
Bacterial Outer Membrane Vesicles Promote Lung Inflammatory Responses and Macrophage Activation via Multi-Signaling Pathways
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Cell Culture, Reagents, and Supplies
2.2. Mice
2.3. Animal Study
2.4. BALF and Inflammatory Cell Counts
2.5. Isolation of Lung Infiltrating Cells in Mice and Cell Counts
2.6. Histopathologic Evaluation
2.7. Outer-Membrane Vesicles (OMV) and Exosome (EXO) Isolation and Characterization
2.8. ELISA and Array Analyses
2.9. RNA Isolation, Reverse Transcription, and qRT-PCR
2.10. Macrophage Phagocytosis, ROS, and Migration Assays
2.11. Statistics
3. Results
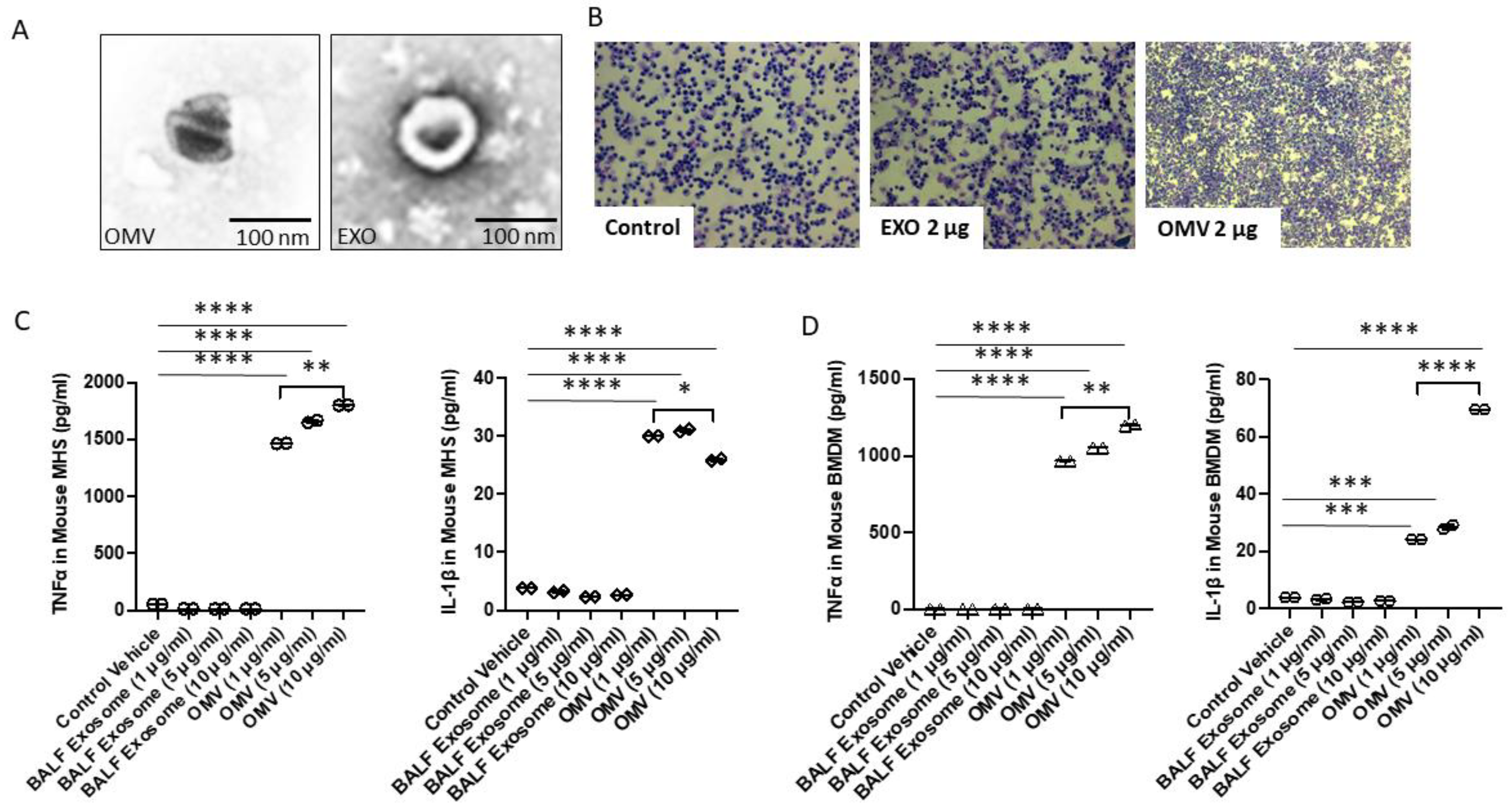
3.1. Gram-Negative Bacterial OMVs Induce Macrophage Pro-Inflammatory Activations
3.2. Gram-Negative Bacterial OMVs Induce Cytokine/Chemokine Release from Both Systemic and Lung Macrophages
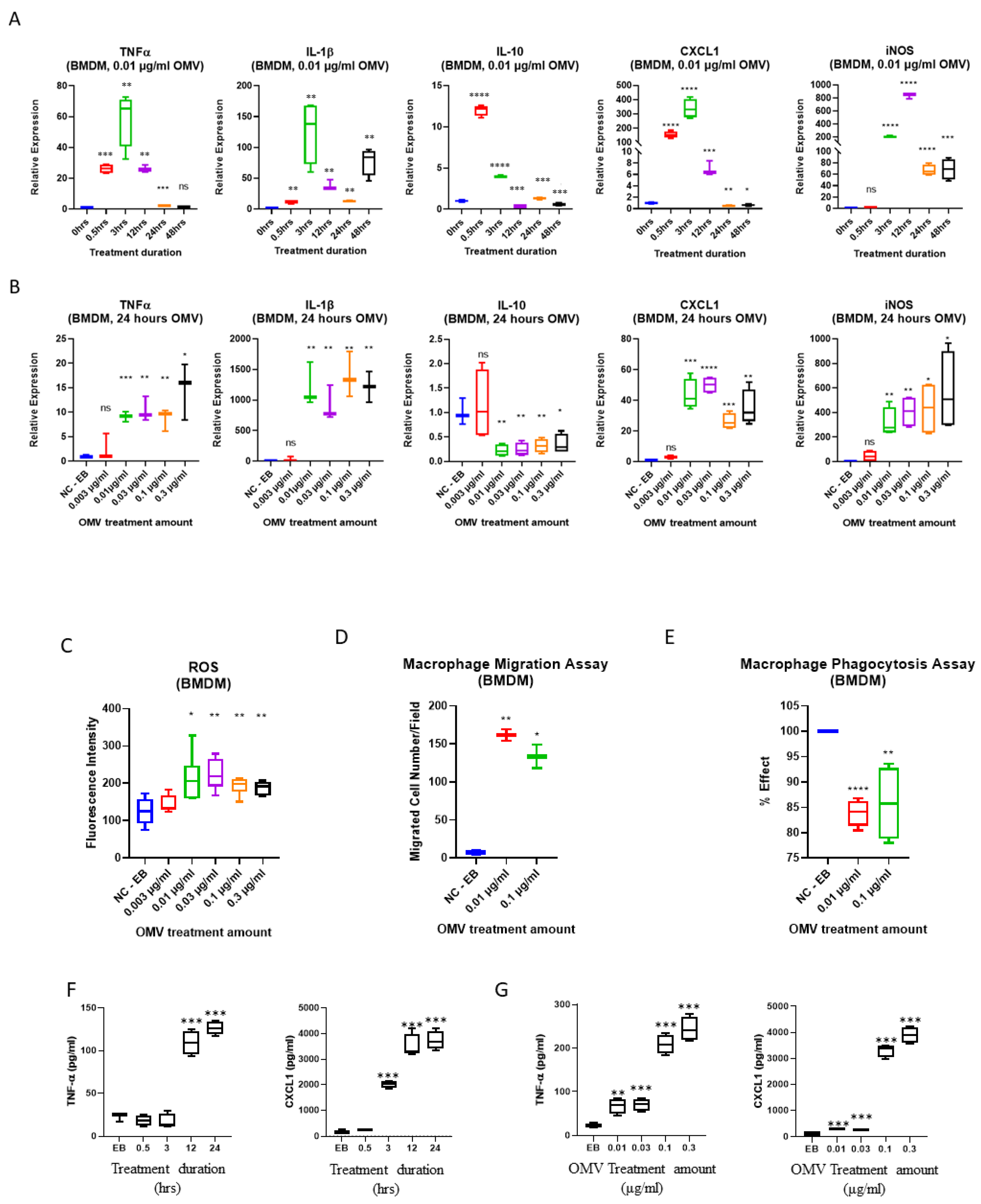
3.3. Gram-Negative Bacterial OMVs Induce Cell Migration, Phagocytosis, ROS Generation, and NO Production
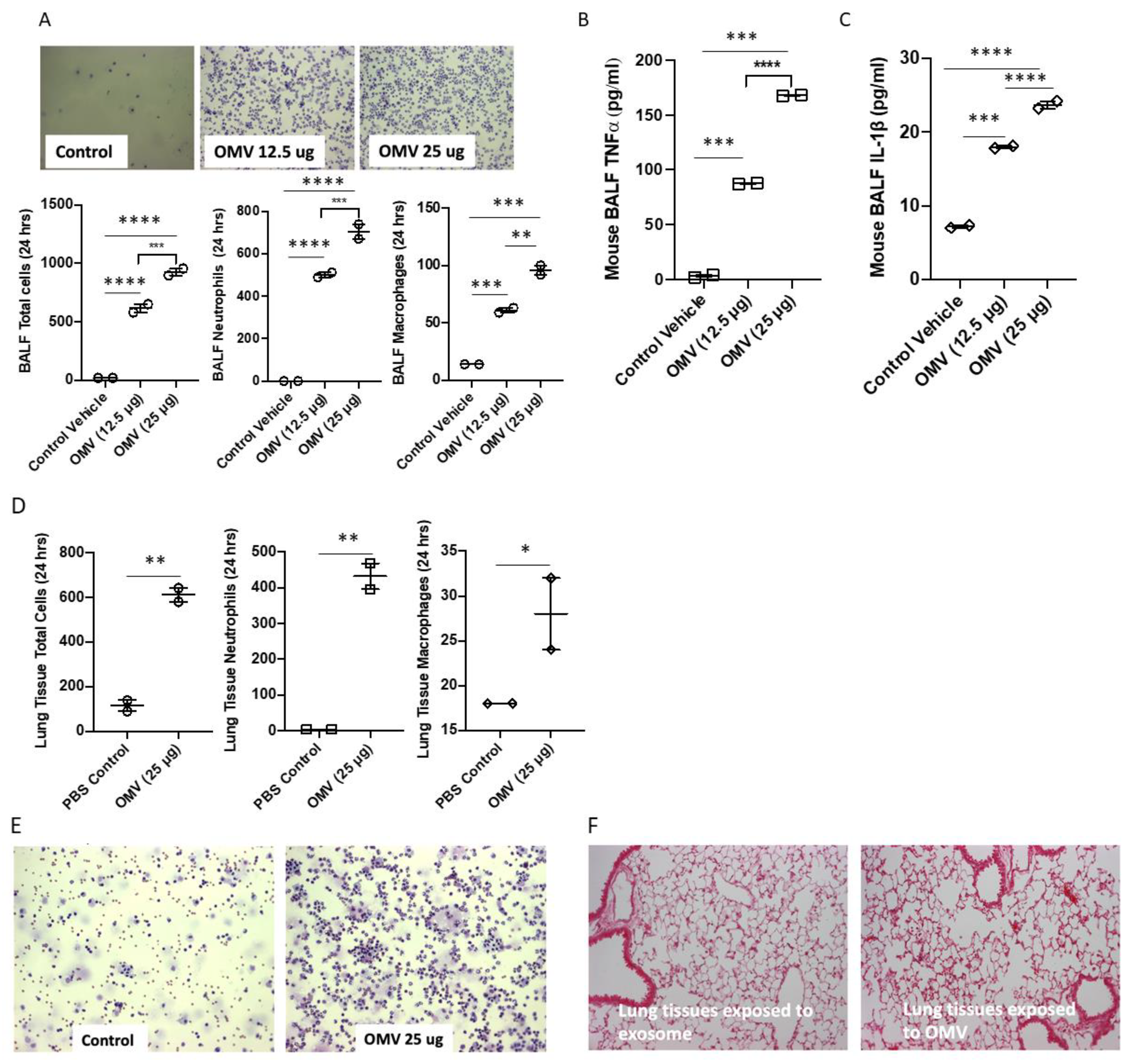
3.4. Gram-Negative Bacterial OMVs Induce Inflammatory Lung Responses In Vivo
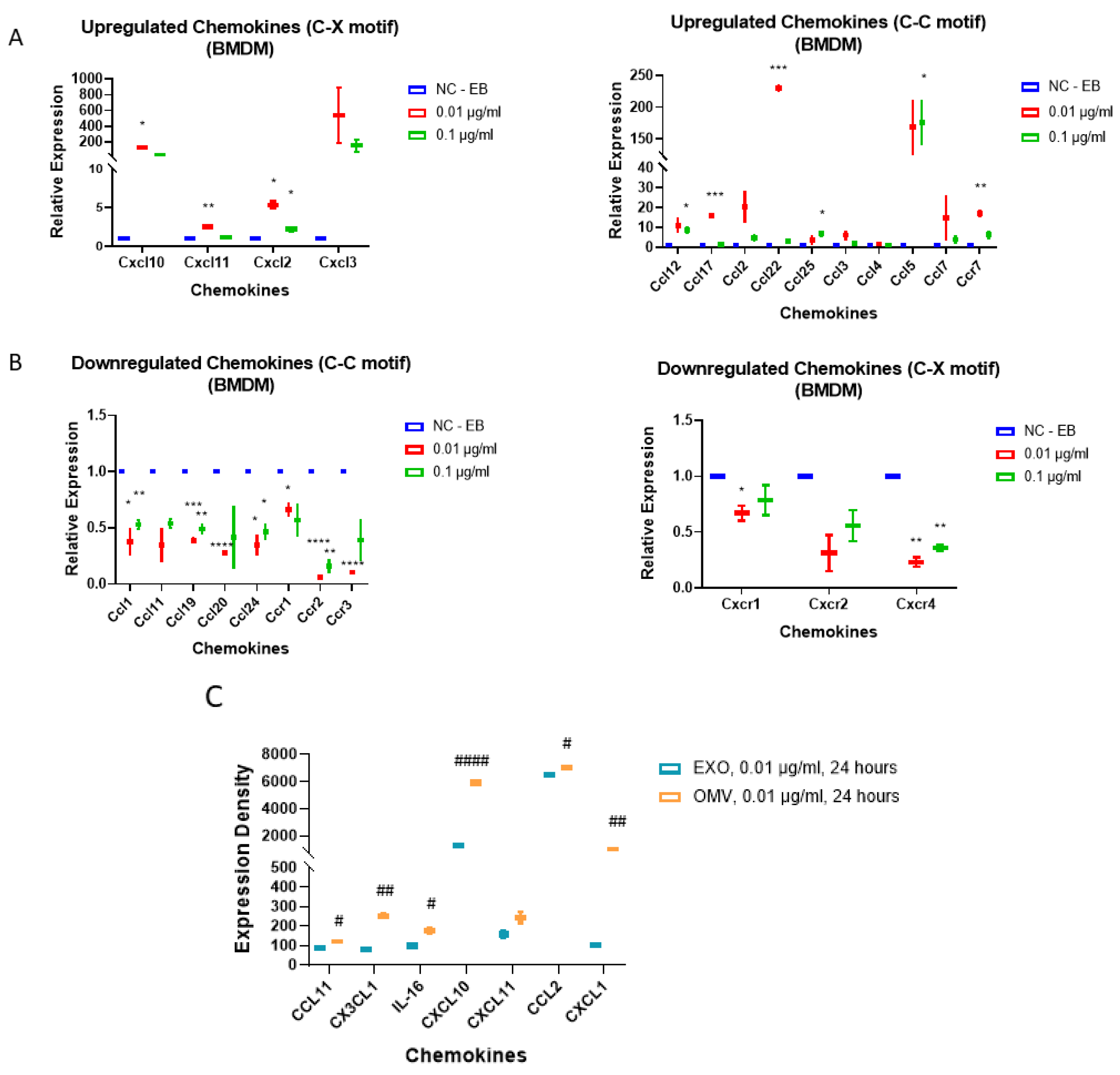
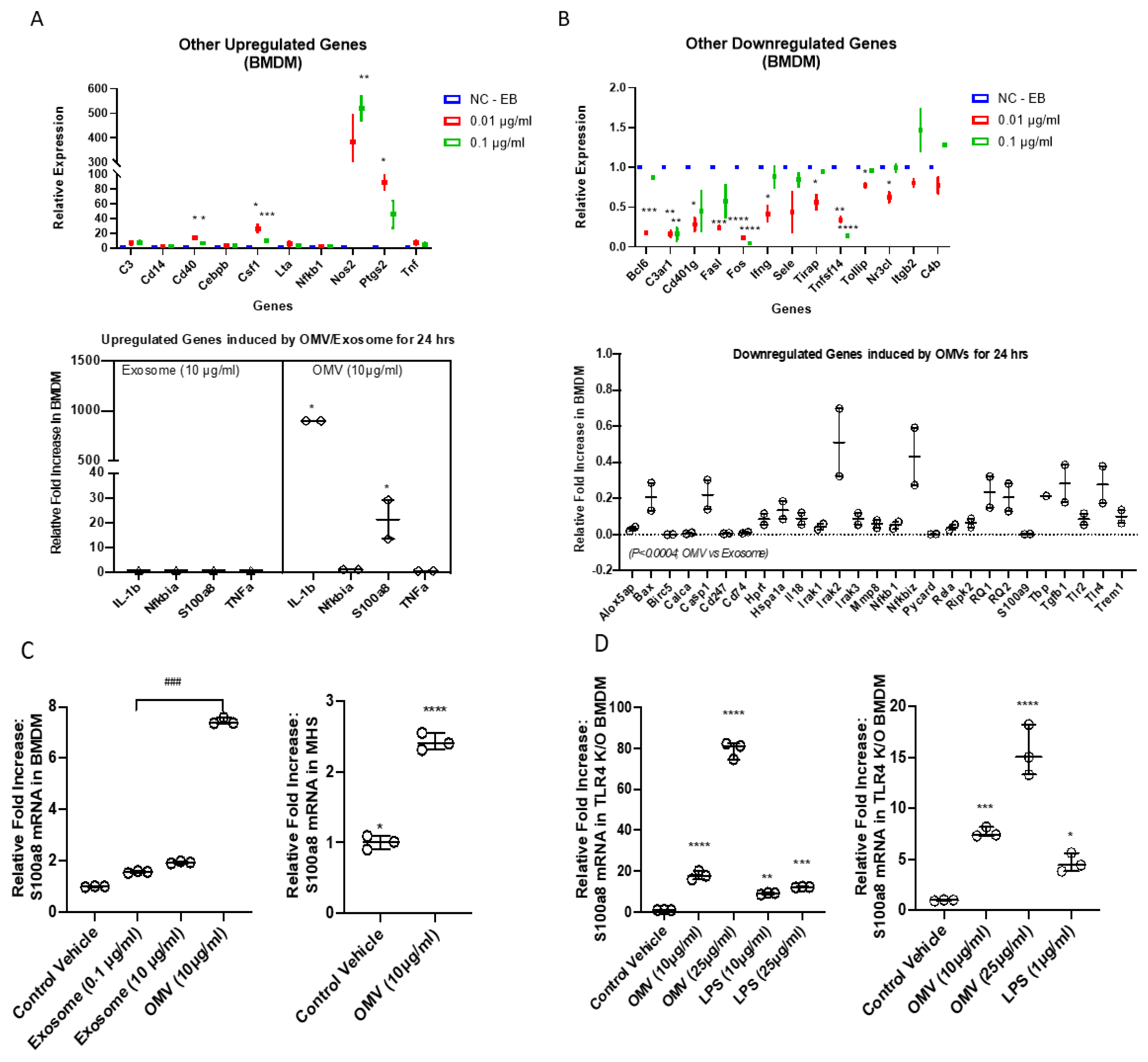
3.5. Gram-Negative Bacterial OMVs Induce Multiple Pro-Inflammatory Cytokines and Chemokines in Macrophages In Vitro
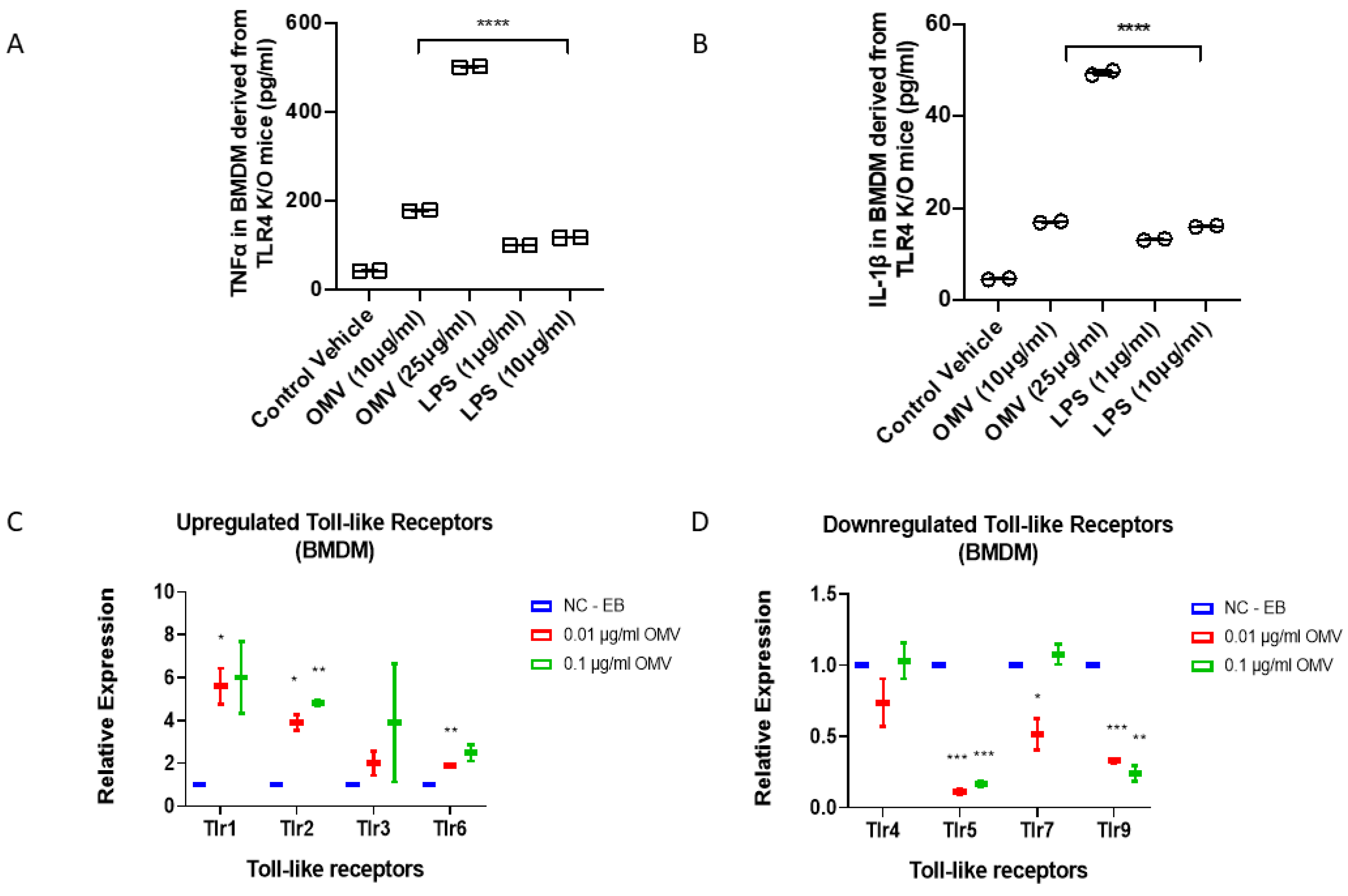
3.6. In Addition to TLR4, Gram-Negative Bacterial OMVs Induce Macrophage Activation via Multiple Signaling Pathways
3.7. Gram-Negative Bacterial OMVs Induce Pro-Inflammatory DAMPs Expression in Macrophages
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peleg, A.Y.; Hooper, D.C. Hospital-Acquired Infections Due to Gram-Negative Bacteria. N. Engl. J. Med. 2010, 362, 1804–1813. [Google Scholar] [CrossRef] [PubMed]
