

Characterization and Advancement of an Evaluation Method for the Treatment of Spontaneous Osteoarthritis in STR/ort Mice: GRGDS Peptides as a Potential Treatment for Osteoarthritis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animal Studies
2.2. Safranin O and Immunofluorescence Staining
2.3. OARSI Score
2.4. Micro-CT Bone Imaging
2.5. Statistical Analysis
3. Results
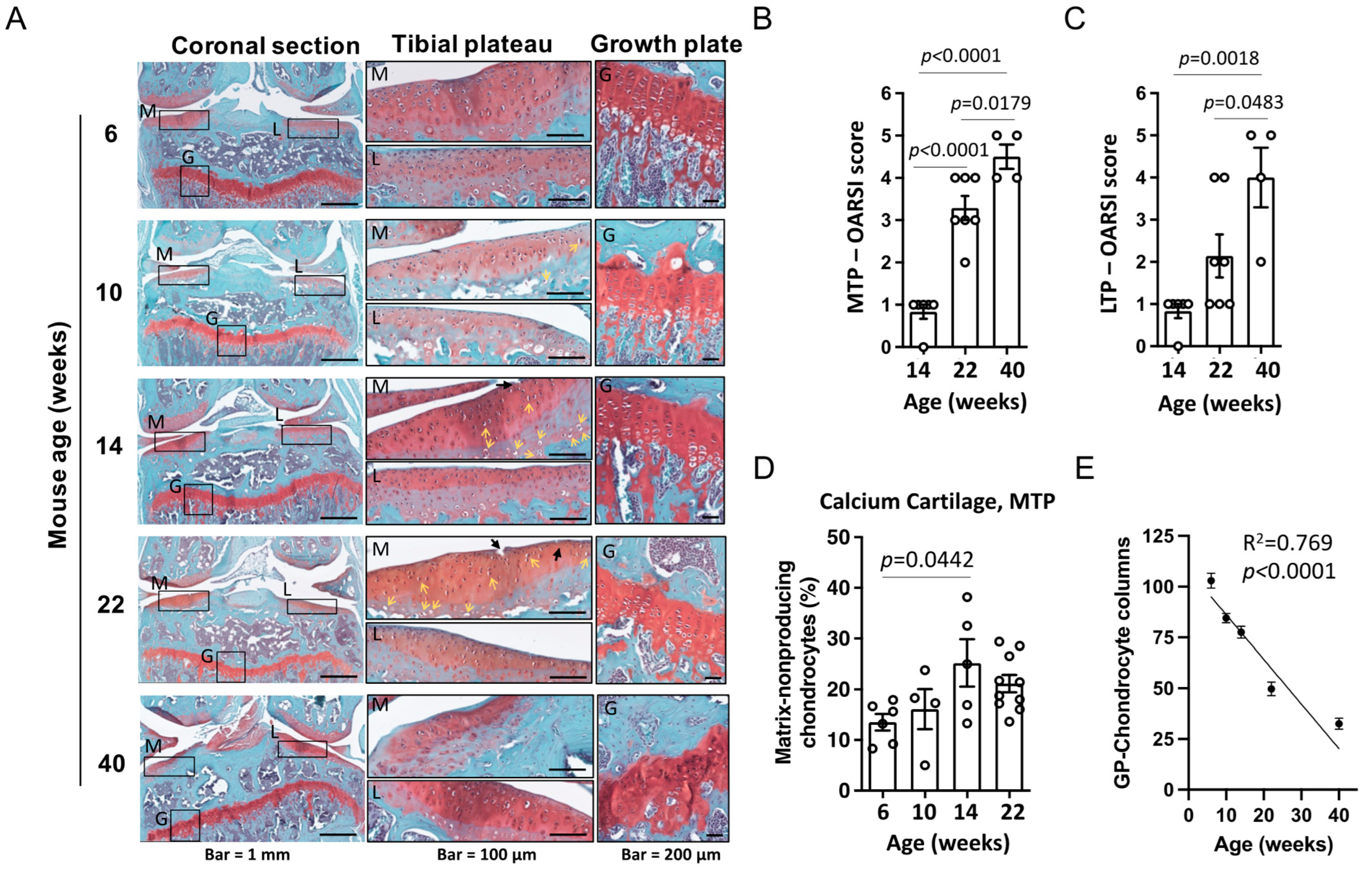
3.1. Histological Assessment of OA Severity Associated Cartilage Damage with Age
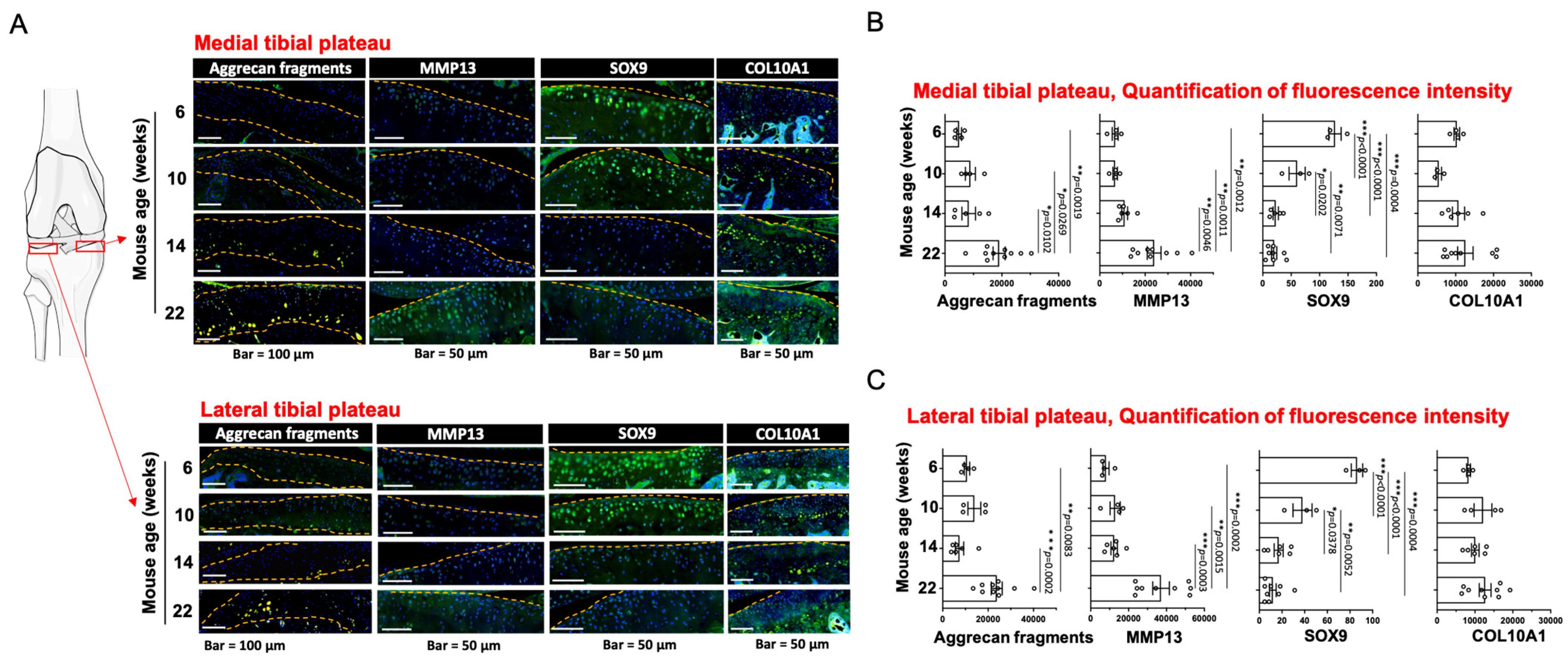
3.2. Assessment of Articular Chondrocyte Hypertrophy Associated Markers with Age
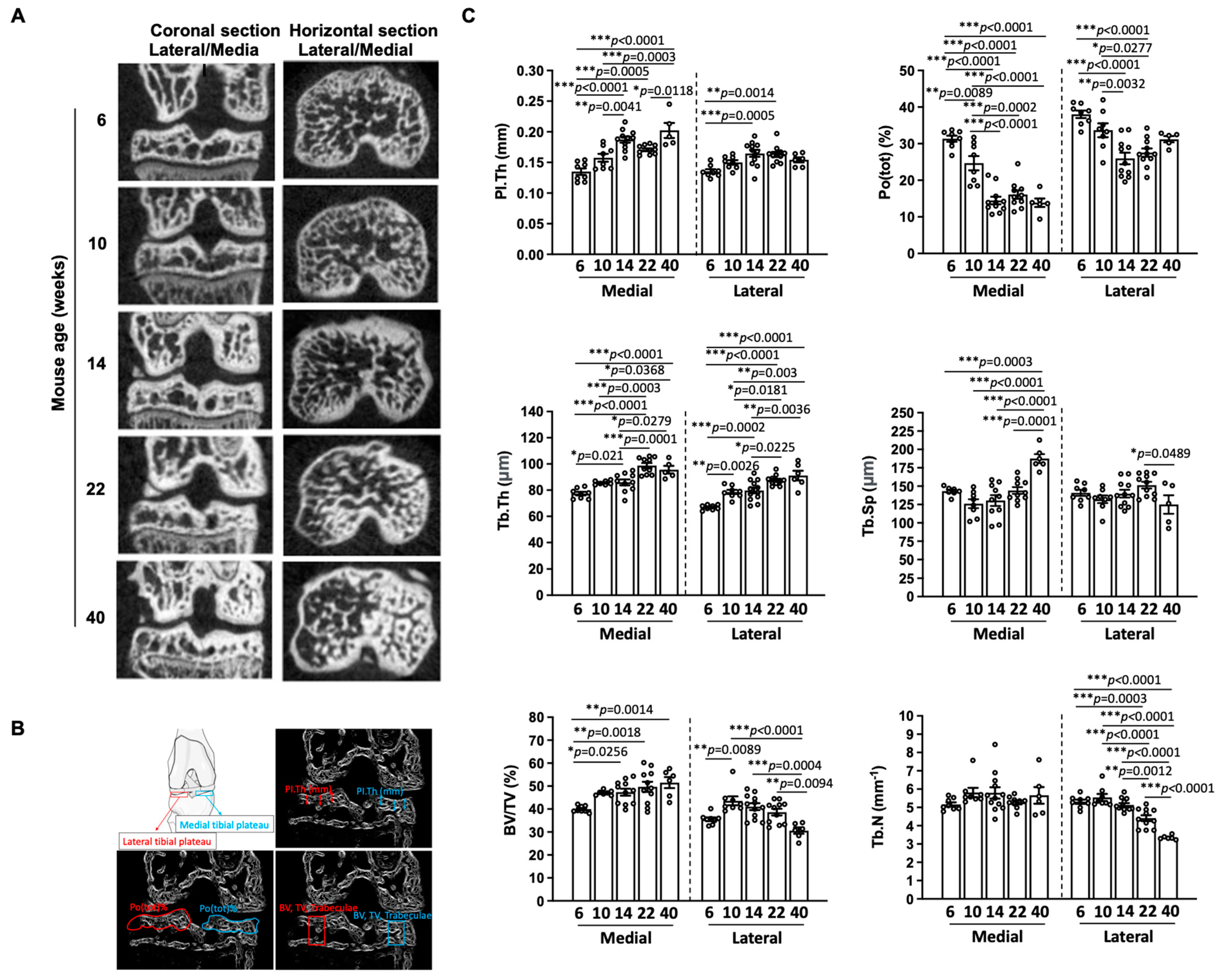
3.3. Subchondral Bone Plate Thickness and Epiphyseal Trabecular Parameters
3.4. GRGDS Treatment Prevents Cartilage Degeneration and Subchondral Bone Mineralization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dieppe, P.A.; Lohmander, L.S. Pathogenesis and management of pain in osteoarthritis. Lancet 2005, 365, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Greene, M.A.; Loeser, R.F. Aging-related inflammation in osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1966–1971. [Google Scholar] [CrossRef] [Green Version]
- Sandell, L.J.; Aigner, T. Articular cartilage and changes in arthritis. An introduction: Cell biology of osteoarthritis. Arthritis Res. 2001, 3, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Mort, J.S.; Billington, C.J. Articular cartilage and changes in arthritis: Matrix degradation. Arthritis Res. 2001, 3, 337–341. [Google Scholar] [CrossRef]
- Li, H.; Wang, D.; Yuan, Y.; Min, J. New insights on the MMP-13 regulatory network in the pathogenesis of early osteoarthritis. Arthritis Res. Ther. 2017, 19, 248. [Google Scholar] [CrossRef] [Green Version]
- Stone, A.V.; Loeser, R.F.; Vanderman, K.S.; Long, D.L.; Clark, S.C.; Ferguson, C.M. Pro-inflammatory stimulation of meniscus cells increases production of matrix metalloproteinases and additional catabolic factors involved in osteoarthritis pathogenesis. Osteoarthr. Cartil. 2014, 22, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Gratal, P.; Mediero, A.; Sanchez-Pernaute, O.; Prieto-Potin, I.; Lamuedra, A.; Herrero-Beaumont, G.; Largo, R. Chondrocyte enlargement is a marker of osteoarthritis severity. Osteoarthr. Cartil. 