
LncRNA SNHG4 Modulates EMT Signal and Antitumor Effects in Endometrial Cancer through Transcription Factor SP-1
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Serum Sample
2.3. Transfection of SNHG4
2.4. Trans-Well Migration Assay
2.5. Matrigel Invasion Assay
2.6. Colony Formation
2.7. Cell Proliferation Assay
2.8. Cell Cycle Assay
2.9. Real-Time PCR
2.10. Transcription Factor (TF) Binding Motif
2.11. Western Blotting Analysis
2.12. Xenograft
2.13. Hematoxylin and Eosin (H&E) Staining
2.14. Statistical Analysis
3. Results
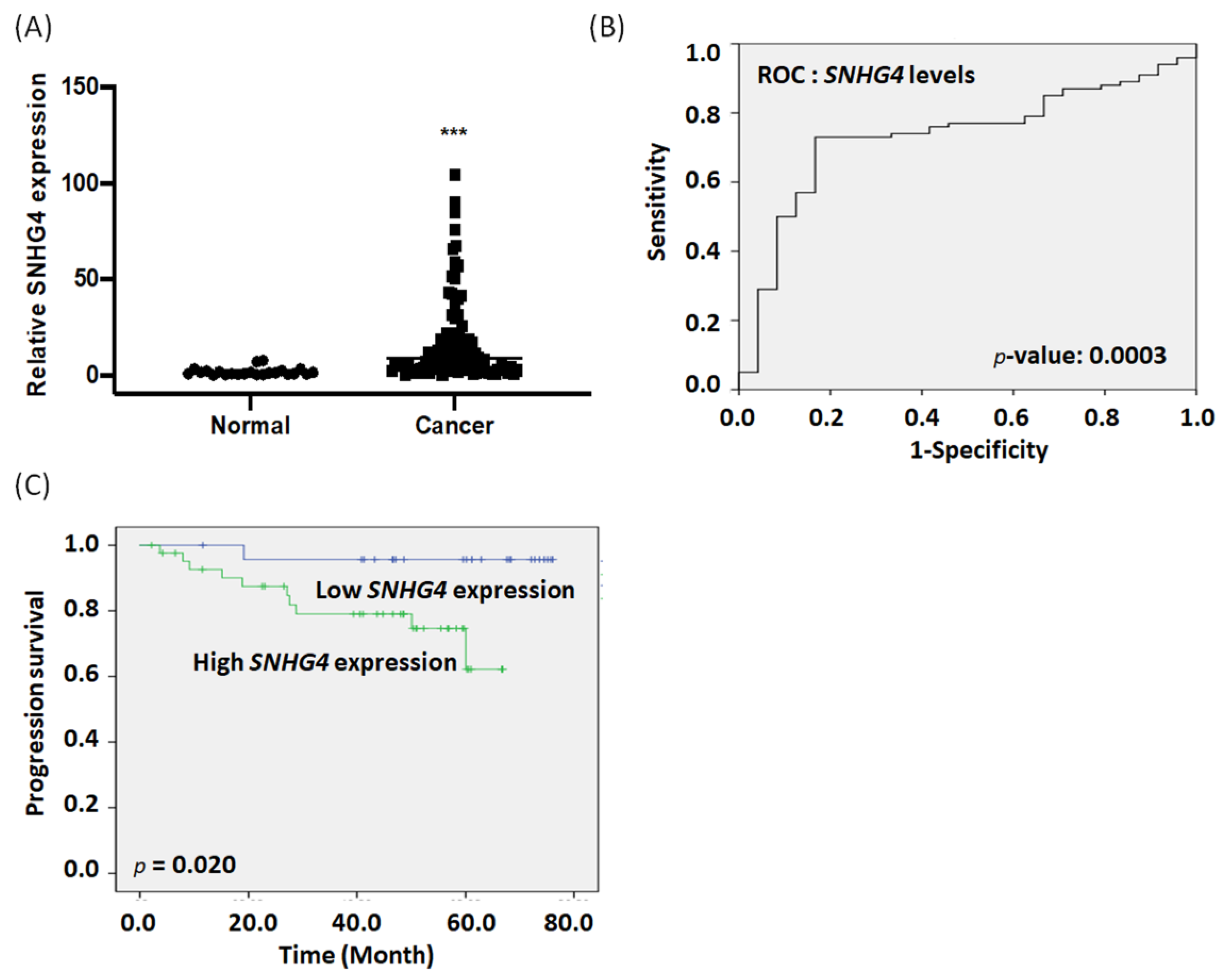
3.1. Expression of SNHG4 in Endometrial Cancer Serum Correlates with Poor Prognosis
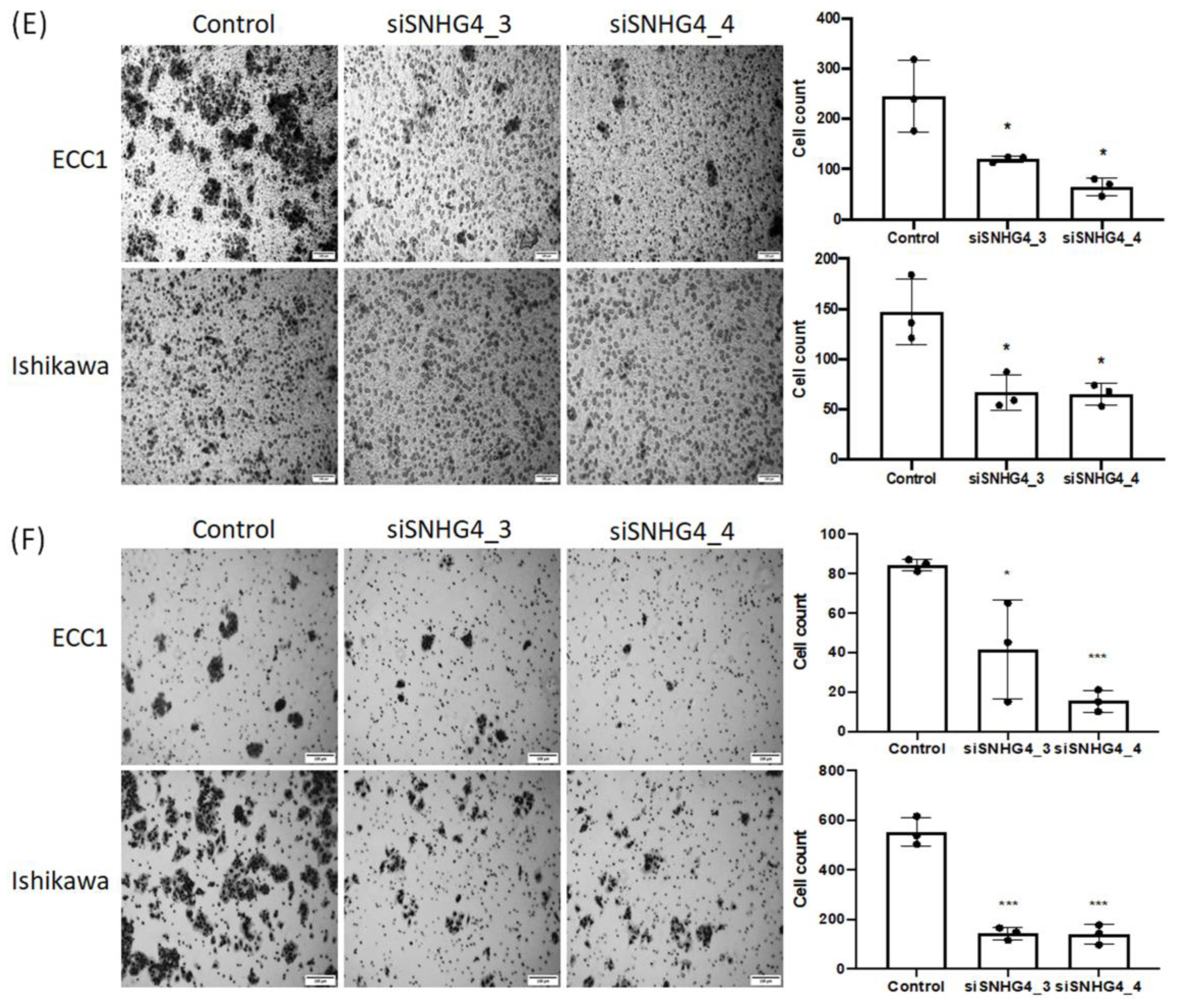
3.2. SNHG4 Expression Is Increased in Endometrial Cancer Cell Lines and Correlates with Cell Proliferation, Invasion, Migration, and Colony Formation
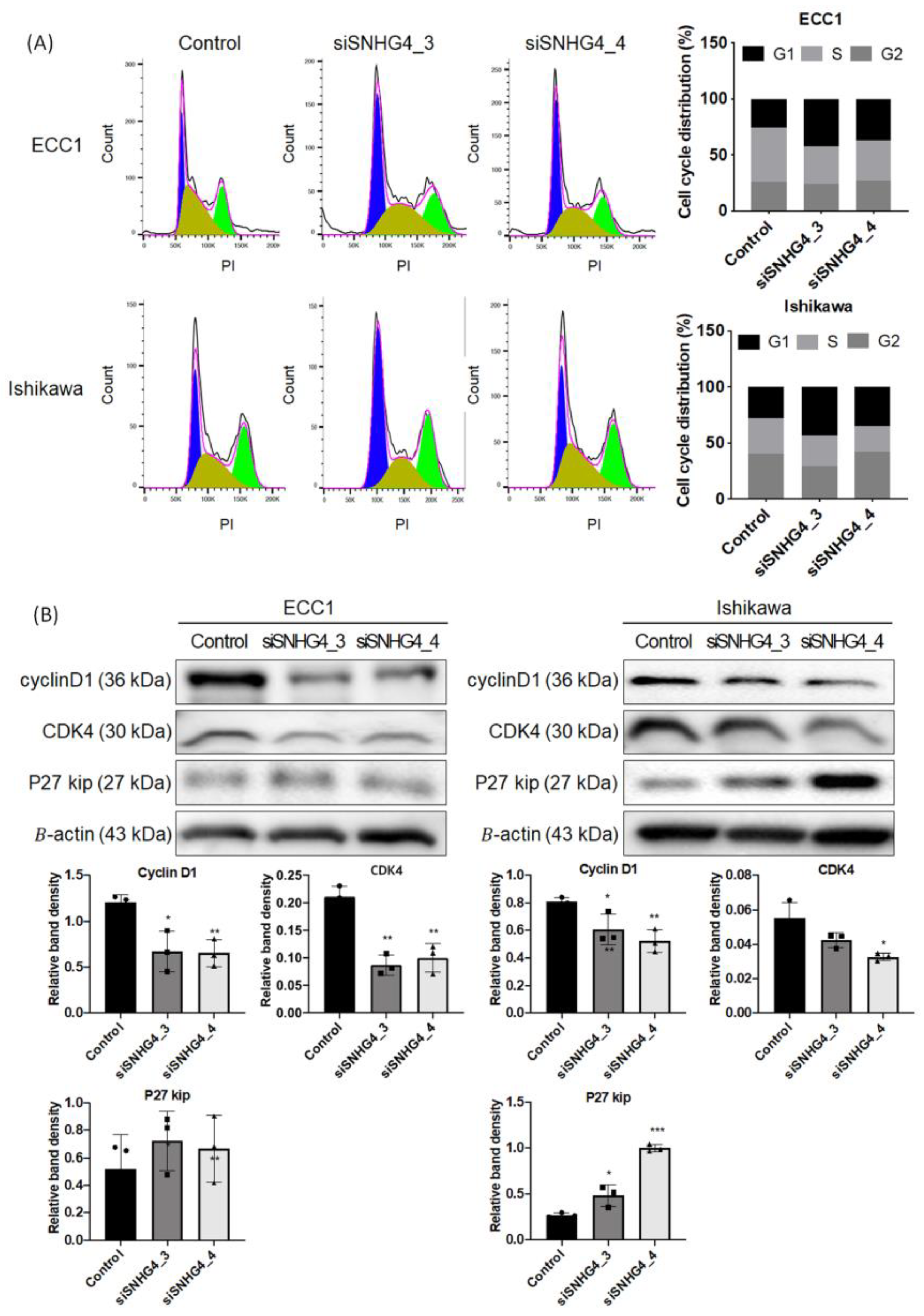
3.3. SNHG4 Knock-Down Regulates the Cell Cycle in the G1 Phase
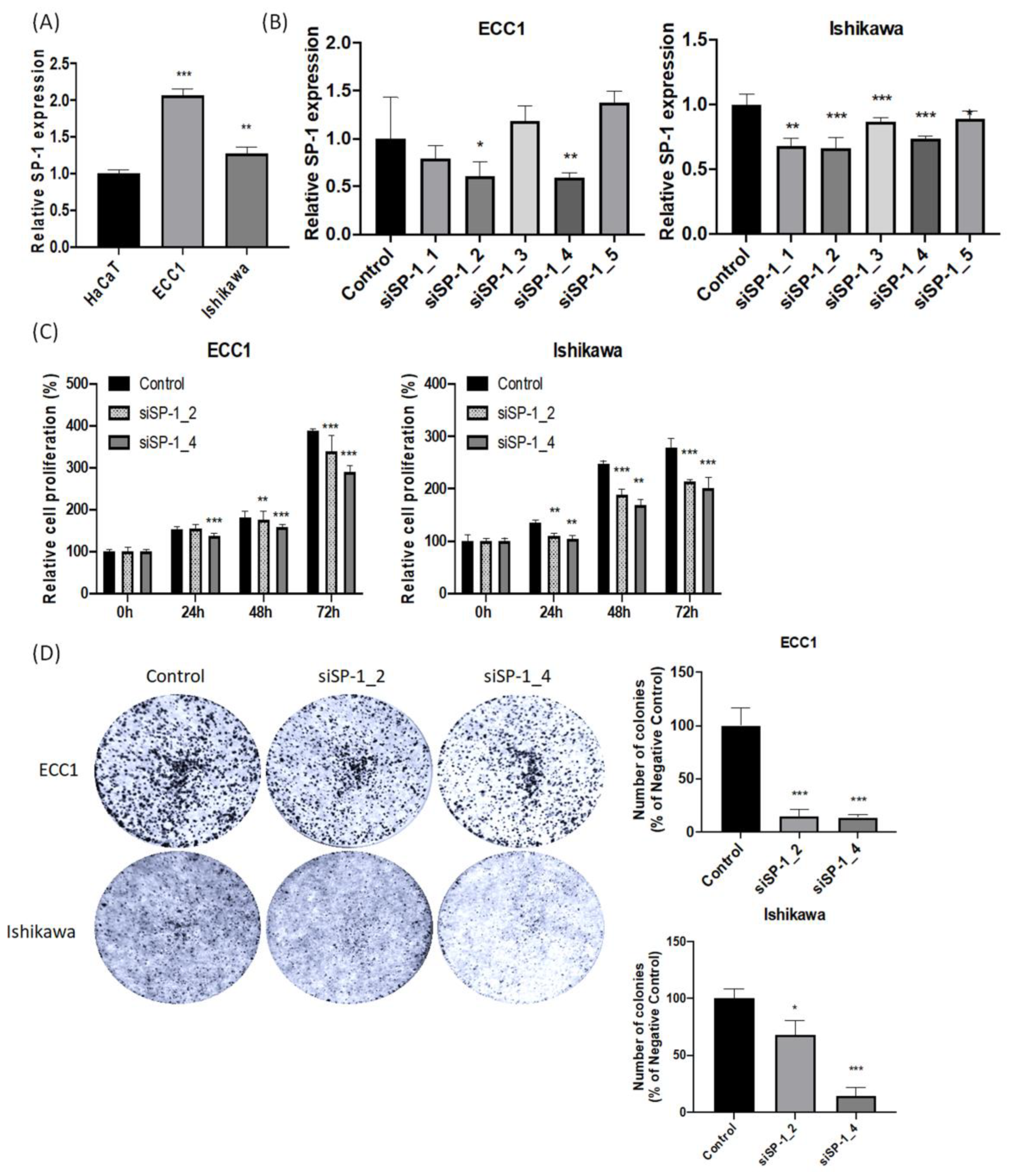
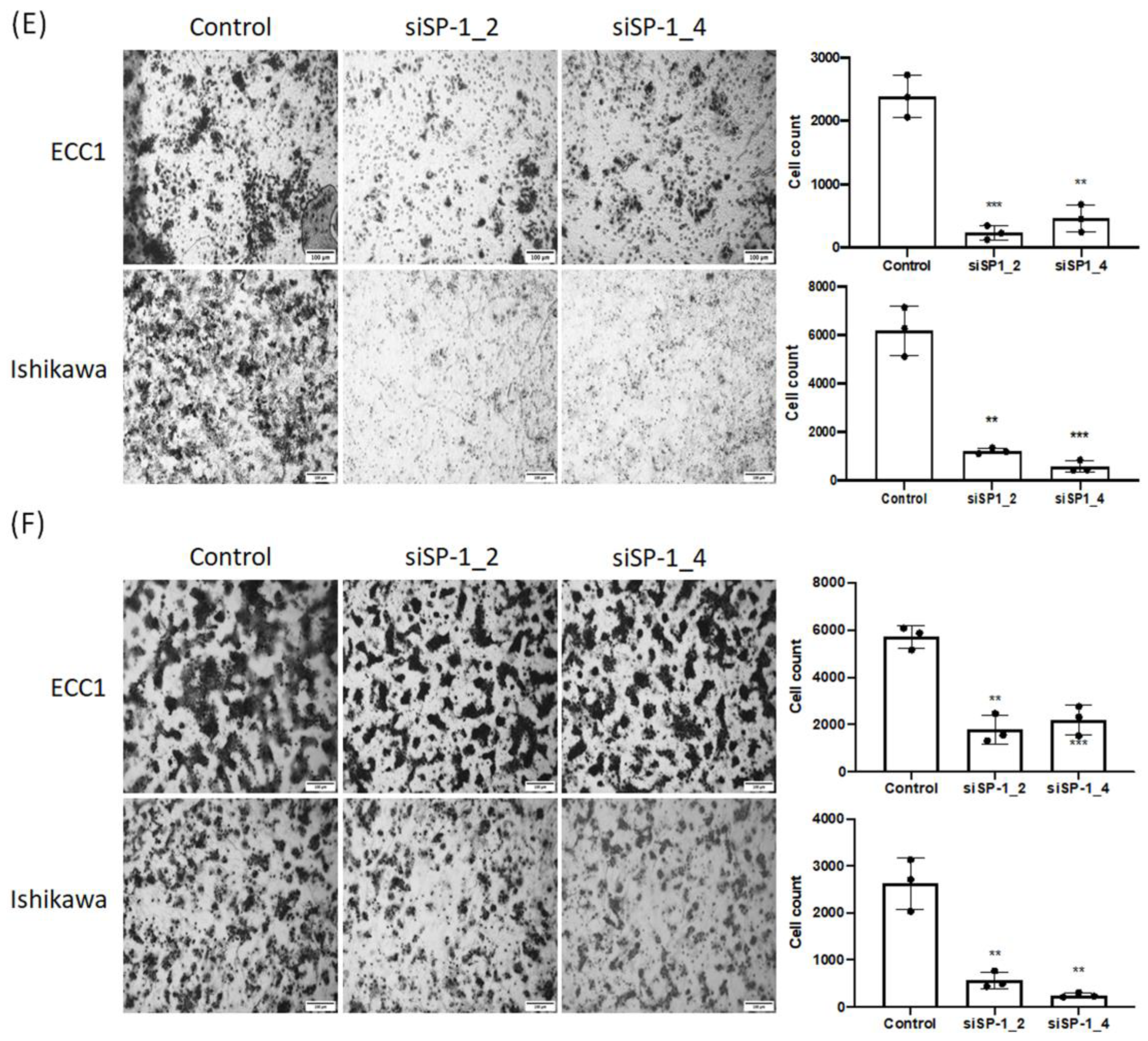
3.4. SP-1 Expression Is Increased in Endometrial Cancer Cell Lines and Correlates with Cell Proliferation, Invasion, Migration, and Colony Formation
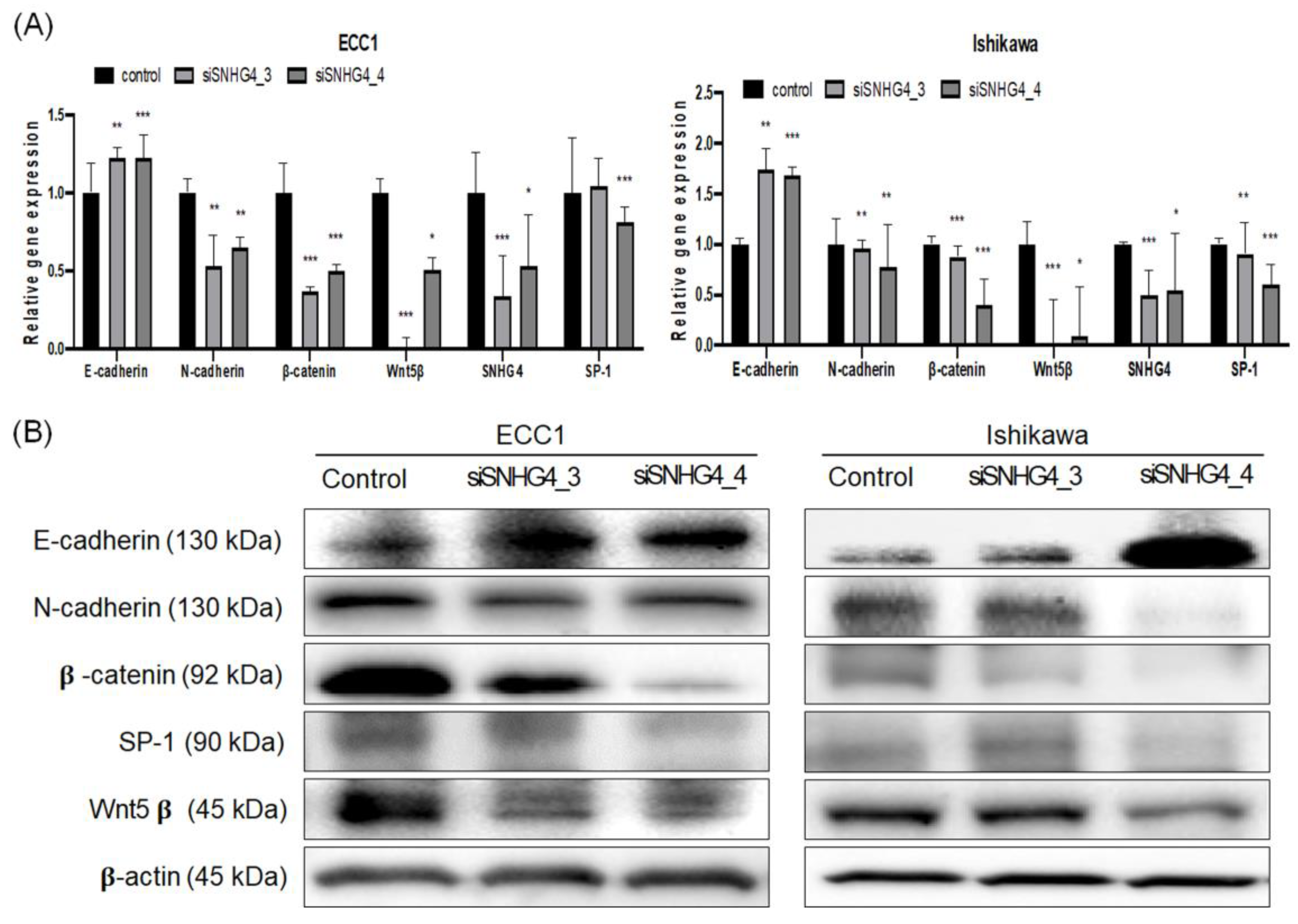
3.5. SNHG4 Regulated EMT and SP-1 Expression in Endometrial Cancer Cells
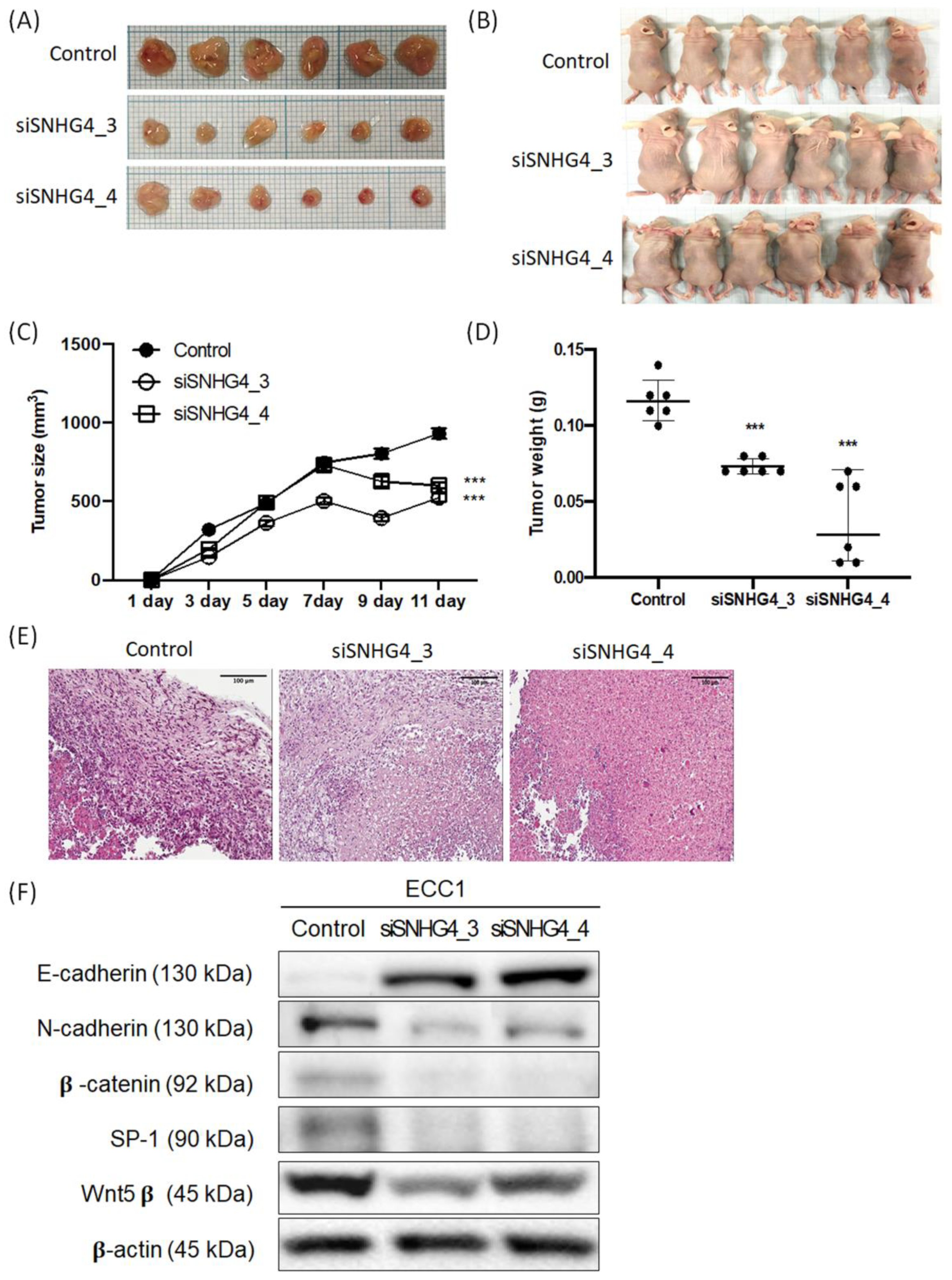