- Falguera, M.; Trujillano, J.; Caro, S.; Menéndez, R.; Carratala, J.; Ruiz-González, A.; Vilà, M.; García, M.; Porcel, J.M.; Torres, A.; et al. A Prediction Rule for Estimating the Risk of Bacteremia in Patients with Community-Acquired Pneumonia. Clin. Infect. Dis. 2009, 49, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Menéndez, R.; Torres, A.; Reyes, S.; Zalacain, R.; Capelastegui, A.; Aspa, J.; Borderias, L.; Martin-Villasclaras, J.J.; Bello, S.; Alfageme, I.; et al. Initial management of pneumonia and sepsis: Factors associated with improved outcome. Eur. Respir. J. 2012, 39, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-Y. Pneumonia, Acute Respiratory Distress Syndrome, and Early Immune-Modulator Therapy. Int. J. Mol. Sci. 2017, 18, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herold, S.; Mayer, K.; Lohmeyer, J. Acute Lung Injury: How Macrophages Orchestrate Resolution of Inflammation and Tissue Repair. Front. Immunol. 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmiel, J.F.; Konstan, M.W.; Elborn, J.S. Antibiotic and Anti-Inflammatory Therapies for Cystic Fibrosis. Cold Spring Harb. Perspect. Med. 2013, 3, a009779. [Google Scholar] [CrossRef] [Green Version]
- Benoit, M.; Desnues, B.; Mege, J.-L. Macrophage Polarization in Bacterial Infections. J. Immunol. 2008, 181, 3733–3739. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-J. Microbe-Host Communication by Small RNAs in Extracellular Vesicles: Vehicles for Transkingdom RNA Transportation. Int. J. Mol. Sci. 2019, 20, 1487. [Google Scholar] [CrossRef] [Green Version]