2019, 27, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ji, Q.; Wang, X.; Kang, L.; Fu, Y.; Yin, Y.; Li, Z.; Liu, Y.; Xu, X.; Wang, Y. SOX9 is a regulator of ADAMTSs-induced cartilage degeneration at the early stage of human osteoarthritis. Osteoarthr. Cartil. 2015, 23, 2259–2268. [Google Scholar] [CrossRef] [Green Version]
- Kuyinu, E.L.; Narayanan, G.; Nair, L.S.; Laurencin, C.T. Animal models of osteoarthritis: Classification, update, and measurement of outcomes. J. Orthop. Surg. Res. 2016, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Little, C.B.; Zaki, S.; Blaker, C.L.; Clarke, E.C. Animal models of osteoarthritis: The benefits and costs of reducing variability. Bone Joint Res. 2022, 11, 514–517. [Google Scholar] [CrossRef]
- Hu, W.; Lin, J.; Wei, J.; Yang, Y.; Fu, K.; Zhu, T.; Zhu, H.; Zheng, X. Modelling osteoarthritis in mice via surgical destabilization of the medial meniscus with or without a stereomicroscope. Bone Joint Res. 2022, 11, 518–527. [Google Scholar] [CrossRef]
- Strong, L.C. Genetic Nature of the Constitutional States of Cancer Susceptibility and Resistance in Mice and Men. Yale J. Biol. Med. 1944, 17, 289–299. [Google Scholar]
- Sokoloff, L.; Crittenden, L.B.; Yamamoto, R.S.; Jay, G.E., Jr. The genetics of degenerative joint disease in mice. Arthritis Rheum. 1962, 5, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.M.; Chambers, M.G.; Flannelly, J.; Gaffen, J.D.; Dudhia, J.; Bayliss, M.T. The STR/ort mouse and its use as a model of osteoarthritis. Osteoarthr. Cartil. 2001, 9, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Little, C.B.; Hunter, D.J. Post-traumatic osteoarthritis: From mouse models to clinical trials. Nat. Rev. Rheumatol. 2013, 9, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Staines, K.A.; Poulet, B.; Wentworth, D.N.; Pitsillides, A.A. The STR/ort mouse model of spontaneous osteoarthritis—An update. Osteoarthr. Cartil. 2017, 25, 802–808. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, K.; Suzuki, S.; Kanri, Y.; Matsubara, R.; Fujii, K.; Wake, M.; Suzuki, R.; Hamada, Y. Spontaneously developed osteoarthritis in the temporomandibular joint in STR/ort mice. Biomed. Rep. 2015, 3, 453–456. [Google Scholar] [CrossRef] [Green Version]
