
Growth Conditions Influence Lactobacillus Cell-Free Supernatant Impact on Viability, Biofilm Formation, and Co-Aggregation of the Oral Periodontopathogens Fusobacterium nucleatum and Porphyromonas gingivalis

 ,
,  ,
,  , and
, and 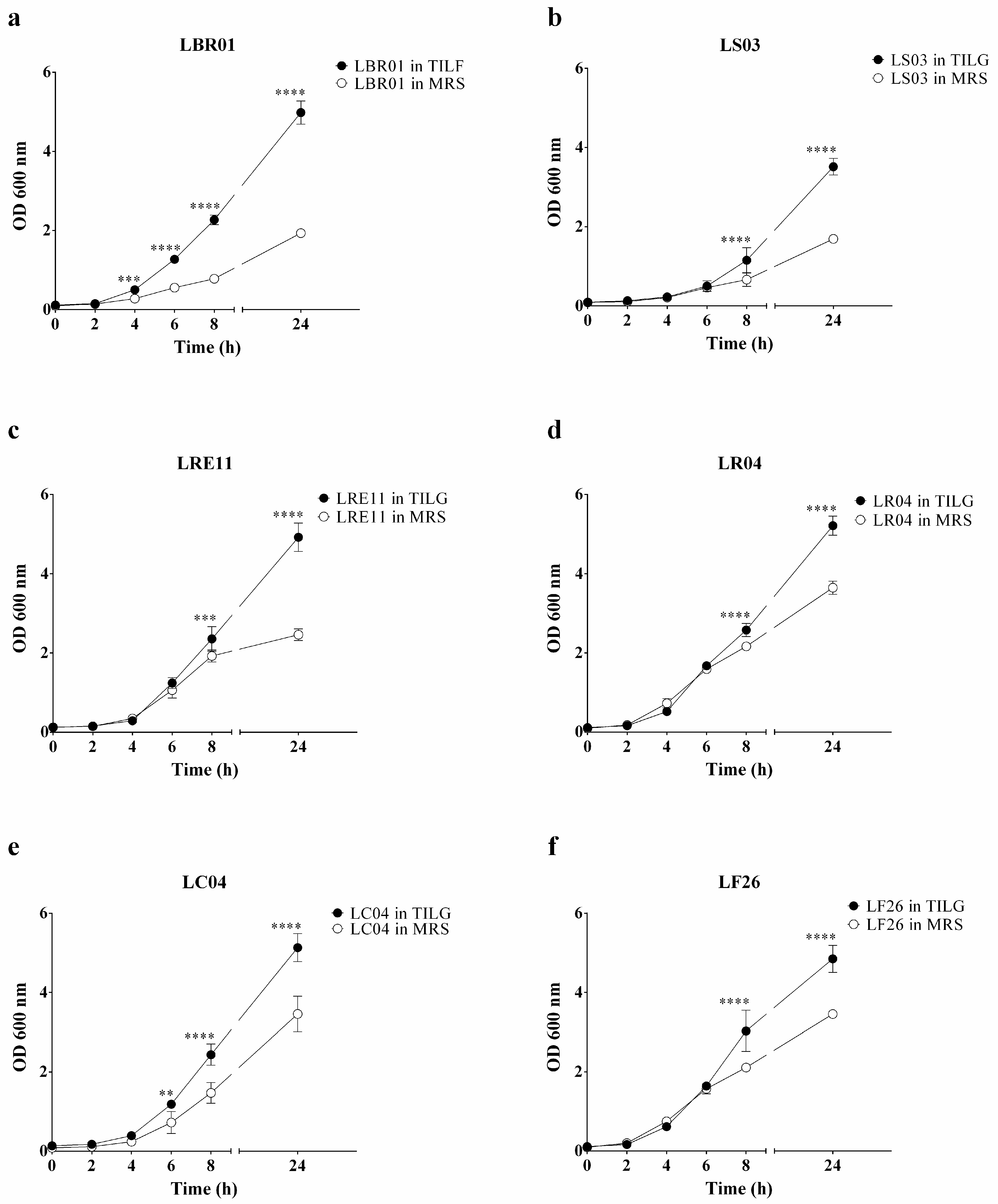{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures and Growth Curves
2.2. Probiotic Cell-Free Supernatant Production
2.3. Viability Assay
2.4. Biofilm Formation Assay
2.5. Co-Aggregation Assay
2.6. Statistical Analysis
3. Results and Discussion
3.1. Probiotic Growth Curves
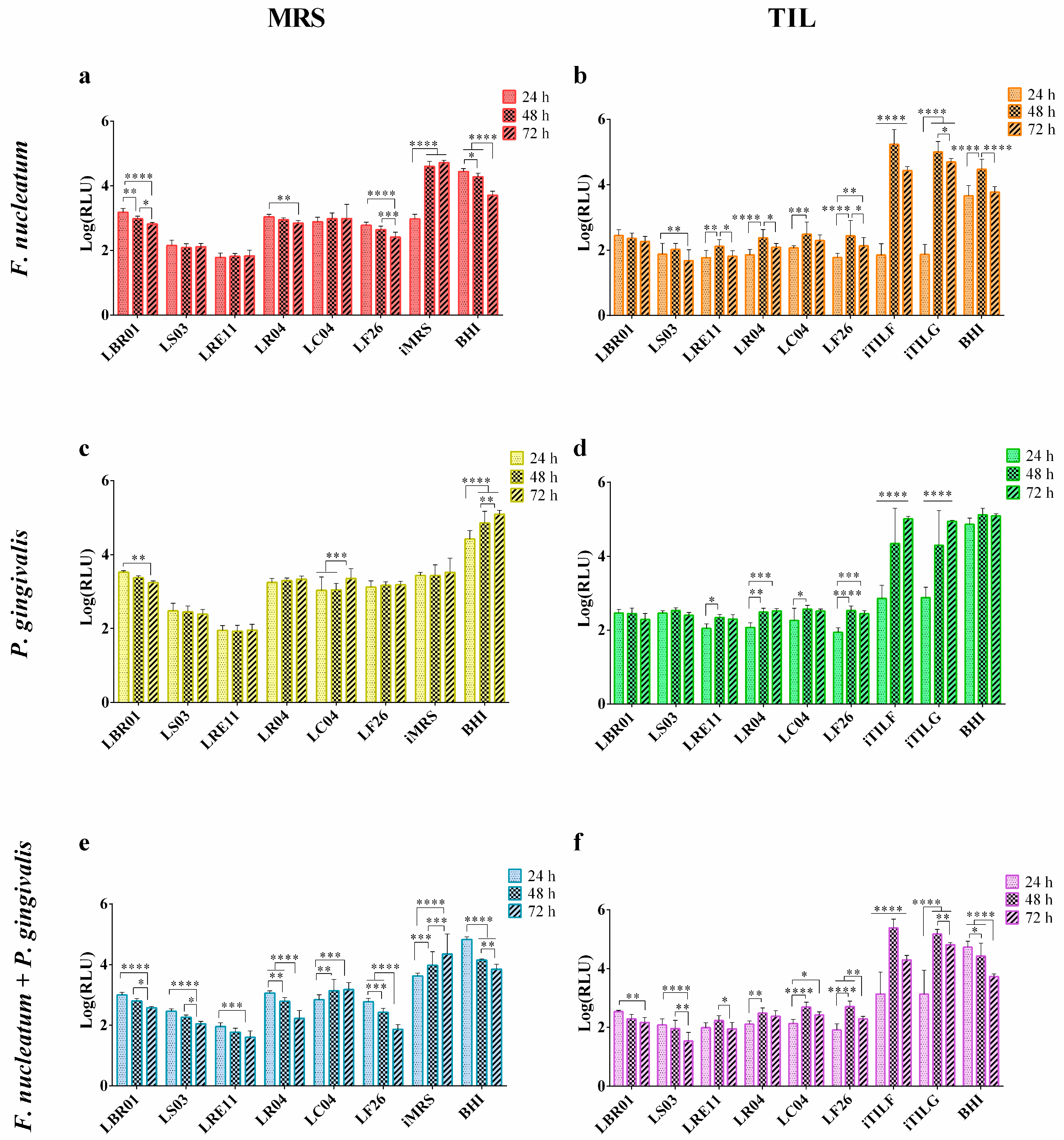
3.2. Viability Assay
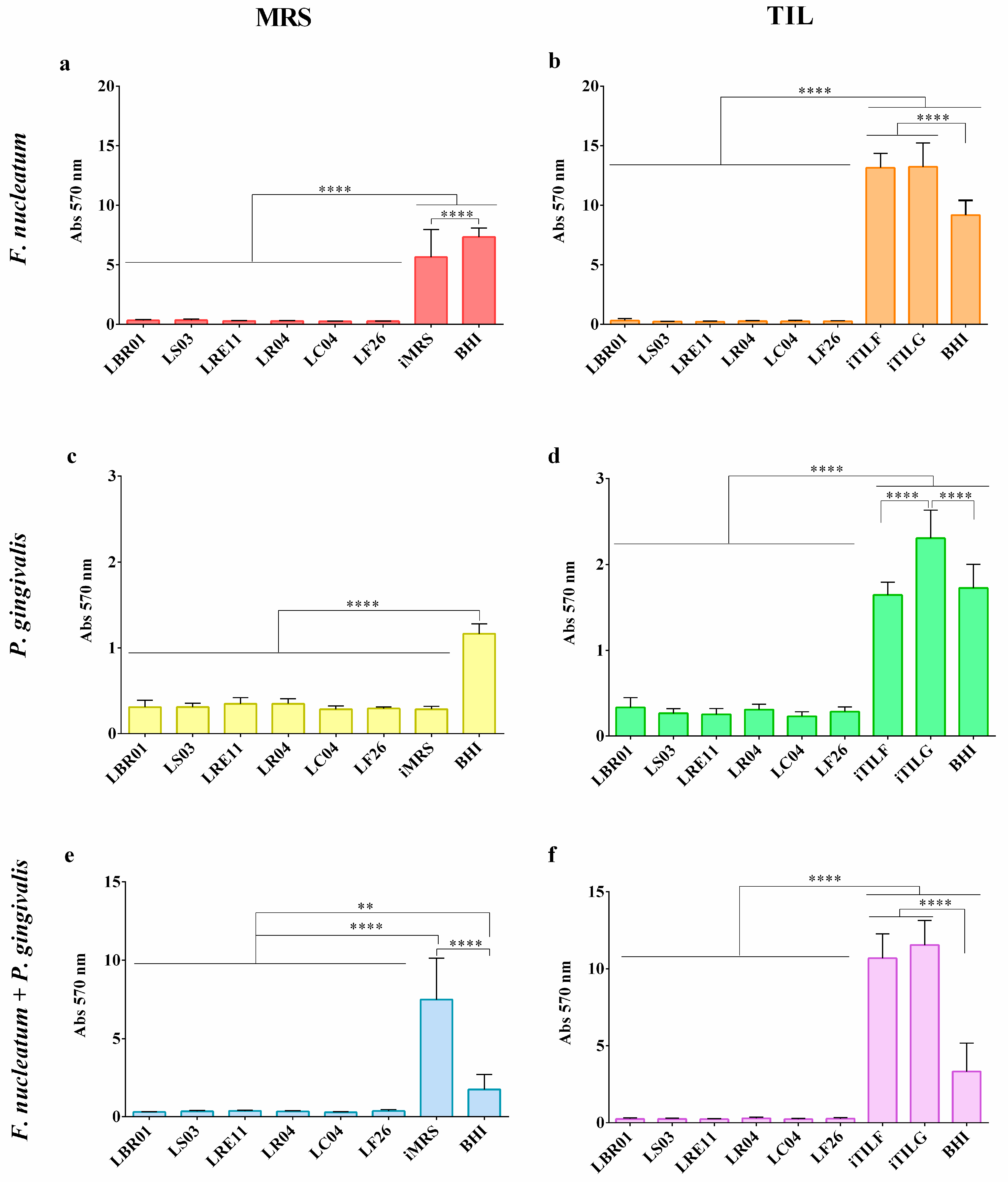
3.3. Biofilm Formation Assay
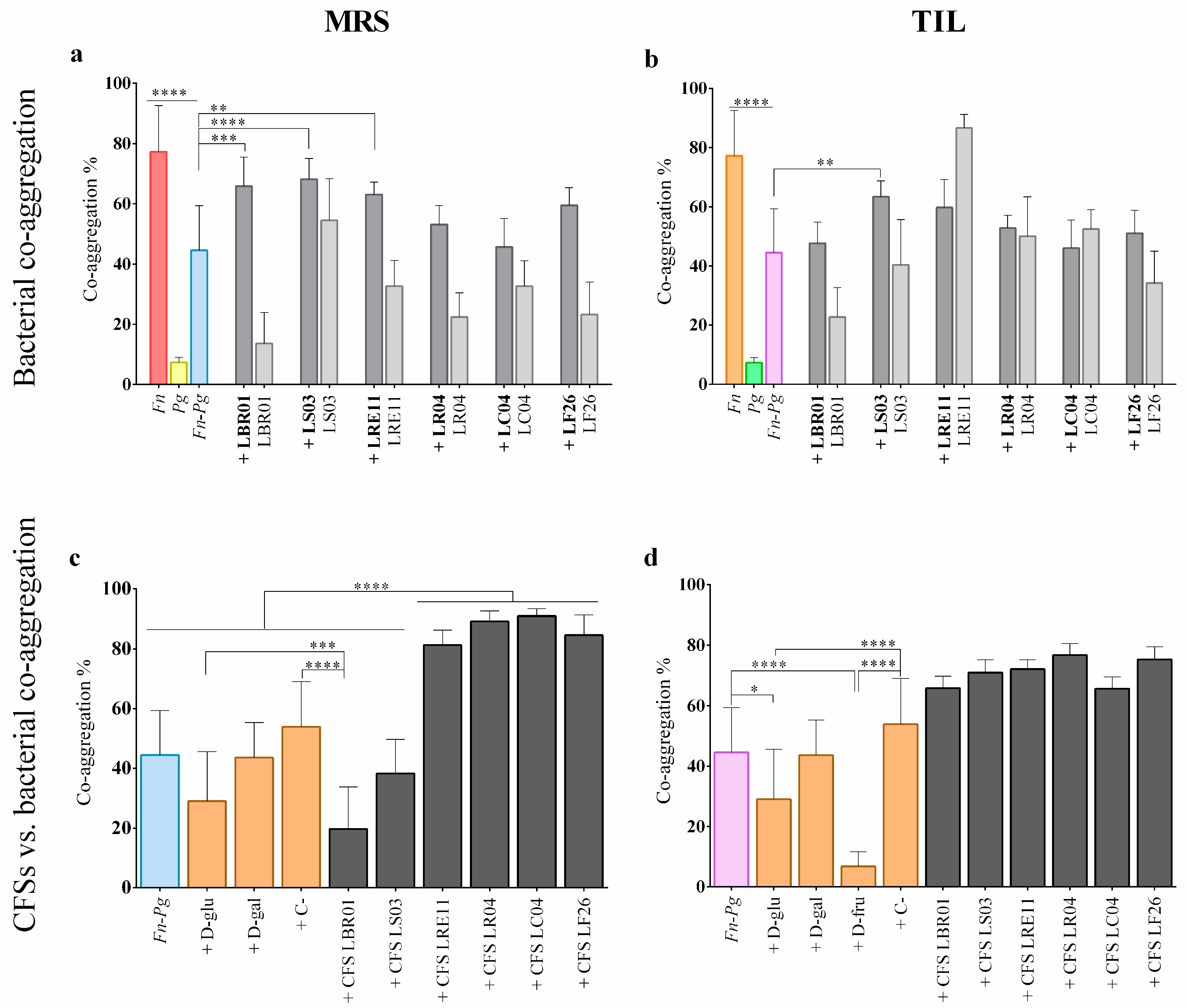
3.4. Co-Aggregation Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanetta, P.; Squarzanti, D.F.; Sorrentino, R.; Rolla, R.; Aluffi Valletti, P.; Garzaro, M.; Dell’Era, V.; Amoruso, A.; Azzimonti, B. Oral microbiota and vitamin D impact on oropharyngeal squamous cell carcinogenesis: A narrative literature review. Crit. Rev. Microbiol. 2021, 47, 224–239. [Google Scholar] [CrossRef]
- Zhao, T.; Chen, J.; Liu, S.; Yang, J.; Wu, J.; Miao, L.; Sun, W. Transcriptome analysis of Fusobacterium nucleatum reveals differential gene expression patterns in the biofilm versus planktonic cells. Biochem. Biophys. Res. Commun. 2022, 593, 151–157. [Google Scholar] [CrossRef]