- Margolis, L.; Sadovsky, Y. The biology of extracellular vesicles: The known unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.N.; Kuehn, M.J. Virulence and Immunomodulatory Roles of Bacterial Outer Membrane Vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in Innate Immunity and Inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakihana, Y.; Ito, T.; Nakahara, M.; Yamaguchi, K.; Yasuda, T. Sepsis-induced myocardial dysfunction: Pathophysiology and management. J. Intensiv. Care 2016, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-Like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Marion, C.R.; Lee, J.; Sharma, L.; Park, K.-S.; Lee, C.; Liu, W.; Liu, P.; Feng, J.; Gho, Y.S.; Dela Cruz, C.S. Toll-like Receptors 2 and 4 Modulate Pulmonary Inflammation and Host Factors Mediated by Outer Membrane Vesicles Derived from Acinetobacter baumannii. Infect. Immun. 2019, 87, e00243-19. [Google Scholar] [CrossRef] [Green Version]
- Deo, P.; Chow, S.H.; Han, M.-L.; Speir, M.; Huang, C.; Schittenhelm, R.B.; Dhital, S.; Emery, J.; Li, J.; Kile, B.T.; et al. Mitochondrial dysfunction caused by outer membrane vesicles from Gram-negative bacteria activates intrinsic apoptosis and inflammation. Nat. Microbiol. 2020, 5, 1418–1427. [Google Scholar] [CrossRef]
- Cecil, J.D.; O’Brien-Simpson, N.M.; Lenzo, J.C.; Holden, J.A.; Singleton, W.; Perez-Gonzalez, A.; Mansell, A.; Reynolds, E.C. Outer Membrane Vesicles Prime and Activate Macrophage Inflammasomes and Cytokine Secretion In Vitro and In Vivo. Front. Immunol. 2017, 8, 1017. [Google Scholar] [CrossRef] [Green Version]
- Deo, P.; Chow, S.H.; Hay, I.D.; Kleifeld, O.; Costin, A.; Elgass, K.D.; Jiang, J.-H.; Ramm, G.; Gabriel, K.; Dougan, G.; et al. Outer membrane vesicles from Neisseria gonorrhoeae target PorB to mitochondria and induce apoptosis. PLoS Pathog. 2018, 14, e1006945. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.L.; Stoiber, C.; Herkt, C.E.; Schulz, C.; Bertrams, W.; Schmeck, B. Legionella pneumophila-Derived Outer Membrane Vesicles Promote Bacterial Replication in Macrophages. PLoS Pathog. 2016, 12, e1005592. [Google Scholar] [CrossRef]
