
Immunopathology of SARS-CoV-2 Infection: A Focus on T Regulatory and B Cell Responses in Children Compared with Adults

 , , , ,
, , , ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
- -
- Asymptomatic infection: without any clinical symptoms and signs, nor abnormal radiologic findings.
- -
- Mild: symptoms of acute upper respiratory tract infection, including fever, fatigue, myalgia, cough, sore throat, runny nose, and sneezing.
- -
- Moderate: pneumonia, associated often but not necessarily with fever, and cough (mostly dry cough, followed by productive cough); oxygen saturation is >92% without any hypoxia manifestation.
- -
- Severe: early respiratory symptoms, such as fever and cough, may be accompanied by gastrointestinal symptoms, such as diarrhea. Oxygen saturation is <92% with other hypoxia manifestations.
2.1. Inclusion and Exclusion Criteria
2.2. Cytokines Analyses
2.3. Flow Cytometry Analysis
2.4. Ethic Committee Approval
2.5. Quantification and Statistical Analysis
3. Results
3.1. Study Population
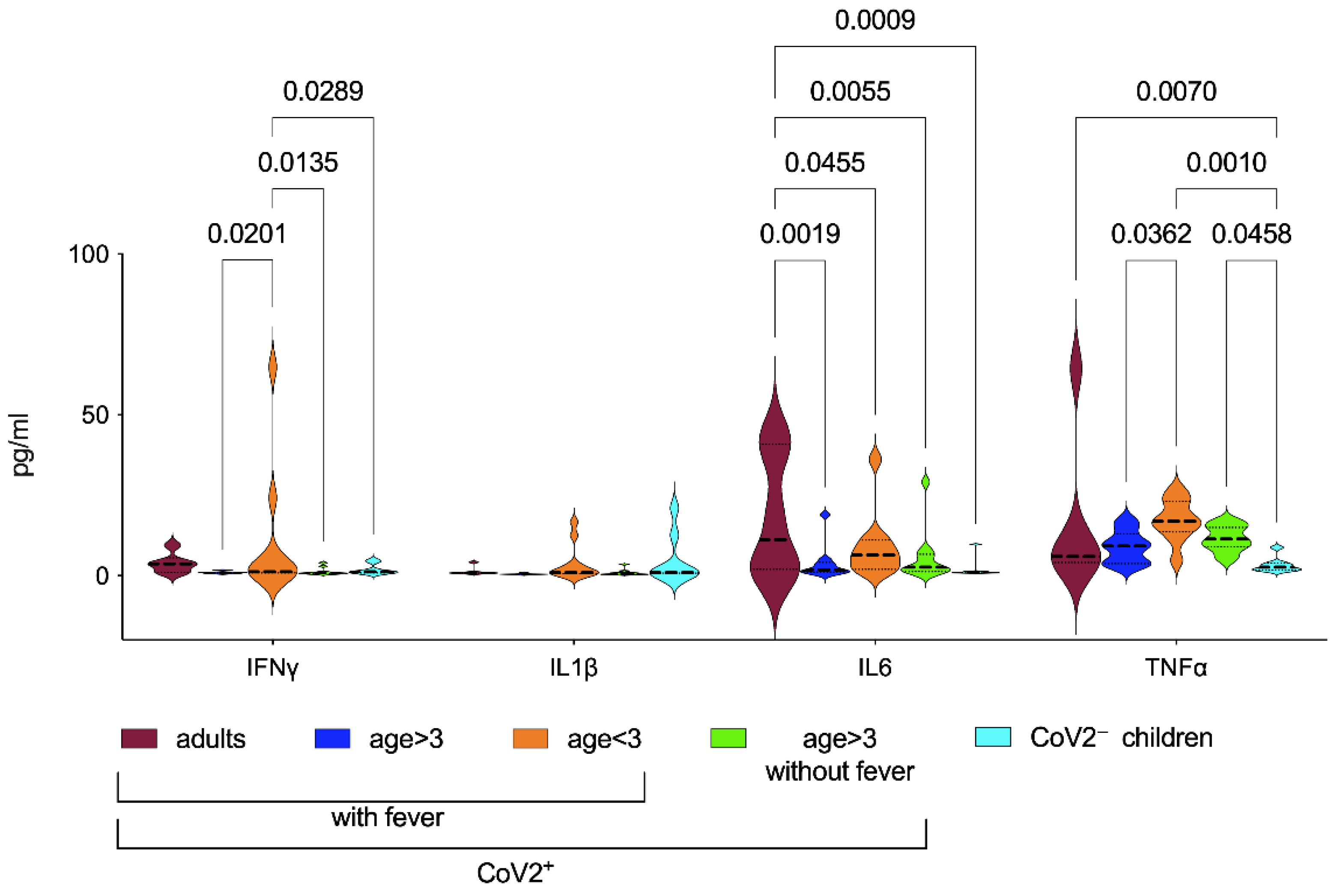
3.2. Inflammatory Biomarkers in CoV2+ Patients
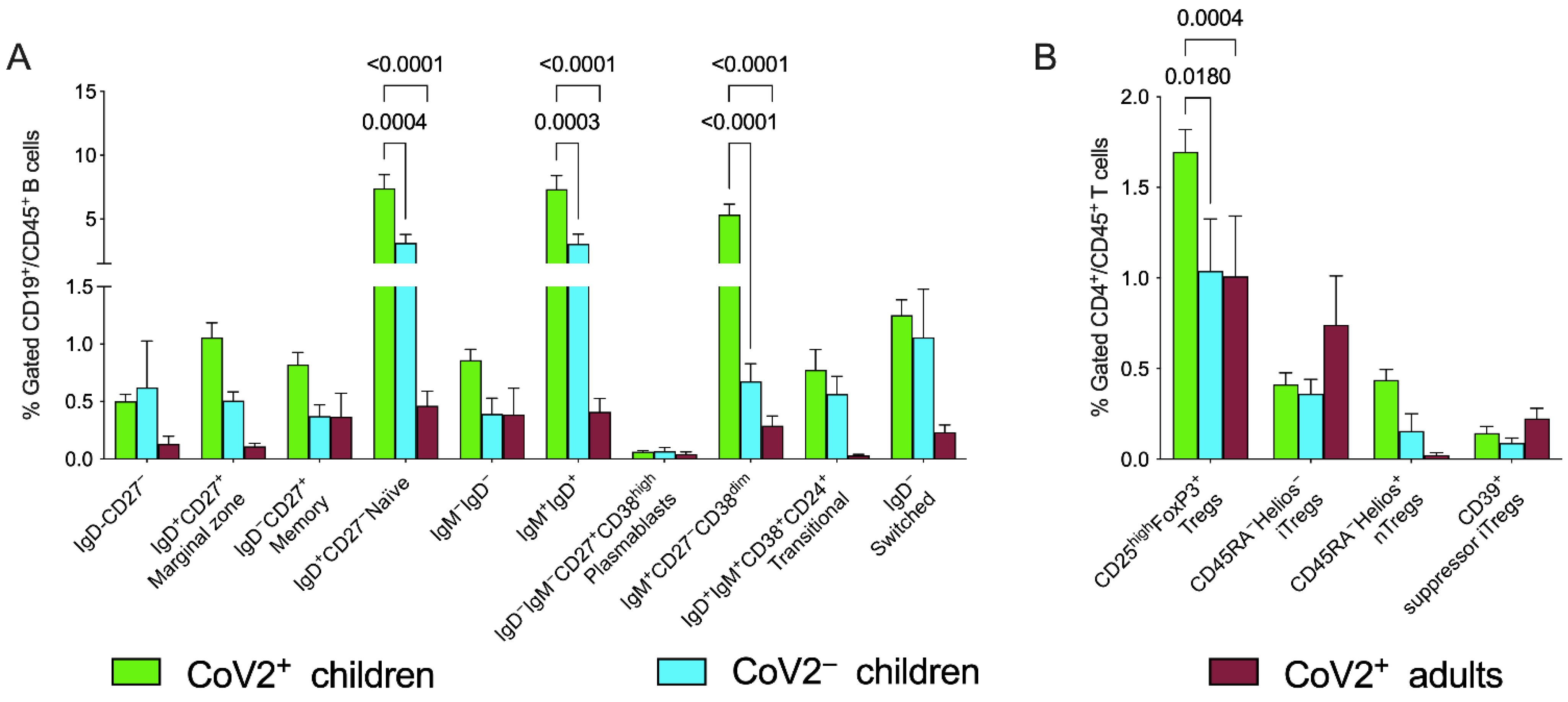
3.3. Peripheral Blood Distribution of B Cells and Treg Subsets in Patients with CoV2 and Controls at Baseline
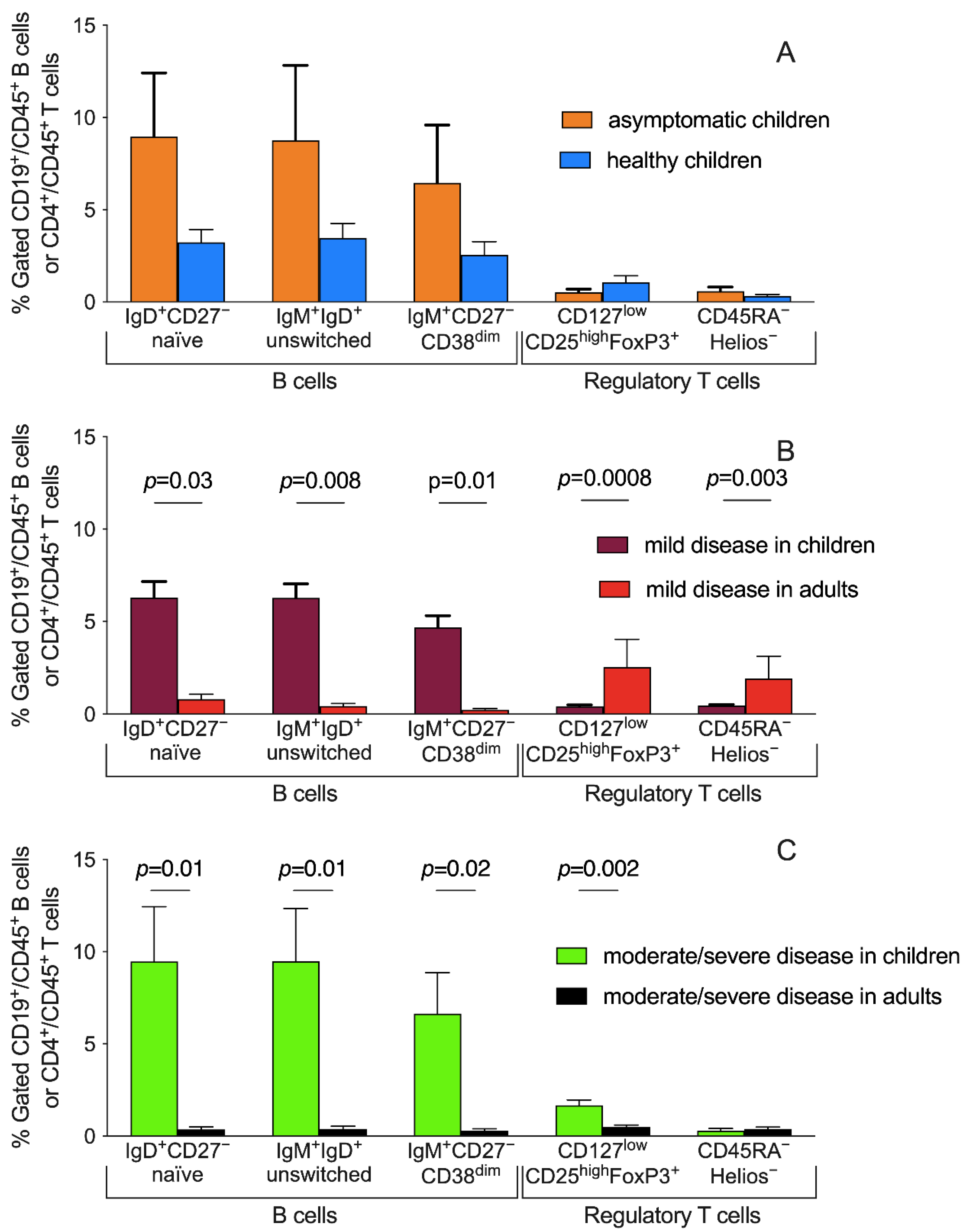