- Katz, J.; Onate, M.D.; Pauley, K.M.; Bhattacharyya, I.; Cha, S. Presence of Porphyromonas gingivalis in gingival squamous cell carcinoma. Int. J. Oral Sci. 2011, 3, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Coppenhagen-Glazer, S.; Sol, A.; Abed, J.; Naor, R.; Zhang, X.; Han, Y.W.; Bachrach, G. Fap2 of Fusobacterium nucleatum is a galactose-inhibitable adhesin involved in coaggregation, cell adhesion, and preterm birth. Infect. Immun. 2015, 83, 1104–1113. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, H.; Wang, X.; Lu, Y.; Yang, C.; Yang, P. Coinfection with Fusobacterium nucleatum can enhance the attachment and invasion of Porphyromonas gingivalis or Aggregatibacter actinomycetemcomitans to human gingival epithelial cells. Arch. Oral Biol. 2015, 60, 1387–1393. [Google Scholar] [CrossRef]
- Kramer, C.D.; Genco, C.A. Microbiota, Immune Subversion, and Chronic Inflammation. Front. Immunol. 2017, 8, 255. [Google Scholar] [CrossRef] [Green Version]
- Li, T.-J.; Hao, Y.-H.; Tang, Y.-L.; Liang, X.-H. Periodontal Pathogens: A Crucial Link Between Periodontal Diseases and Oral Cancer. Front. Microbiol. 2022, 13, 919633. [Google Scholar] [CrossRef]
- Geng, F.; Zhang, Y.; Lu, Z.; Zhang, S.; Pan, Y. Fusobacterium nucleatum Caused DNA Damage and Promoted Cell Proliferation by the Ku70/p53 Pathway in Oral Cancer Cells. DNA Cell Biol. 2020, 39, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, I.; Verma, M.; Panda, M. Role of Oral Microbiome Signatures in Diagnosis and Prognosis of Oral Cancer. Technol. Cancer Res. Treat. 2019, 18, 153303381986735. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Shao, Z.; Liu, K.; Zhou, X.; Wang, L.; Jiang, E.; Luo, T.; Shang, Z. Salivary Porphyromonas gingivalis predicts outcome in oral squamous cell carcinomas: A cohort study. BMC Oral Health 2021, 21, 228. [Google Scholar] [CrossRef]
