghisaidoobe, A.B.T.; Chung, S.J. Intrinsic tryptophan fluorescence in the detection and analysis of proteins: A focus on Förster resonance energy transfer techniques. Int. J. Mol. Sci. 2014, 15, 22518–22538. [Google Scholar] [CrossRef] [PubMed]
- Beutel, S.; Henkel, S. In situ sensor techniques in modern bioprocess monitoring. Appl. Microbiol. Biotechnol. 2011, 91, 1493–1505. [Google Scholar] [CrossRef]
- Flores-Cosío, G.; Herrera-López, E.J.; Arellano-Plaza, M.; Gschaedler-Mathis, A.; Kirchmayr, M.; Amaya-Delgado, L. Application of dielectric spectroscopy to unravel the physiological state of microorganisms: Current state, prospects and limits. Appl. Microbiol. Biotechnol. 2020, 104, 6101–6113. [Google Scholar] [CrossRef] [PubMed]
- Cannizzaro, C.; Gügerli, R.; Marison, I.; von Stockar, U. On-line biomass monitoring of CHO perfusion culture with scanning dielectric spectroscopy. Biotechnol. Bioeng. 2003, 84, 597–610. [Google Scholar] [CrossRef]
- Yardley, J.E.; Kell, D.B.; Barrett, J.; Davey, C.L. On-Line, Real-Time Measurements of Cellular Biomass using Dielectric Spectroscopy. Biotechnol. Genet. Eng. Rev. 2000, 17, 3–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Tsang, V.L.; Moore, B.; Shen, V.; Huang, Y.M.; Kshirsagar, R.; Ryll, T. Advanced process monitoring and feedback control to enhance cell culture process production and robustness. Biotechnol. Bioeng. 2015, 112, 2495–2504. [Google Scholar] [CrossRef]
- Ulber, R.; Frerichs, J.-G.; Beutel, S. Optical sensor systems for bioprocess monitoring. Anal. Bioanal. Chem. 2003, 376, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Harms, P.; Kostov, Y.; Rao, G. Bioprocess monitoring. Curr. Opin. Biotechnol. 2002, 13, 124–127. [Google Scholar] [CrossRef]
- Hanson, M.A.; Ge, X.; Kostov, Y.; Brorson, K.A.; Moreira, A.R.; Rao, G. Comparisons of optical pH and dissolved oxygen sensors with traditional electrochemical probes during mammalian cell culture. Biotechnol. Bioeng. 2007, 97, 833–841. [Google Scholar] [CrossRef]
- Stine, J.M.; Beardslee, L.A.; Sathyam, R.M.; Bentley, W.E.; Ghodssi, R. Electrochemical Dissolved Oxygen Sensor-Integrated Platform for Wireless In Situ Bioprocess Monitoring. Sens. Actuators B Chem. 2020, 320, 128381. [Google Scholar] [CrossRef]
- Tric, M.; Lederle, M.; Neuner, L.; Dolgowjasow, I.; Wiedemann, P.; Wölfl, S.; Werner, T. Optical biosensor optimized for continuous in-line glucose monitoring in animal cell culture. Anal. Bioanal. Chem. 2017, 409, 5711–5721. [Google Scholar] [CrossRef] [PubMed]
- Vojinović, V.; Cabral, J.M.S.; Fonseca, L.P. Real-time bioprocess monitoring: Part I: In situ sensors. Sens. Actuators B Chem. 2006, 114, 1083–1091. [Google Scholar] [CrossRef]
- Xu, J.; Tang, P.; Yongky, A.; Drew, B.; Borys, M.C.; Liu, S.; Li, Z.J. Systematic development of temperature shift strategies for Chinese hamster ovary cells based on short duration cultures and kinetic modeling. mAbs 2019, 11, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KDBIO. Flow Cell Biosensor for Glucose and/or Lactate. Available online: https://www.kdbio.com/products/glucose-lactate-flow-cell-biosensor/ (accessed on 22 August 2021).