- Staines, K.A.; Madi, K.; Mirczuk, S.M.; Parker, S.; Burleigh, A.; Poulet, B.; Hopkinson, M.; Bodey, A.J.; Fowkes, R.C.; Farquharson, C.; et al. Endochondral Growth Defect and Deployment of Transient Chondrocyte Behaviors Underlie Osteoarthritis Onset in a Natural Murine Model. Arthritis Rheumatol. 2016, 68, 880–891. [Google Scholar] [CrossRef] [Green Version]
- Sumsuzzman, D.M.; Khan, Z.A.; Choi, J.; Hong, Y. Assessment of functional roles and therapeutic potential of integrin receptors in osteoarthritis: A systematic review and meta-analysis of preclinical studies. Ageing Res. Rev. 2022, 81, 101729. [Google Scholar] [CrossRef] [PubMed]
- Gigout, A.; Jolicoeur, M.; Nelea, M.; Raynal, N.; Farndale, R.; Buschmann, M.D. Chondrocyte aggregation in suspension culture is GFOGER-GPP- and beta1 integrin-dependent. J. Biol. Chem. 2008, 283, 31522–31530. [Google Scholar] [CrossRef] [Green Version]
- Pesesse, L.; Sanchez, C.; Delcour, J.P.; Bellahcene, A.; Baudouin, C.; Msika, P.; Henrotin, Y. Consequences of chondrocyte hypertrophy on osteoarthritic cartilage: Potential effect on angiogenesis. Osteoarthr. Cartil. 2013, 21, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments--the ARRIVE guidelines. J. Cereb. Blood Flow Metab. 2011, 31, 991–993. [Google Scholar] [CrossRef] [Green Version]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The OARSI histopathology initiative—Recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18 (Suppl. S3), S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Javaheri, B.; Razi, H.; Piles, M.; de Souza, R.; Chang, Y.M.; Maric-Mur, I.; Hopkinson, M.; Lee, P.D.; Pitsillides, A.A. Sexually dimorphic tibia shape is linked to natural osteoarthritis in STR/Ort mice. Osteoarthr. Cartil. 2018, 26, 807–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiusaroli, R.; Visentini, M.; Galimberti, C.; Casseler, C.; Mennuni, L.; Covaceuszach, S.; Lanza, M.; Ugolini, G.; Caselli, G.; Rovati, L.C.; et al. Targeting of ADAMTS5’s ancillary domain with the recombinant mAb CRB0017 ameliorates disease progression in a spontaneous murine model of osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1807–1810. [Google Scholar] [CrossRef] [Green Version]