3.6. SNHG4 Regulates Tumor Growth in a Xenograft Nude Mouse Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silva, J.L.; Paulino, E.; Dias, M.F.; Melo, A.C. Endometrial cancer: Redefining the molecular-targeted approach. Cancer Chemother. Pharm. 2015, 76, 1–11. [Google Scholar] [CrossRef]
- Colombo, N.; Creutzberg, C.; Amant, F.; Bosse, T.; González-Martín, A.; Ledermann, J.; Marth, C.; Nout, R.; Querleu, D.; Mirza, M.R.; et al. ESMO-ESGO-ESTRO Consensus Conference on Endometrial Cancer: Diagnosis, treatment and follow-up. Ann. Oncol. 2016, 27, 16–41. [Google Scholar] [CrossRef]
- Piulats, J.M.; Guerra, E.; Gil-Martín, M.; Roman-Canal, B.; Gatius, S.; Sanz-Pamplona, R.; Velasco, A.; Vidal, A.; Matias-Guiu, X. Molecular approaches for classifying endometrial carcinoma. Gynecol. Oncol. 2017, 145, 200–207. [Google Scholar] [CrossRef]
- Morice, P.; Leary, A.; Creutzberg, C.; Abu-Rustum, N.; Darai, E. Endometrial cancer. Lancet 2016, 387, 1094–1108. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Xiao, M.; Feng, Y.; Liu, C.; Zhang, Z. Prognostic values of long noncoding RNA PVT 1 in various carcinomas: An updated systematic review and meta-analysis. Cell Prolif. 2018, 51, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Cen, X.; Huang, X.-Q.; Sun, W.-T.; Liu, Q.; Liu, J. Long noncoding RNAs: A new regulatory code in osteoarthritis. Am. J. Transl. Res. 2017, 9, 4747–4755. [Google Scholar]