- Shillitoe, E.J. The Microbiome of Oral Cancer. Crit. Rev. Oncog. 2018, 23, 153–160. [Google Scholar] [CrossRef]
- Ardila, C.M.; Vivares-Builes, A.M. Antibiotic Resistance in Patients with Peri-Implantitis: A Systematic Scoping Review. Int. J. Environ. Res. Public Health 2022, 19, 5609. [Google Scholar] [CrossRef]
- Zanetta, P.; Squarzanti, D.F.; di Coste, A.; Rolla, R.; Valletti, P.A.; Garzaro, M.; Dell’Era, V.; Amoruso, A.; Pane, M.; Azzimonti, B. In Vitro Selection of Lactobacillus and Bifidobacterium Probiotic Strains for the Management of Oral Pathobiont Infections Associated to Systemic Diseases. Int. J. Mol. Sci. 2022, 23, 6163. [Google Scholar] [CrossRef]
- Datta, A.; Stapleton, F.; Willcox, M.D.P. Bacterial Coaggregation Among the Most Commonly Isolated Bacteria From Contact Lens Cases. Investig. Ophthalmol. Vis. Sci. 2017, 58, 50–58. [Google Scholar] [CrossRef]
- Squarzanti, D.F.; Zanetta, P.; Ormelli, M.; Manfredi, M.; Barberis, E.; Vanella, V.V.; Amoruso, A.; Pane, M.; Azzimonti, B. An animal derivative-free medium enhances Lactobacillus johnsonii LJO02 supernatant selective efficacy against the methicillin (oxacillin)-resistant Staphylococcus aureus virulence through key-metabolites. Sci. Rep. 2022, 12, 8666. [Google Scholar] [CrossRef]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.; Bhat, S.G.; Sivaraman, K. Porphyromonas gingivalis adopts intricate and unique molecular mechanisms to survive and persist within the host: A critical update. J. Oral Microbiol. 2020, 12, 1801090. [Google Scholar] [CrossRef]