- Simon, W.H. Scale effects in animal joints. I. Articular cartilage thickness and compressive stress. Arthritis Rheum. 1970, 13, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.F.; Martinez, A.M.; Pedoia, V.; Majumdar, S.; Vail, T.P.; Bini, S.A. Variation in the Thickness of Knee Cartilage. The Use of a Novel Machine Learning Algorithm for Cartilage Segmentation of Magnetic Resonance Images. J. Arthroplast. 2019, 34, 2210–2215. [Google Scholar] [CrossRef]
- Sward, P.; Wang, Y.; Hansson, M.; Lohmander, L.S.; Grodzinsky, A.J.; Struglics, A. Coculture of bovine cartilage with synovium and fibrous joint capsule increases aggrecanase and matrix metalloproteinase activity. Arthritis Res. Ther. 2017, 19, 157. [Google Scholar] [CrossRef] [Green Version]
- Roughley, P.J. Articular cartilage and changes in arthritis: Noncollagenous proteins and proteoglycans in the extracellular matrix of cartilage. Arthritis Res. 2001, 3, 342–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, S.; Englund, M.; Struglics, A.; Lohmander, L.S. Association between synovial fluid levels of aggrecan ARGS fragments and radiographic progression in knee osteoarthritis. Arthritis Res. Ther. 2010, 12, R230. [Google Scholar] [CrossRef] [Green Version]
- Struglics, A.; Hansson, M.; Lohmander, L.S. Human aggrecanase generated synovial fluid fragment levels are elevated directly after knee injuries due to proteolysis both in the inter globular and chondroitin sulfate domains. Osteoarthr. Cartil. 2011, 19, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Germaschewski, F.M.; Matheny, C.J.; Larkin, J.; Liu, F.; Thomas, L.R.; Saunders, J.S.; Sully, K.; Whittall, C.; Boyle, Y.; Peters, G.; et al. Quantitation OF ARGS aggrecan fragments in synovial fluid, serum and urine from osteoarthritis patients. Osteoarthr. Cartil. 2014, 22, 690–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiani, C.; Chen, L.; Wu, Y.J.; Yee, A.J.; Yang, B.B. Structure and function of aggrecan. Cell Res. 2002, 12, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inada, M.; Wang, Y.; Byrne, M.H.; Rahman, M.U.; Miyaura, C.; Lopez-Otin, C.; Krane, S.M. Critical roles for collagenase-3 (Mmp13) in development of growth plate cartilage and in endochondral ossification. Proc. Natl. Acad. Sci. USA 2004, 101, 17192–17197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, S.P.; Liang, S.; Akdemir, K.C.; de Crombrugghe, B. The postnatal role of Sox9 in cartilage. J. Bone Miner. Res. 2012, 27, 2511–2525. [Google Scholar] [CrossRef]