- Nagano, T.; Fraser, P. No-Nonsense Functions for Long Noncoding RNAs. Cell 2011, 145, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Sun, W.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Zhang, H. Long Non-Coding Small Nucleolar RNA Host Genes (SNHGs) in Endocrine-Related Cancers. Onco Targets Ther. 2020, 13, 7699–7717. [Google Scholar] [CrossRef]
- Gong, J.; Li, Y.; Liu, C.J.; Xiang, Y.; Li, C.; Ye, Y.; Zhang, Z.; Hawke, D.H.; Park, P.K.; Diao, L.; et al. A Pan-cancer Analysis of the Expression and Clinical Relevance of Small Nucleolar RNAs in Human Cancer. Cell Rep. 2017, 21, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Jiang, Z.; Wang, S.; Zhao, Y.; Song, X.; Xiao, Y.; Yang, S. Long non-coding small nucleolar RNA host genes in digestive cancers. Cancer Med. 2019, 8, 7693–7704. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Wang, W. LncRNA SNHG4 regulates miR-10a/PTEN to inhibit the proliferation of acute myeloid leukemia cells. Hematology 2020, 25, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Mahalaxmi, I.; Santhy, K.S. Role and hallmarks of Sp1 in promoting ovarian cancer. J. Oncol. Sci. 2018, 4, 102–105. [Google Scholar] [CrossRef]
- Mir, R.; Sharma, A.; Pradhan, S.J.; Galande, S. Regulation of transcription factor SP1 by β-catenin destruction complex modulates Wnt response. bioRxiv 2018, 38, 308841. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, X.; Wang, Y.; Xing, L. Long Non-Coding RNA LINC00491 contributes to the malignancy of non-small-cell lung cancer via competitively binding to microRNA-324-5p and thereby increasing specificity protein 1 expression. Cancer Manag. Res. 2020, 12, 6779. [Google Scholar] [CrossRef]
- Cong, Z.; Diao, Y.; Xu, Y.; Li, X.; Jiang, Z.; Shao, C.; Ji, S.; Shen, Y.; De, W.; Qiang, Y. Long non-coding RNA linc00665 promotes lung adenocarcinoma progression and functions as ceRNA to regulate AKR1B10-ERK signaling by sponging miR-98. Cell Death Dis. 2019, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Wang, W.; Mo, S.; Chen, R.; Zou, K.; Han, J.; Zhang, F.; Hu, J. SP1-induced lncRNA AGAP2-AS1 expression promotes chemoresistance of breast cancer by epigenetic regulation of MyD88. J. Exp. Clin. Cancer Res. 2018, 37, 202. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Chen, X.; Lin, K.; Zeng, K.; Liu, X.; Xu, X.; Pan, B.; Xu, T.; Sun, L.; He, B. lncRNA SNHG6 regulates EZH2 expression by sponging miR-26a/b and miR-214 in colorectal cancer. J. Hematol. Oncol. 2019, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.T.; Farzaneh, F. Are snoRNAs and snoRNA host genes new players in cancer? Nat. Rev. Cancer 2012, 12, 84–88. [Google Scholar] [CrossRef]
- Li, Y.-H.; Hu, Y.-Q.; Wang, S.-C.; Li, Y.; Chen, D.-M. LncRNA SNHG5: A new budding star in human cancers. Gene 2020, 749, 144724. [Google Scholar] [CrossRef]