3.4. Peripheral Blood Distribution of B Cells and Treg Subsets and Disease Severity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parri, N.; Lenge, M.; Buonsenso, D. Children with COVID-19 in Pediatric Emergency Departments in Italy. N. Engl. J. Med. 2020, 383, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Götzinger, F.; Santiago-García, B.; Noguera-Julián, A.; Lanaspa, M.; Lancella, L.; Calò Carducci, F.I.; Gabrovska, N.; Velizarova, S.; Prunk, P.; Osterman, V.; et al. COVID-19 in children and adolescents in Europe: A multinational, multicentre cohort study. Lancet Child Adolesc. Health 2020, 4, 653–661. [Google Scholar] [CrossRef]
- Dong, Y.; Mo, X.; Hu, Y.; Qi, X.; Jiang, F.; Jiang, Z.; Tong, S. Epidemiology of COVID-19 Among Children in China. Pediatrics 2020, 145, e20200702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC COVID-19 Response Team; Bialek, S.; Boundy, E.; Bowen, V.; Chow, N.; Cohn, A.; Dowling, N.; Ellington, S.; Gierke, R.; Hall, A.; et al. Severe Outcomes Among Patients with Coronavirus Disease 2019 (COVID-19)—United States, February 12–March 16, 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Onder, G.; Rezza, G.; Brusaferro, S. Case-Fatality Rate and Characteristics of Patients Dying in Relation to COVID-19 in Italy. JAMA 2020, 323, 1775–1776. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72 314 Cases From the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Feldstein, L.R.; Rose, E.B.; Horwitz, S.M.; Collins, J.P.; Newhams, M.M.; Son, M.B.F.; Newburger, J.W.; Kleinman, L.C.; Heidemann, S.M.; Martin, A.A.; et al. Multisystem Inflammatory Syndrome in U.S. Children and Adolescents. N. Engl. J. Med. 2020, 383, 334–346. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Du, H.; Zhang, J.; Li, Y.Y.; Qu, J.; Zhang, W.; Wang, Y.; Bao, S.; Li, Y.; et al. CoV2 Infection in Children. N. Engl. J. Med. 2020, 382, 1663–1665. [Google Scholar] [CrossRef] [Green Version]
- Bai, K.; Liu, W.; Liu, C.; Fu, Y.; Hu, J.; Qin, Y.; Zhang, Q.; Chen, H.; Xu, F.; Li, C. Clinical Analysis of 25 COVID-19 Infections in Children. Pediatric Infect. Dis. J. 2020, 39, e100–e103. [Google Scholar] [CrossRef]
- CMMID COVID-19 Working Group; Davies, N.G.; Klepac, P.; Liu, Y.; Prem, K.; Jit, M.; Eggo, R.M. Age-dependent effects in the transmission and control of COVID-19 epidemics. Nat. Med. 2020, 26, 1205–1211. [Google Scholar] [CrossRef]
- Sinha, P.; Matthay, M.A.; Calfee, C.S. Is a “Cytokine Storm” Relevant to COVID-19? JAMA Intern. Med. 2020, 180, 1152–1154. [Google Scholar] [CrossRef] [PubMed]
- Loske, J.; Röhmel, J.; Lukassen, S.; Stricker, S.; Magalhães, V.G.; Liebig, J.; Chua, R.L.; Thürmann, L.; Messingschlager, M.; Seegebarth, A.; et al. Pre-activated antiviral innate immunity in the upper airways controls early SARS-CoV-2 infection in children. Nat. Biotechnol. 2021, 40, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.A.; Preston-Hurlburt, P.; Dai, Y.; Aschner, C.B.; Cheshenko, N.; Galen, B.; Garforth, S.J.; Herrera, N.G.; Jangra, R.K.; Morano, N.C.; et al. Immune responses to SARS-CoV-2 infection in hospitalized pediatric and adult patients. Sci. Transl. Med. 2020, 12, eabd5487. [Google Scholar] [CrossRef]