- Haseeb, A.; Kc, R.; Angelozzi, M.; de Charleroy, C.; Rux, D.; Tower, R.J.; Yao, L.; Pellegrino da Silva, R.; Pacifici, M.; Qin, L.; et al. SOX9 keeps growth plates and articular cartilage healthy by inhibiting chondrocyte dedifferentiation/osteoblastic redifferentiation. Proc. Natl. Acad. Sci. USA 2021, 118, e2019152118. [Google Scholar] [CrossRef]
- Wang, Z.; Boyko, T.; Tran, M.C.; LaRussa, M.; Bhatia, N.; Rashidi, V.; Longaker, M.T.; Yang, G.P. DEL1 protects against chondrocyte apoptosis through integrin binding. J. Surg. Res. 2018, 231, 1–9. [Google Scholar] [CrossRef]
- Zhang, J.; Mujeeb, A.; Du, Y.; Lin, J.; Ge, Z. Probing cell-matrix interactions in RGD-decorated macroporous poly (ethylene glycol) hydrogels for 3D chondrocyte culture. Biomed. Mater. 2015, 10, 035016. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Park, K.; Kim, J.J.; Lee, J.H.; Han, K.O.; Han, D.K. Effect of RGD-immobilized dual-pore poly(L-lactic acid) scaffolds on chondrocyte proliferation and extracellular matrix production. Artif. Organs 2008, 32, 981–989. [Google Scholar] [CrossRef]
- Tan, H.; Huang, D.; Lao, L.; Gao, C. RGD modified PLGA/gelatin microspheres as microcarriers for chondrocyte delivery. J. Biomed. Mater. Res. B Appl. Biomater. 2009, 91, 228–238. [Google Scholar] [CrossRef]
- Jo, S.; Lee, J.S.; Nam, B.; Lee, Y.L.; Kim, H.; Lee, E.Y.; Park, Y.S.; Kim, T.H. SOX9(+) enthesis cells are associated with spinal ankylosis in ankylosing spondylitis. Osteoarthr. Cartil. 2022, 30, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Yang, Z.M.; Li, Y.C.; Wang, H.X.; Lo, J.H.T.; Zhang, X.T.; Li, G. Linc-ROR promotes mesenchymal stem cells chondrogenesis and cartilage formation via regulating SOX9 expression. Osteoarthr. Cartil. 2021, 29, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, J.; Xu, F.; Ji, E.; Wang, C.; Pan, Z. Long noncoding RNA H19 alleviates inflammation in osteoarthritis through interactions between TP53, IL-38, and IL-36 receptor. Bone Joint Res. 2022, 11, 594–607. [Google Scholar] [CrossRef]