- Zhou, Y.; Tian, B.; Tang, J.; Wu, J.; Wang, H.; Wu, Z.; Li, X.; Yang, D.; Zhang, B.; Xiao, Y. SNHG7: A novel vital oncogenic lncRNA in human cancers. Biomed. Pharmacother. 2020, 124, 109921. [Google Scholar] [CrossRef]
- Tamang, S.; Acharya, V.; Roy, D.; Sharma, R.; Aryaa, A.; Sharma, U.; Khandelwal, A.; Prakash, H.; Vasquez, K.M.; Jain, A. SNHG12: An LncRNA as a Potential Therapeutic Target and Biomarker for Human Cancer. Front. Oncol. 2019, 9, 901. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiao, T.; Ou, W.; Wu, Z.; Wu, J.; Tang, J.; Tian, B.; Zhou, Y.; Su, M.; Wang, W. LncRNA SNHG16 as a potential biomarker and therapeutic target in human cancers. Biomark. Res. 2020, 8, 42. [Google Scholar] [CrossRef]
- Malek, R.; Wang, H.; Taparra, K.; Tran, P.T. Therapeutic Targeting of Epithelial Plasticity Programs: Focus on the Epithelial-Mesenchymal Transition. Cells Tissues Organs 2017, 203, 114–127. [Google Scholar] [CrossRef]
- Zaravinos, A. The Regulatory Role of MicroRNAs in EMT and Cancer. J. Oncol. 2015, 2015, 865816. [Google Scholar] [CrossRef] [Green Version]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Sun, Q.; Hao, Q.; Prasanth, K.V. Nuclear long noncoding RNAs: Key regulators of gene expression. Trends Genet. 2018, 34, 142–157. [Google Scholar] [CrossRef]