- Meckiff, B.J.; Ramírez-Suástegui, C.; Fajardo, V.; Chee, S.J.; Kusnadi, A.; Simon, H.; Eschweiler, S.; Grifoni, A.; Pelosi, E.; Weiskopf, D.; et al. Imbalance of Regulatory and Cytotoxic SARS-CoV-2-Reactive CD4+ T Cells in COVID-19. Cell 2020, 183, 1340–1353.e16. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Tian, Y.; Sette, A.; Weiskopf, D. Transcriptomic immune profiles of human flavivirus-specific T-cell responses. Immunology 2020, 160, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Consiglio, C.R.; Cotugno, N.; Sardh, F.; Pou, C.; Amodio, D.; Rodriguez, L.; Tan, Z.; Zicari, S.; Ruggiero, A.; Pascucci, G.R.; et al. The Immunology of Multisystem Inflammatory Syndrome in Children with COVID-19. Cell 2020, 183, 968–981.e7. [Google Scholar] [CrossRef]
- Cotugno, N.; Ruggiero, A.; Bonfante, F.; Petrara, M.R.; Zicari, S.; Pascucci, G.R.; Zangari, P.; De Ioris, M.A.; Santilli, V.; Manno, E.C.; et al. Virological and immunological features of SARS-CoV-2-infected children who develop neutralizing antibodies. Cell Rep. 2021, 34, 108852. [Google Scholar] [CrossRef]
- World Health Organization. Clinical Management of COVID-19—Interim Guidance. 2020. Available online: https://apps.who.int/iris/bitstream/handle/10665/332196/WHO-2019-nCoV-clinical-2020.5-eng.pdf?sequence=1&isAllowed=y (accessed on 18 January 2022).