- Luttmann, R.; Bracewell, D.G.; Cornelissen, G.; Gernaey, K.V.; Glassey, J.; Hass, V.C.; Kaiser, C.; Preusse, C.; Striedner, G.; Mandenius, C.-F. Soft sensors in bioprocessing: A status report and recommendations. Biotechnol. J. 2012, 7, 1040–1048. [Google Scholar] [CrossRef]
- Carrondo, M.J.T.; Alves, P.M.; Carinhas, N.; Glassey, J.; Hesse, F.; Merten, O.-W.; Micheletti, M.; Noll, T.; Oliveira, R.; Reichl, U.; et al. How can measurement, monitoring, modeling and control advance cell culture in industrial biotechnology? Biotechnol. J. 2012, 7, 1522–1529. [Google Scholar] [CrossRef]
- Sommeregger, W.; Sissolak, B.; Kandra, K.; von Stosch, M.; Mayer, M.; Striedner, G. Quality by control: Towards model predictive control of mammalian cell culture bioprocesses. Biotechnol. J. 2017, 12, 1600546. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Kurian, V.; Ogunnaike, B.A. Bioprocess systems analysis, modeling, estimation, and control. Curr. Opin. Chem. Eng. 2021, 33, 100705. [Google Scholar] [CrossRef]
- Mandenius, C.-F.; Gustavsson, R. Mini-review: Soft sensors as means for PAT in the manufacture of bio-therapeutics. J. Chem. Technol. Biotechnol. 2015, 90, 215–227. [Google Scholar] [CrossRef]
- Sha, S.; Huang, Z.; Wang, Z.; Yoon, S. Mechanistic modeling and applications for CHO cell culture development and production. Curr. Opin. Chem. Eng. 2018, 22, 54–61. [Google Scholar] [CrossRef]
- Ohadi, K.; Legge, R.L.; Budman, H.M. Development of a soft-sensor based on multi-wavelength fluorescence spectroscopy and a dynamic metabolic model for monitoring mammalian cell cultures. Biotechnol. Bioeng. 2015, 112, 197–208. [Google Scholar] [CrossRef]
- Narayanan, H.; Behle, L.; Luna, M.F.; Sokolov, M.; Guillén-Gosálbez, G.; Morbidelli, M.; Butté, A. Hybrid-EKF: Hybrid model coupled with extended Kalman filter for real-time monitoring and control of mammalian cell culture. Biotechnol. Bioeng. 2020, 117, 2703–2714. [Google Scholar] [CrossRef]
- Tsopanoglou, A.; Jiménez del Val, I. Moving towards an era of hybrid modelling: Advantages and challenges of coupling mechanistic and data-driven models for upstream pharmaceutical bioprocesses. Curr. Opin. Chem. Eng. 2021, 32, 100691. [Google Scholar] [CrossRef]
- Simutis, R.; Lübbert, A. Hybrid Approach to State Estimation for Bioprocess Control. Bioengineering 2017, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Amribt, Z.; Dewasme, L.; Wouwer, A.V.; Bogaerts, P. Parameter Identification for State Estimation: Design of an Extended Kalman Filter for Hybridoma Cell Fed-Batch Cultures. IFAC Proc. Vol. 2014, 47, 1170–1175. [Google Scholar] [CrossRef] [Green Version]
- Hille, R.; Brandt, H.; Colditz, V.; Classen, J.; Hebing, L.; Langer, M.; Kreye, S.; Neymann, T.; Krämer, S.; Tränkle, J.; et al. Application of Model-based Online Monitoring and Robust Optimizing Control to Fed-Batch Bioprocesses. IFAC-Pap. 2020, 53, 16846–16851. [Google Scholar] [CrossRef]
- Kroll, P.; Stelzer, I.V.; Herwig, C. Soft sensor for monitoring biomass subpopulations in mammalian cell culture processes. Biotechnol. Lett. 2017, 39, 1667–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohadi, K.; Aghamohseni, H.; Legge, R.L.; Budman, H.M. Fluorescence-based soft sensor for at situ monitoring of chinese hamster ovary cell cultures. Biotechnol. Bioeng. 2014, 111, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Schwab, K.; Amann, T.; Schmid, J.; Handrick, R.; Hesse, F. Exploring the capabilities of fluorometric online monitoring on chinese hamster ovary cell cultivations producing a monoclonal antibody. Biotechnol. Prog. 2016, 32, 1592–1600. [Google Scholar] [CrossRef] [PubMed]
- Kozma, B.; Salgó, A.; Gergely, S. Comparison of multivariate data analysis techniques to improve glucose concentration prediction in mammalian cell cultivations by Raman spectroscopy. J. Pharm. Biomed. Anal. 2018, 158, 269–279. [Google Scholar] [CrossRef]
- Rafferty, C.; O’Mahony, J.; Burgoyne, B.; Rea, R.; Balss, K.M.; Latshaw II, D.C. Raman spectroscopy as a method to replace off-line pH during mammalian cell culture processes. Biotechnol. Bioeng. 2020, 117, 146–156. [Google Scholar] [CrossRef]