- Liu, G.; Ye, Z.; Zhao, X.; Ji, Z. SP1-induced up-regulation of lncRNA SNHG14 as a ceRNA promotes migration and invasion of clear cell renal cell carcinoma by regulating N-WASP. Am. J. Cancer Res. 2017, 7, 2515–2525. [Google Scholar]
- Sheng, K.; Lu, J.; Zhao, H. ELK1-induced upregulation of lncRNA HOXA10-AS promotes lung adenocarcinoma progression by increasing Wnt/β-catenin signaling. Biochem. Biophys. Res. Commun. 2018, 501, 612–618. [Google Scholar] [CrossRef]
- Zhang, E.; Yin, D.; Han, L.; He, X.; Si, X.; Chen, W.; Xia, R.; Xu, T.; Gu, D.; De, W. E2F1-induced upregulation of long noncoding RNA LINC00668 predicts a poor prognosis of gastric cancer and promotes cell proliferation through epigenetically silencing of CKIs. Oncotarget 2016, 7, 23212–23226. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-A.; Kim, L.K.; Kim, Y.T.; Heo, T.-H.; Kim, H.J. Long non-coding RNA steroid receptor activator promotes the progression of endometrial cancer via Wnt/β-catenin signaling pathway. Int. J. Biol. Sci. 2020, 16, 99–115. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-A.; Kim, L.K.; Kim, Y.T.; Heo, T.-H.; Kim, H.J. Long Noncoding RNA E2F4as Promotes Progression and Predicts Patient Prognosis in Human Ovarian Cancer. Cancers 2020, 12, 3626. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Duan, Y.; Wang, P. SP1-mediated upregulation of lncRNA SNHG4 functions as a ceRNA for miR-377 to facilitate prostate cancer progression through regulation of ZIC5. J. Cell Physiol. 2020, 235, 3916–3927. [Google Scholar] [CrossRef]