- Kleiner, G.; Marcuzzi, A.; Zanin, V.; Monasta, L.; Zauli, G. Cytokine Levels in the Serum of Healthy Subjects. Mediat. Inflamm. 2013, 2013, 434010. [Google Scholar] [CrossRef]
- Hartel, C.; Adam, N.; Strunk, T.; Temming, P.; Muller-Steinhardt, M.; Schultz, C. Cytokine responses correlate differentially with age in infancy and early childhood. Clin. Exp. Immunol. 2005, 142, 446–453. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, L.; Xia, Y.; Wang, W.; Wang, Y.; Li, L.; Qi, Z.; Zhong, L.; Sun, J.; Tang, W.; et al. Reference values for peripheral blood lymphocyte subsets of healthy children in China. J. Allergy Clin. Immunol. 2018, 142, 970–973.e8. [Google Scholar] [CrossRef]
- Schatorjé, E.J.H.; Gemen, E.F.A.; Driessen, G.J.A.; Leuvenink, J.; van Hout, R.W.N.M.; de Vries, E. Pediatric Reference Values for the Peripheral T cell Compartment: T Lymphocyte Subsets in Children. Scand. J. Immunol. 2012, 75, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Altmann, D.M.; Boyton, R.J. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci. Immunol. 2020, 5, eabd6160. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Schmitz, K.S.; Raadsen, M.P.; Grifoni, A.; Okba, N.M.A.; Endeman, H.; van den Akker, J.P.C.; Molenkamp, R.; Koopmans, M.P.G.; van Gorp, E.C.M.; et al. Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Sci. Immunol. 2020, 5, eabd2071. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Diorio, C.; Goodwin, E.C.; McNerney, K.O.; Weirick, M.E.; Gouma, S.; Bolton, M.J.; Arevalo, C.P.; Chase, J.; Hicks, P.; et al. SARS-CoV-2 antibody responses in children with MIS-C and mild and severe COVID-19. Infect. Dis. (except HIV/AIDS) 2020. [Google Scholar] [CrossRef]
- Anderson, E.M.; Diorio, C.; Goodwin, E.C.; McNerney, K.O.; Weirick, M.E.; Gouma, S.; Bolton, M.J.; Arevalo, C.P.; Chase, J.; Hicks, P.; et al. Severe Acute Respiratory Syndrome-Coronavirus-2 (SARS-CoV-2) Antibody Responses in Children With Multisystem Inflammatory Syndrome in Children (MIS-C) and Mild and Severe Coronavirus Disease 2019 (COVID-19). J. Pediatric Infect. Dis. Soc. 2021, 10, 669–673. [Google Scholar] [CrossRef]
- Mold, J.E.; Venkatasubrahmanyam, S.; Burt, T.D.; Michaelsson, J.; Rivera, J.M.; Galkina, S.A.; Weinberg, K.; Stoddart, C.A.; McCune, J.M. Fetal and Adult Hematopoietic Stem Cells Give Rise to Distinct T Cell Lineages in Humans. Science 2010, 330, 1695–1699. [Google Scholar] [CrossRef] [Green Version]
- Diaz, F.; Bustos, B.R.; Yagnam, F.; Karsies, T.J.; Vásquez-Hoyos, P.; Jaramillo-Bustamante, J.-C.; Gonzalez-Dambrauskas, S.; Drago, M.; Cruces, P. Comparison of Interleukin-6 Plasma Concentration in Multisystem Inflammatory Syndrome in Children Associated With SARS-CoV-2 and Pediatric Sepsis. Front. Pediatr. 2021, 9, 756083. [Google Scholar] [CrossRef]