- Caron, M.M.J.; Eveque, M.; Cillero-Pastor, B.; Heeren, R.M.A.; Housmans, B.; Derks, K.; Cremers, A.; Peffers, M.J.; van Rhijn, L.W.; van den Akker, G.; et al. Sox9 Determines Translational Capacity During Early Chondrogenic Differentiation of ATDC5 Cells by Regulating Expression of Ribosome Biogenesis Factors and Ribosomal Proteins. Front. Cell Dev. Biol. 2021, 9, 686096. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Madry, H.; Venkatesan, J.K.; Schmitt, G.; Speicher-Mentges, S.; Zurakowski, D.; Menger, M.D.; Laschke, M.W.; Cucchiarini, M. rAAV-Mediated sox9 Overexpression Improves the Repair of Osteochondral Defects in a Clinically Relevant Large Animal Model Over Time In Vivo and Reduces Perifocal Osteoarthritic Changes. Am. J. Sports Med. 2021, 49, 3696–3707. [Google Scholar] [CrossRef]
- Han, X.; Cui, J.; Chu, L.; Zhang, W.; Xie, K.; Jiang, X.; He, Z.; Du, J.; Ai, S.; Sun, Q.; et al. Abnormal subchondral trabecular bone remodeling in knee osteoarthritis under the influence of knee alignment. Osteoarthr. Cartil. 2022, 30, 100–109. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.-F.; Hu, C.-C.; Hsu, Y.-H.; Chiu, Y.-T.; Chen, K.-L.; Ueng, S.W.N.; Chang, Y. Characterization and Advancement of an Evaluation Method for the Treatment of Spontaneous Osteoarthritis in STR/ort Mice: GRGDS Peptides as a Potential Treatment for Osteoarthritis. Biomedicines 2023, 11, 1111. https://doi.org/10.3390/biomedicines11041111
Chen M-F, Hu C-C, Hsu Y-H, Chiu Y-T, Chen K-L, Ueng SWN, Chang Y. Characterization and Advancement of an Evaluation Method for the Treatment of Spontaneous Osteoarthritis in STR/ort Mice: GRGDS Peptides as a Potential Treatment for Osteoarthritis. Biomedicines. 2023; 11(4):1111. https://doi.org/10.3390/biomedicines11041111
Chicago/Turabian StyleChen, Mei-Feng, Chih-Chien Hu, Yung-Heng Hsu, Yu-Tien Chiu, Kai-Lin Chen, Steve W. N. Ueng, and Yuhan Chang. 2023. "Characterization and Advancement of an Evaluation Method for the Treatment of Spontaneous Osteoarthritis in STR/ort Mice: GRGDS Peptides as a Potential Treatment for Osteoarthritis" Biomedicines 11, no. 4: 1111. https://doi.org/10.3390/biomedicines11041111