- López-Sáez, J.F.; De la Torre, C.; Pincheira, J.; Martín, G.G. Cell proliferation and cancer. Histol. Histopathol. 1998, 13, 1197–1214. [Google Scholar]
- Ersvær, E.; Kildal, W.; Vlatkovic, L.; Cyll, K.; Pradhan, M.; Kleppe, A.; Hveem, T.S.; Askautrud, H.A.; Novelli, M.; Wæhre, H. Prognostic value of mitotic checkpoint protein BUB3, cyclin B1, and pituitary tumor-transforming 1 expression in prostate cancer. Mod. Pathol. 2020, 33, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Qie, S.; Diehl, J.A. Cyclin D1, cancer progression, and opportunities in cancer treatment. J. Mol. Med. 2016, 94, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Landis, M.W.; Pawlyk, B.S.; Li, T.; Sicinski, P.; Hinds, P.W. Cyclin D1-dependent kinase activity in murine development and mammary tumorigenesis. Cancer Cell 2006, 9, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Chu, Q.; Gu, X.; Zheng, Q.; Guo, Z.; Shan, D.; Wang, J.; Zhu, H. Long noncoding RNA SNHG4: A novel target in human diseases. Cancer Cell Int. 2021, 21, 583. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PFS | ||||
|---|---|---|---|---|
| Univariate Analysis | Multivariate Analysis | |||
| HR (95% CI) | p-Value | HR (95% CI) | p-Value | |
| SNHG4 expression | 8.102 (1.014–64.758) | 0.049 | 4.851 (0.538–43.757) | 0.159 |