- Asare, P.T.; Zurfluh, K.; Greppi, A.; Lynch, D.; Schwab, C.; Stephan, R.; Lacroix, C. Reuterin Demonstrates Potent Antimicrobial Activity Against a Broad Panel of Human and Poultry Meat Campylobacter spp. Isolates. Microorganisms 2020, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Jansen, P.M.; Abdelbary, M.M.H.; Conrads, G. A concerted probiotic activity to inhibit periodontitis-associated bacteria. PLoS ONE 2021, 16, e0248308. [Google Scholar] [CrossRef]
- Santos, T.A.; Scorzoni, L.; Correia, R.; Junqueira, J.C.; Anbinder, A.L. Interaction between Lactobacillus reuteri and periodontopathogenic bacteria using in vitro and in vivo (G. mellonella) approaches. Pathog. Dis. 2020, 78, ftaa044. [Google Scholar] [CrossRef]
- Kang, M.-S.; Oh, J.-S.; Lee, H.-C.; Lim, H.-S.; Lee, S.-W.; Yang, K.-H.; Choi, N.-K.; Kim, S.-M. Inhibitory effect of Lactobacillus reuteri on periodontopathic and cariogenic bacteria. J. Microbiol. 2011, 49, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-S.; Yum, H.; Cho, Y.-D.; Kim, S. Improvement of halitosis by probiotic bacterium Weissella cibaria CMU: A randomized controlled trial. Front. Microbiol. 2023, 14, 1108762. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.M.; Kim, J.-S.; Kim, H.-S.; Kim, Y.-Y.; Oh, J.-K.; Jung, H.-W.; Park, D.-S.; Bae, K.-H. Lactobacillus reuteri AN417 cell-free culture supernatant as a novel antibacterial agent targeting oral pathogenic bacteria. Sci. Rep. 2021, 11, 1631. [Google Scholar] [CrossRef] [PubMed]