- Moon, H.-G.; Yang, J.; Zheng, Y.; Jin, Y. miR-15a/16 Regulates Macrophage Phagocytosis after Bacterial Infection. J. Immunol. 2014, 193, 4558–4567. [Google Scholar] [CrossRef] [Green Version]
- Carnino, J.M.; Ryu, S.; Ni, K.; Jin, Y. Pretreated household materials carry similar filtration protection against pathogens when compared with surgical masks. Am. J. Infect. Control. 2020, 48, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lee, H.; Zhu, Z.; Minhas, J.K.; Jin, Y. Enrichment of selective miRNAs in exosomes and delivery of exosomal miRNAs in vitro and in vivo. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L110–L121. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Li, C.; Zhang, Y.; Zhang, D.; Otterbein, L.E.; Jin, Y. Caveolin-1 selectively regulates microRNA sorting into microvesicles after noxious stimuli. J. Exp. Med. 2019, 216, 2202–2220. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Zhang, D.; Wu, J.; Otterbein, L.E.; Jin, Y. Lung Epithelial Cell–Derived Microvesicles Regulate Macrophage Migration via MicroRNA-17/221–Induced Integrin β1 Recycling. J. Immunol. 2017, 199, 1453–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, F.S.Y.; De Almeida, S.R. The Role of Phagocytes and NETs in Dermatophytosis. Mycopathologia 2017, 182, 263–272. [Google Scholar] [CrossRef]
- Mlcochova, P.; Winstone, H.; Zuliani-Alvarez, L.; Gupta, R.K. TLR4-Mediated Pathway Triggers Interferon-Independent G0 Arrest and Antiviral SAMHD1 Activity in Macrophages. Cell Rep. 2020, 30, 3972–3980.e5. [Google Scholar] [CrossRef] [PubMed]