- Saule, P.; Trauet, J.; Dutriez, V.; Lekeux, V.; Dessaint, J.-P.; Labalette, M. Accumulation of memory T cells from childhood to old age: Central and effector memory cells in CD4+ versus effector memory and terminally differentiated memory cells in CD8+ compartment. Mech. Ageing Dev. 2006, 127, 274–281. [Google Scholar] [CrossRef]
- Rudolph, M.E.; McArthur, M.A.; Barnes, R.S.; Magder, L.S.; Chen, W.H.; Sztein, M.B. Differences Between Pediatric and Adult T Cell Responses to In Vitro Staphylococcal Enterotoxin B Stimulation. Front. Immunol. 2018, 9, 498. [Google Scholar] [CrossRef] [Green Version]
- Cohen, C.A.; Li, A.P.; Hachim, A.; Hui, D.S.; Kwan, M.Y.; Tsang, O.T.; Chiu, S.S.; Chan, W.H.; Yau, Y.S.; Kavian, N.; et al. SARS-CoV-2 specific T cell responses are lower in children and increase with age and time after infection. Nat. Commun. 2021, 12, 4678. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Yang, L.; Li, X.; Sun, M.; Zhang, A.; Qi, S.; Chen, Z.; Zhang, L.; Li, J.; Xiong, H. Early immune responses and prognostic factors in children with COVID-19: A single-center retrospective analysis. BMC Pediatr. 2021, 21, 181. [Google Scholar] [CrossRef] [PubMed]
- Grimsholm, O.; Piano Mortari, E.; Davydov, A.N.; Shugay, M.; Obraztsova, A.S.; Bocci, C.; Marasco, E.; Marcellini, V.; Aranburu, A.; Farroni, C.; et al. The Interplay between CD27dull and CD27bright B Cells Ensures the Flexibility, Stability, and Resilience of Human B Cell Memory. Cell Rep. 2020, 30, 2963–2977.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carsetti, R.; Zaffina, S.; Piano Mortari, E.; Terreri, S.; Corrente, F.; Capponi, C.; Palomba, P.; Mirabella, M.; Cascioli, S.; Palange, P.; et al. Different Innate and Adaptive Immune Responses to SARS-CoV-2 Infection of Asymptomatic, Mild, and Severe Cases. Front. Immunol. 2020, 11, 610300. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Why is COVID-19 less severe in children? A review of the proposed mechanisms underlying the age-related difference in severity of SARS-CoV-2 infections. Arch. Dis. Child. 2021, 106, 429–439. [Google Scholar] [CrossRef]
- Brodin, P. Immune determinants of COVID-19 disease presentation and severity. Nat. Med. 2021, 27, 28–33. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Bunyavanich, S.; Do, A.; Vicencio, A. Nasal Gene Expression of Angiotensin-Converting Enzyme 2 in Children and Adults. JAMA 2020, 323, 2427–2429. [Google Scholar] [CrossRef]
- Wang, A.; Chiou, J.; Poirion, O.B.; Buchanan, J.; Valdez, M.J.; Verheyden, J.M.; Hou, X.; Kudtarkar, P.; Narendra, S.; Newsome, J.M.; et al. Single-cell multiomic profiling of human lungs reveals cell-type-specific and age-dynamic control of SARS-CoV2 host genes. eLife 2020, 9, e62522. [Google Scholar] [CrossRef]