- Mulla, M.; Mulla, M.; Hegde, S.; Koshy, A. V In vitro assessment of the effect of probiotic lactobacillus reuteri on peri-implantitis microflora. BMC Oral Health 2021, 21, 408. [Google Scholar] [CrossRef] [PubMed]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, e0230345. [Google Scholar] [CrossRef]
- Van Holm, W.; Carvalho, R.; Delanghe, L.; Eilers, T.; Zayed, N.; Mermans, F.; Bernaerts, K.; Boon, N.; Claes, I.; Lebeer, S.; et al. Antimicrobial potential of known and novel probiotics on in vitro periodontitis biofilms. NPJ Biofilms Microbiomes 2023, 9, 3. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Hsieh, P.-S.; Ho, H.-H.; Hsieh, S.-H.; Kuo, Y.-W.; Yang, S.-F.; Lin, C.-W. Antibacterial activity of viable and heat-killed probiotic strains against oral pathogens. Lett. Appl. Microbiol. 2020, 70, 310–317. [Google Scholar] [CrossRef]
- Michaylova, M.; Yungareva, T.; Urshev, Z.; Dermendzieva, Y.; Yaneva, B.; Dobrev, I. Probiotic candidates among dairy Lactobacilli and Streptococcus thermophiles strains for control of the oral pathogen Porphyromonas gingivalis. Folia Med. 2021, 63, 720–725. [Google Scholar] [CrossRef]
- Gönczi, N.N.; Strang, O.; Bagi, Z.; Rákhely, G.; Kovács, K.L. Interactions between probiotic and oral pathogenic strains. Biol. Futur. 2021, 72, 461–471. [Google Scholar] [CrossRef]
- Mann, S.; Park, M.S.; Johnston, T.V.; Ji, G.E.; Hwang, K.T.; Ku, S. Isolation, Characterization and Biosafety Evaluation of Lactobacillus Fermentum OK with Potential Oral Probiotic Properties. Probiotics Antimicrob. Proteins 2021, 13, 1363–1386. [Google Scholar] [CrossRef]