- Srikrishna, G. S100A8 and S100A9: New Insights into Their Roles in Malignancy. J. Innate Immun. 2012, 4, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Avila-Calderon, E.D.; Araiza-Villanueva, M.G.; Cancino-Diaz, J.C.; Lopez-Villegas, E.O.; Sriranganathan, N.; Boyle, S.M.; Contreras-Rodriguez, A. Roles of bacterial membrane vesicles. Arch. Microbiol. 2015, 197, 1–10. [Google Scholar] [CrossRef]
- Doerrler, W.T. Lipid trafficking to the outer membrane of Gram-negative bacteria. Mol. Microbiol. 2006, 60, 542–552. [Google Scholar] [CrossRef]
- Chatterjee, S.N.; Das, J. Electron Microscopic Observations on the Excretion of Cell-wall Material by Vibrio cholerae. J. Gen. Microbiol. 1967, 49, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.-S.; Lee, J.; Jang, S.C.; Kim, S.R.; Jang, M.H.; Lötvall, J.; Kim, Y.-K.; Gho, Y.S. Pulmonary Inflammation Induced by Bacteria-Free Outer Membrane Vesicles from Pseudomonas aeruginosa. Am. J. Respir. Cell Mol. Biol. 2013, 49, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yoon, Y.J.; Kim, J.H.; Dinh, N.T.H.; Go, G.; Tae, S.; Park, K.-S.; Park, H.T.; Lee, C.; Roh, T.-Y.; et al. Outer Membrane Vesicles Derived From Escherichia coli Regulate Neutrophil Migration by Induction of Endothelial IL-8. Front. Microbiol. 2018, 9, 2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfalzgraff, A.; Correa, W.; Heinbockel, L.; Schromm, A.B.; Lübow, C.; Gisch, N.; Martinez-De-Tejada, G.; Brandenburg, K.; Weindl, G. LPS-neutralizing peptides reduce outer membrane vesicle-induced inflammatory responses. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 1503–1513. [Google Scholar] [CrossRef]
- Manning, A.J.; Kuehn, M.J. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Badi, S.A.; Bruno, S.P.; Moshiri, A.; Tarashi, S.; Siadat, S.D.; Masotti, A. Small RNAs in Outer Membrane Vesicles and Their Function in Host-Microbe Interactions. Front. Microbiol. 2020, 11, 1209. [Google Scholar] [CrossRef]