- Saheb Sharif-Askari, N.; Saheb Sharif-Askari, F.; Alabed, M.; Temsah, M.-H.; Al Heialy, S.; Hamid, Q.; Halwani, R. Airways Expression of SARS-CoV-2 Receptor, ACE2, and TMPRSS2 Is Lower in Children Than Adults and Increases with Smoking and COPD. Mol. Ther. Methods Clin. Dev. 2020, 18, 1–6. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Neeland, M.R.; Bannister, S.; Clifford, V.; Nguyen, J.; Dohle, K.; Overmars, I.; Toh, Z.Q.; Anderson, J.; Donato, C.M.; Sarkar, S.; et al. Children and Adults in a Household Cohort Study Have Robust Longitudinal Immune Responses Following SARS-CoV-2 Infection or Exposure. Front. Immunol. 2021, 12, 741639. [Google Scholar] [CrossRef]
- Neeland, M.R.; Bannister, S.; Clifford, V.; Dohle, K.; Mulholland, K.; Sutton, P.; Curtis, N.; Steer, A.C.; Burgner, D.P.; Crawford, N.W.; et al. Innate cell profiles during the acute and convalescent phase of SARS-CoV-2 infection in children. Nat. Commun. 2021, 12, 1084. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Worlock, K.B.; Huang, N.; Lindeboom, R.G.H.; Butler, C.R.; Kumasaka, N.; Conde, C.D.; Mamanova, L.; Bolt, L.; Richardson, L.; et al. Local and systemic responses to SARS-CoV-2 infection in children and adults. Nature 2021, 602, 321–327. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| CoV2− Children (n = 7) | CoV2+ Children (n = 40) | CoV2+ Adults (n = 16) | ||||
|---|---|---|---|---|---|---|
| 7 | Asympt. 4 (10%) | Mild 24 (60) | Mod./Severe 12 (30) | Mild 3 (18.75) | Mod./Severe 13 (81.25) | |
| Age, mean ± SD, (years) | 7.3 ± 5.9 | 8.3 ± 6.9 | 8.3 ± 6.9 | 8.5 ± 4.9 | 45.4 ± 16.9 | 64.7 ± 16.6 |
| Female n., (%) | 4 (57.1) | 2 (50) | 10 (41.7) | 2 (16.7) | 1 (33.3) | 2 (15.4) |
| Fever n., (%) | N/A | 0 | 15 (62.5) | 12 (100) | 3 (100) | 11 (84.6) |
| Febrile days, mean ± SD, | 0 | 0 | 1.8 ± 2.2 | 2.45 ± 2.66 | 2 (66.7) | 8.5 ± 2.1 |
| Cough, n., (%) | 0 | 0 | 10 (41.7) | 8 (66.7) | 3 (100) | 12 (92.3) |
| Upper airway symptom, n. (%) | 0 | 0 | 10 (41.7) | 3 (25) | 2 (66.7) | 0 |
| Anosmia, n. (%) | 0 | 0 | 1 (4.2) | 1 (8.3) | 0 | 0 |
| Dyspnea, n., (%) | 0 | 0 | 0 | 1 (8.3) | 0 | 5 (38.5) |
| Headache, n., (%) | 0 | 0 | 6 (25) | 1 (8.3) | 0 | 1 (7.7) |
| Diarrhea, n., (%) | 0 | 0 | 2 (8.3) | 3 (25) | 0 | 0 |
| Lung interstitiopathy, RX, n. (%) | 0 | 0 | 1 (4.2) | 6 (50) | 0 | 12 (92.3) |
| Lung interstitiopathy, Echo, n. (%) | 0 | 0 | 6 (25) | 10 (83.3) | Nd | Nd |
| Hospitalization needed, n., (%) | 0 | 0 | 7 (29.2) | 12 (100) | 0 | 8 (61.5) |
| O2 therapy, n., (%) | 0 | 0 | 0 | 3 (25) | 0 | 6 (46.2) |
| HFNC, n., (%) | 0 | 0 | 0 | 1 (8.3) | 0 | 5 (38.5) |
| Corticosteroids treat., n., (%) | 0 | 0 | 0 | 5 (41.7) | 0 | 4 (30.8) |
| IGIV, n., (%) | 0 | 0 | 0 | 2 (16.7) | 0 | 0 |