- Chen, L.-J.; Tsai, H.-T.; Chen, W.-J.; Hsieh, C.-Y.; Wang, P.-C.; Chen, C.-S.; Wang, L.; Yang, C.-C. In vitro antagonistic growth effects of Lactobacillus fermentum and lactobacillus salivarius and their fermentative broth on periodontal pathogens. Braz. J. Microbiol. 2012, 43, 1376–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.K.; Hasslöf, P.; Stecksén-Blicks, C.; Twetman, S. Co-aggregation and growth inhibition of probiotic lactobacilli and clinical isolates of mutans streptococci: An in vitro study. Acta Odontol. Scand. 2011, 69, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.H.; Mita, D.; Kawamoto, D.; Nicoli, J.R.; Albuquerque-Souza, E.; Lorenzetti Simionato, M.R.; Mayer, M.P.A. Probiotics alter biofilm formation and the transcription of Porphyromonas gingivalis virulence-associated genes. J. Oral Microbiol. 2020, 12, 1805553. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J. Cell. Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef] [Green Version]
- Scillato, M.; Spitale, A.; Mongelli, G.; Privitera, G.F.; Mangano, K.; Cianci, A.; Stefani, S.; Santagati, M. Antimicrobial properties of Lactobacillus cell-free supernatants against multidrug-resistant urogenital pathogens. Microbiologyopen 2021, 10, e1173. [Google Scholar] [CrossRef]