- Love, A.C.; Schwartz, I.; Petzke, M.M. Borrelia burgdorferi RNA Induces Type I and III Interferons via Toll-Like Receptor 7 and Contributes to Production of NF-κB-Dependent Cytokines. Infect. Immun. 2014, 82, 2405–2416. [Google Scholar] [CrossRef] [Green Version]
- Petzke, M.M.; Iyer, R.; Love, A.C.; Spieler, Z.; Brooks, A.; Schwartz, I. Borrelia burgdorferi induces a type I interferon response during early stages of disseminated infection in mice. BMC Microbiol. 2016, 16, 29. [Google Scholar] [CrossRef] [Green Version]
- Kaparakis, M.; Turnbull, L.; Carneiro, L.; Firth, S.; Coleman, H.A.; Parkington, H.C.; Le Bourhis, L.; Karrar, A.; Viala, J.; Mak, J.; et al. Bacterial membrane vesicles deliver peptidoglycan to NOD1 in epithelial cells. Cell. Microbiol. 2010, 12, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Patten, D.A.; Hussein, E.; Davies, S.P.; Humphreys, P.N.; Collett, A. Commensal-derived OMVs elicit a mild proinflammatory response in intestinal epithelial cells. Microbiology 2017, 163, 702–711. [Google Scholar] [CrossRef]
- Yu, Y.-J.; Wang, X.-H.; Fan, G.-C. Versatile effects of bacterium-released membrane vesicles on mammalian cells and infectious/inflammatory diseases. Acta Pharmacol. Sin. 2018, 39, 514–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathirana, R.D.; Kaparakis-Liaskos, M. Bacterial membrane vesicles: Biogenesis, immune regulation and pathogenesis. Cell. Microbiol. 2016, 18, 1518–1524. [Google Scholar] [CrossRef] [Green Version]
- Moon, D.C.; Choi, C.H.; Lee, J.H.; Choi, C.-W.; Kim, H.-Y.; Park, J.S.; Kim, S.I.; Lee, J.C. Acinetobacter baumannii outer membrane protein a modulates the biogenesis of outer membrane vesicles. J. Microbiol. 