- Bhatia, H.; Mehdizadeh, H.; Drapeau, D.; Yoon, S. In line monitoring of amino acids in mammalian cell cultures using Raman spectroscopy and multivariate chemometrics models. Eng. Life Sci. 2018, 18, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yang, O.; Sampat, C.; Bhalode, P.; Ramachandran, R.; Ierapetritou, M. Digital Twins in Pharmaceutical and Biopharmaceutical Manufacturing: A Literature Review. Processes 2020, 8, 1088. [Google Scholar] [CrossRef]
- Brunner, V.; Siegl, M.; Geier, D.; Becker, T. Challenges in the Development of Soft Sensors for Bioprocesses: A Critical Review. Front. Bioeng. Biotechnol. 2021, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Kabugo, J.C.; Jämsä-Jounela, S.-L.; Schiemann, R.; Binder, C. Industry 4.0 based process data analytics platform: A waste-to-energy plant case study. Int. J. Electr. Power Energy Syst. 2020, 115, 105508. [Google Scholar] [CrossRef]
- Mears, L.; Stocks, S.M.; Sin, G.; Gernaey, K.V. A review of control strategies for manipulating the feed rate in fed-batch fermentation processes. J. Biotechnol. 2017, 245, 34–46. [Google Scholar] [CrossRef]
- Rathore, A.S.; Mishra, S.; Nikita, S.; Priyanka, P. Bioprocess Control: Current Progress and Future Perspectives. Life 2021, 11, 557. [Google Scholar] [CrossRef]
- Velez-Suberbie, M.L.; Betts, J.P.J.; Walker, K.L.; Robinson, C.; Zoro, B.; Keshavarz-Moore, E. High throughput automated microbial bioreactor system used for clone selection and rapid scale-down process optimization. Biotechnol. Prog. 2018, 34, 58–68. [Google Scholar] [CrossRef]
- Honda, H.; Kobayashi, T. Fuzzy control of bioprocess. J. Biosci. Bioeng. 2000, 89, 401–408. [Google Scholar] [CrossRef]
- Anderlei, T.; Schulte, A.; Laidlaw, D. Kuhner TOM for off-gas analysis in shake flasks. In Proceedings of the Recent Advances in Fermentation Technology (RAFT 13), Bonita Springs, FL, USA, 27–30 October 2019. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Type | Bioprocess Parameter | Sensor Type |
|---|---|---|
| Physical | Temperature | Thermostat, thermistor |
| Foam | Conductance | |
| Viscosity | Viscometer | |
| Pressure | Capacitance | |
| Stirring | Torque | |
| Chemical | Oxygen | Optical, electrochemical |
| pH | Electrochemical, optical | |
| Lactate | Spectroscopic, biochemical | |
| Glucose | Spectroscopic, biochemical | |
| Carbon Dioxide | Optical, electrochemical | |
| Biological | Cell count (viable cell density, total cell density, viability, cell size, aggregation) | Microscopy, spectroscopic |
| Protein | Spectroscopy | |
| Cellular morphology | Flow cytometry, spectroscopic | |
| Intermediate metabolites | Spectroscopy |
| Characteristic | Definition | Remarks |
|---|---|---|
| Selectivity | Ability to detect analyte of interest or a group of analytes. | One example of selectivity in a biosensor is the interaction of an antigen with the antibody. |
| Reproducibility | Capacity of the sensor to generate identical responses in separate experimental runs. | This is usually characterized by measuring variance, standard deviation, or coefficient of variation. This is important in bioprocess, given that manufacturing runs depend on specific sensors that are reused as in the case of stainless-steel or glass bioreactors. |
| Accuracy | Ability of the sensor to determine a mean value similar to the true value when the analyte is measured more than once. | It is generally expressed as a percentage of full-range output. If the accuracy of the sensor is high, the difference between the measured analyte value and the real analyte value is small. |
| Stability | Capacity of the sensor to produce an identical output for a constant input over a certain period of time. | It represents the degree of susceptibility that the sensor has to environmental disturbances. Over compounded time, such disturbances can generate a drift in output signals. |
| Sensitivity | Magnitude of output signal per unit change in the variable of interest. It is the relationship between the input physical signal and the output electrical signal. | The sensitivity can also be described as the Limit Of Detection (LOD) of the sensor, which is the concentration at which the mean output signal value is equal to two standard deviations. If a sensor possesses both high selectivity and high sensitivity, it is able to detect and quantify small concentrations of the analyte of interest in the presence of various substances. |