| Age, years (continuous) | 1.021 (0.956–1.091) | 0.529 | 1.028 (0.932–1.134) | 0.58 |
| FIGO stage | 2.462 (1.347–4.502) | 0.003 | 2.079 (0.517–8.350) | 0.302 |
| Grade | 1.374 (0.662–2.852) | 0.394 | 1.019 (0.371–2.802) | 0.97 |
| Lymph node metastasis | 8.138 (2.153–30.757) | 0.002 | 3.068 (0.248–38.013) | 0.383 |
| Lymphovascular invasion | 2.927 (0.855–10.024) | 0.087 | 0.376 (0.069–2.046) | 0.258 |
| BMI | 0.541 (0.158–1.853) | 0.328 | 0.438 (0.095–2.022) | 0.29 |
| Tumor size | 2.170 (0.920–5.120) | 0.077 | 1.086 (0.362–3.257) | 0.883 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, L.K.; Park, S.-A.; Nam, E.J.; Kim, Y.T.; Heo, T.-H.; Kim, H.J. LncRNA SNHG4 Modulates EMT Signal and Antitumor Effects in Endometrial Cancer through Transcription Factor SP-1. Biomedicines 2023, 11, 1018. https://doi.org/10.3390/biomedicines11041018
Kim LK, Park S-A, Nam EJ, Kim YT, Heo T-H, Kim HJ. LncRNA SNHG4 Modulates EMT Signal and Antitumor Effects in Endometrial Cancer through Transcription Factor SP-1. Biomedicines. 2023; 11(4):1018. https://doi.org/10.3390/biomedicines11041018
Chicago/Turabian StyleKim, Lee Kyung, Sun-Ae Park, Eun Ji Nam, Young Tae Kim, Tae-Hwe Heo, and Hee Jung Kim. 2023. "LncRNA SNHG4 Modulates EMT Signal and Antitumor Effects in Endometrial Cancer through Transcription Factor SP-1" Biomedicines 11, no. 4: 1018. https://doi.org/10.3390/biomedicines11041018