2012, 50, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Rodriguez, D.L.; Kuehn, M.J. NlpI-mediated modulation of outer membrane vesicle production through peptidoglycan dynamics in Escherichia coli. Microbiologyopen 2015, 4, 375–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, L.; Praszkier, J.; Hutton, M.L.; Steer, D.; Ramm, G.; Kaparakis-Liaskos, M.; Ferrero, R.L. Increased Outer Membrane Vesicle Formation in aHelicobacter pylori tolBMutant. Helicobacter 2015, 20, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Sinha, R.; Mitobe, J.; Koley, H. Development of a cost-effective vaccine candidate with outer membrane vesicles of a tolA-disrupted Shigella boydii strain. Vaccine 2016, 34, 1839–1846. [Google Scholar] [CrossRef]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 2005, 437, 422–425. [Google Scholar] [CrossRef]
- McMahon, K.J.; Castelli, M.E.; Véscovi, E.G.; Feldman, M.F. Biogenesis of Outer Membrane Vesicles in Serratia marcescens Is Thermoregulated and Can Be Induced by Activation of the Rcs Phosphorelay System. J. Bacteriol. 2012, 194, 3241–3249. [Google Scholar] [CrossRef] [Green Version]
- McBroom, A.J.; Kuehn, M.J. Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response. Mol. Microbiol. 2007, 63, 545–558. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Beveridge, T.J. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: A novel mechanism of enzyme secretion. J. Bacteriol. 1995, 177, 3998–4008. [Google Scholar] [CrossRef]
- Sartz, L.; De Jong, B.; Hjertqvist, M.; Plym-Forshell, L.; Alsterlund, R.; Löfdahl, S.; Osterman, B.; Ståhl, A.; Eriksson, E.; Hansson, H.-B.; et al. An outbreak of Escherichia coli O157:H7 infection in southern Sweden associated with consumption of fermented sausage; aspects of sausage production that increase the risk of contamination. Epidemiol. Infect. 2008, 136, 370–380. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′–3′) |
|---|---|
| TNFα-F | 5′-GACGTGGAACTGGCAGAAGAG-3′ |