| Antibiotic(s) treat., n., (%) | 0 | 0 | 0 | 3 (25) | 0 | 3 (23.1) |
| RBC × 1012/L | Nd | 4.6 ± 0.6 | 4.9 ± 0.4 | 4.7 ± 0.6 | 3.9 ± 0.3 | 4.2 ± 1.4 |
| WBC cells × 109/L | Nd | 10 ± 4.6 | 8.7 ± 3.5 | 10.6 ± 3.4 | 8.3 ± 1.1 | 9.1 ± 2.4 |
| Lymphocytes × 109/L | Nd | 4.2 ± 2.3 | 3.3 ± 1.5 | 4.2 ± 2.9 | 1.5 ± 0.3 | 1.8 ± 1.3 |
| Neutrophils × 109/L | Nd | 4.7 ± 2.2 | 4.4 ± 2.8 | 5.5 ± 2.6 | 6.1 ± 0.9 | 6.7 ± 1.5 |
| Monocytes × 109/L | Nd | 2.8 ± 4.3 | 0.6 ± 2.9 | 0.8 ± 2.8 | 0.8 ± 0.1 | 0.9 ± 0.7 |
| CRP, g/L | Nd | 39 ± 48.1 | 73.9 ± 160.4 | 206.3 ± 240.6 | 47.9 ± 30.1 | 69.7 ± 38.3 |
| CoV2+ | CoV2− Children | p-Values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| With Fever | No Fever | b vs. c | b vs. E | b vs. d | c vs. e | c vs. d | d vs. e | ||||
| Adults | Age > 3 | Age < 3 | Age > 3 | ||||||||
| a | b | c | d | e | |||||||
| IL1β | 1.4 ± 1.4 | 0.5 ± 0.2 | 3.9 ± 5.8 | 0.9 ± 0.9 | 4.9 ± 7.7 | 0.38 | 0.29 | 0.91 | 0.82 | 0.43 | 0.33 |
| IL6 | 18.2 ± 19.3 | 3.8 ± 5.5 | 9.0 ± 10.2 | 5.6 ± 8.2 | 2.1 ± 3.1 | 0.19 | 0.68 | 0.64 | 0.1 | 0.38 | 0.39 |
| TNFα | 16.3 ± 23.9 | 8.9 ± 5.0 | 17.2 ± 6.4 | 11.5 ± 4.0 | 3.3 ± 2.4 | 0.04 | 0.18 | 0.5 | 0.001 | 0.14 | 0.05 |
| IFNγ | 3.8 ± 3.1 | 1.0 ± 0.4 | 11.4 ± 21.4 | 1.3 ± 1.2 | 1.9 ± 1.6 | 0.02 | 0.83 | 0.94 | 0.03 | 0.01 | 0.87 |
| Children | Adults | p-Values | |||
|---|---|---|---|---|---|
| CoV2+ | CoV2− | CoV2+ | a | b | |
| IgD+CD27− naïve B cells | 7.4 ± 6.4 | 3.1 ± 1.8 | 0.5 ± 0.5 | 0.0004 | <0.0001 |
| IgD+IgM+ B cells | 7.3 ± 6.4 | 3.1 ± 2.0 | 0.4 ± 0.5 | 0.0003 | <0.0001 |
| IgM+CD27−CD38dim B cells | 5.3 ± 4.9 | 0.7 ± 0.4 | 0.3 ± 0.3 | <0.0001 | <0.0001 |
| CD25highFOXP3+ Treg | 1.7 ± 0.8 | 1 ± 0.8 | 1 ± 1.3 | 0.018 | 0.0004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Sante, G.; Buonsenso, D.; De Rose, C.; Tredicine, M.; Palucci, I.; De Maio, F.; Camponeschi, C.; Bonadia, N.; Biasucci, D.; Pata, D.; et al. Immunopathology of SARS-CoV-2 Infection: A Focus on T Regulatory and B Cell Responses in Children Compared with Adults. Children 2022, 9, 681. https://doi.org/10.3390/children9050681
Di Sante G, Buonsenso D, De Rose C, Tredicine M, Palucci I, De Maio F, Camponeschi C, Bonadia N, Biasucci D, Pata D, et al. Immunopathology of SARS-CoV-2 Infection: A Focus on T Regulatory and B Cell Responses in Children Compared with Adults. Children. 2022; 9(5):681. https://doi.org/10.3390/children9050681
Chicago/Turabian StyleDi Sante, Gabriele, Danilo Buonsenso, Cristina De Rose, Maria Tredicine, Ivana Palucci, Flavio De Maio, Chiara Camponeschi, Nicola Bonadia, Daniele Biasucci, Davide Pata, and et al. 2022. "Immunopathology of SARS-CoV-2 Infection: A Focus on T Regulatory and B Cell Responses in Children Compared with Adults" Children 9, no. 5: 681. https://doi.org/10.3390/children9050681