- Malfa, P.; Brambilla, L.; Giardina, S.; Masciarelli, M.; Squarzanti, D.F.; Carlomagno, F.; Meloni, M. Evaluation of Antimicrobial, Antiadhesive and Co-Aggregation Activity of a Multi-Strain Probiotic Composition against Different Urogenital Pathogens. Int. J. Mol. Sci. 2023, 24, 1323. [Google Scholar] [CrossRef]
- Zakaria Gomaa, E. Antimicrobial and anti-adhesive properties of biosurfactant produced by lactobacilli isolates, biofilm formation and aggregation ability. J. Gen. Appl. Microbiol. 2013, 59, 425–436. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanetta, P.; Squarzanti, D.F.; di Coste, A.; Amoruso, A.; Pane, M.; Azzimonti, B. Growth Conditions Influence Lactobacillus Cell-Free Supernatant Impact on Viability, Biofilm Formation, and Co-Aggregation of the Oral Periodontopathogens Fusobacterium nucleatum and Porphyromonas gingivalis. Biomedicines 2023, 11, 859. https://doi.org/10.3390/biomedicines11030859
Zanetta P, Squarzanti DF, di Coste A, Amoruso A, Pane M, Azzimonti B. Growth Conditions Influence Lactobacillus Cell-Free Supernatant Impact on Viability, Biofilm Formation, and Co-Aggregation of the Oral Periodontopathogens Fusobacterium nucleatum and Porphyromonas gingivalis. Biomedicines. 2023; 11(3):859. https://doi.org/10.3390/biomedicines11030859
Chicago/Turabian StyleZanetta, Paola, Diletta Francesca Squarzanti, Alessia di Coste, Angela Amoruso, Marco Pane, and Barbara Azzimonti. 2023. "Growth Conditions Influence Lactobacillus Cell-Free Supernatant Impact on Viability, Biofilm Formation, and Co-Aggregation of the Oral Periodontopathogens Fusobacterium nucleatum and Porphyromonas gingivalis" Biomedicines 11, no. 3: 859. https://doi.org/10.3390/biomedicines11030859