| Resolution | Smallest change in variable that is sufficient to elicit a response from the sensor. | This is key in metabolite monitoring where concentrations within the cell culture broth can be very low and, thus, differentiating slight changes of small concentrations is critical. |
| Linearity | Accuracy of the output response with respect to a straight line. | Non-linearity is an indication of deviation of the measurements from the curve of ideal measurement. |
| Response time | Speed of change in an output signal relative to a stepwise change of the input variable. | Response time should be small relative to the measured process dynamics given that long response times complicate efficient control of the process. |
| Robustness | Durability of the sensor when subjected to varying environmental conditions. | This is key in sensors that undergo sterilization and sensors that will be used on-line for long periods. |
| Technique | Sensing Attributes | References | Developer |
|---|---|---|---|
| NIR spectroscopy | Glucose, lactate, glutamine, and ammonia | [50,51,52,53] | Matrix F FT-NIR, Bruker, Billerica, MA, USA Fossanalytics, Hillerød, Denmark FossNIRSystems Inc., Silver Spring, MD, USA ABB Bomem FT-NIR spectrophotometer, Quebec City, QC, Canada Antaris II MX, Thermo Fisher Scientific, Madison, WI, USA Sartorius Stedim Biotech GmbH, Göttingen, Germany |
| MIR spectroscopy | Cell density, cell viability, lactate dehydrogenase (LDH), secreted antibody, glutamate, lactate, glucose, glutamine, ammonia. | [54,55] | Merck Millipore, Danvers, MA, USA MATRIX-MF, Bruker Optik GmbH, Ettlingen, Germany Mettler Toledo AutoChem, Inc., Columbia, SC, USA |
| UV-Vis spectroscopy | Cell density, viability, glutamine, glutamate, glucose, lactate. | [56,57,58,59,60] | J&M Analytik AG, Esslingen, Germany Thermo Fisher Scientific, Waltham, MA, USA |
| Fluorescence spectroscopy | Cell density, cell viability, recombinant protein, glucose, and ammonia concentrations. | [61,62,63,64] | J&M Analytik AG, Essingen, Germany LabX, Midland, ON, Canada Cary Eclipse CA, USA BioView, Delta Light and Optics, Denmark Horiba Jobin Yvon Fluoromax-4, Kyoto, Japan |
| Raman spectroscopy | Glycoprotein yield, Glucose, glutamine, lactate, ammonia, glutamate, cell density and viability | [65,66,67,68,69,70,71,72,73,74] | VALON Instruments Ltd., Belfast, United Kingdom Kaiser Optical Instruments, Ann Arbor, MI, USA. Perkin-Elmer, Waltham, MA, USA |
| Dielectric spectroscopy | Cell density and viability, viable cell volume | [75,76,77,78,79,80,81,82] | Aber instruments, Aberystwyth, United Kingdom Hamilton, NV, USA |
| * Optical sensors: O2 | Dissolved oxygen (DO) | [83,84,85,86] | Ocean Insight, Orlando, FL, USA Presens, Regensburg, Germany Mettler Toledo, Greifensee, Switzerland. Hamilton, NV, USA |
| * Optical sensors: pH | pH | [41,87] | Pyroscience, Aachen, Germany Presens, Regensburg, Germany Mettler Toledo, Greifensee, Switzerland Hamilton, NV, USA |
| * Optical sensors: CO2 | Dissolved CO2 | [38,88,89,90] | Presens, Regensburg, Germany Mettler Toledo, Greifensee, Switzerland Hamilton, NV, USA |
| Mass spectrometry | CO2, O2 Volatile organic compounds Aglycosylation, glycosylation, and glycation profiles | [91,92,93,94,95,96,97,98,99,100,101,102,103] | Q Exactive, Thermo Fisher Scientific, Winsford, UK Ionimed Analytik, Innsbruck, Austria Xevo G2-XS Q-TOF, Waters, Milford, CT, USA |
| Free-floating wireless sensors | Temperature, conductivity, pH, pressure, and turbidity | [49] | smartCAPS, smartINST, Lyon, France |
| Biosensors | Glucose, lactate, glutamate | [104,105] | C-CIT Sensors AG, Switzerland |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes, S.J.; Durocher, Y.; Pham, P.L.; Henry, O. Modern Sensor Tools and Techniques for Monitoring, Controlling, and Improving Cell Culture Processes. Processes 2022, 10, 189. https://doi.org/10.3390/pr10020189
Reyes SJ, Durocher Y, Pham PL, Henry O. Modern Sensor Tools and Techniques for Monitoring, Controlling, and Improving Cell Culture Processes. Processes. 2022; 10(2):189. https://doi.org/10.3390/pr10020189
Chicago/Turabian StyleReyes, Sebastian Juan, Yves Durocher, Phuong Lan Pham, and Olivier Henry. 2022. "Modern Sensor Tools and Techniques for Monitoring, Controlling, and Improving Cell Culture Processes" Processes 10, no. 2: 189. https://doi.org/10.3390/pr10020189