| TNFα-R | 5′-TTGGTGGTTTGTGAGTGTGAG-3′ |
| IL1β-F | 5′-GCAACTGTTCCTGAACTCAACT-3′ |
| IL1β-R | 5′-ATCTTTTGGGGTCCGTCAACT-3′ |
| iNOS-F | 5′-CAAGCTGAACTTGAGCGAGGA-3′ |
| iNOS-R | 5′-TTTACTCAGTGCCAGAAGCTGGA-3′ |
| CXCL1-F | 5′-CTGGGATTCACCTCAAGAACATC-3′ |
| CXCL1-R | 5′-CAGGGTCAAGGCAAGCCTC-3′ |
| IL10-F | 5′-GCTCTTACTGACTGGCATGAG-3′ |
| IL10-R | 5′_CGCAGCTCTAGGAGCATGTG-3′ |
| TBP-F | 5′-TCAAACCCAGAATTGTTCTCC-3′ |
| TBP-R | 5′-GGGGTAGATGTTTTCAAATGC-3′ |
| S100a8-F | 5′-CAAGGAAATCACCATGCCCTCTA-3′ |
| S100a8-R | 5′-ACCATCGCAAGGAACTCCTCGA-3′ |
| Upregulated Chemokine | Fold |
|---|---|
| CXCL1 | 8.8238 |
| CXCL2 | 5.5132 |
| CXCL9 | 4.6757 |
| CXCL10 | 4.6167 |
| CX3CL1 | 3.0991 |
| CCL27 | 2.8644 |
| CCL6 | 2.8452 |
| CCL5 | 2.6587 |
| Chemerin | 2.6376 |
| CXCL16 | 2.4300 |
| CCL28 | 2.3155 |
| CCL21 | 2.3072 |
| CXCL13 | 2.2632 |
| Downregulated Chemokine | Fold |
| C5/C5a | 0.35372 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, S.; Ni, K.; Wang, C.; Sivanantham, A.; Carnino, J.M.; Ji, H.-L.; Jin, Y. Bacterial Outer Membrane Vesicles Promote Lung Inflammatory Responses and Macrophage Activation via Multi-Signaling Pathways. Biomedicines 2023, 11, 568. https://doi.org/10.3390/biomedicines11020568
Ryu S, Ni K, Wang C, Sivanantham A, Carnino JM, Ji H-L, Jin Y. Bacterial Outer Membrane Vesicles Promote Lung Inflammatory Responses and Macrophage Activation via Multi-Signaling Pathways. Biomedicines. 2023; 11(2):568. https://doi.org/10.3390/biomedicines11020568
Chicago/Turabian StyleRyu, Sunhyo, Kareemah Ni, Chenghao Wang, Ayyanar Sivanantham, Jonathan M. Carnino, Hong-Long Ji, and Yang Jin. 2023. "Bacterial Outer Membrane Vesicles Promote Lung Inflammatory Responses and Macrophage Activation via Multi-Signaling Pathways" Biomedicines 11, no. 2: 568. https://doi.org/10.3390/biomedicines11020568