
Screening MT1-MMP Activity and Inhibition in Three-Dimensional Tumor Spheroids
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
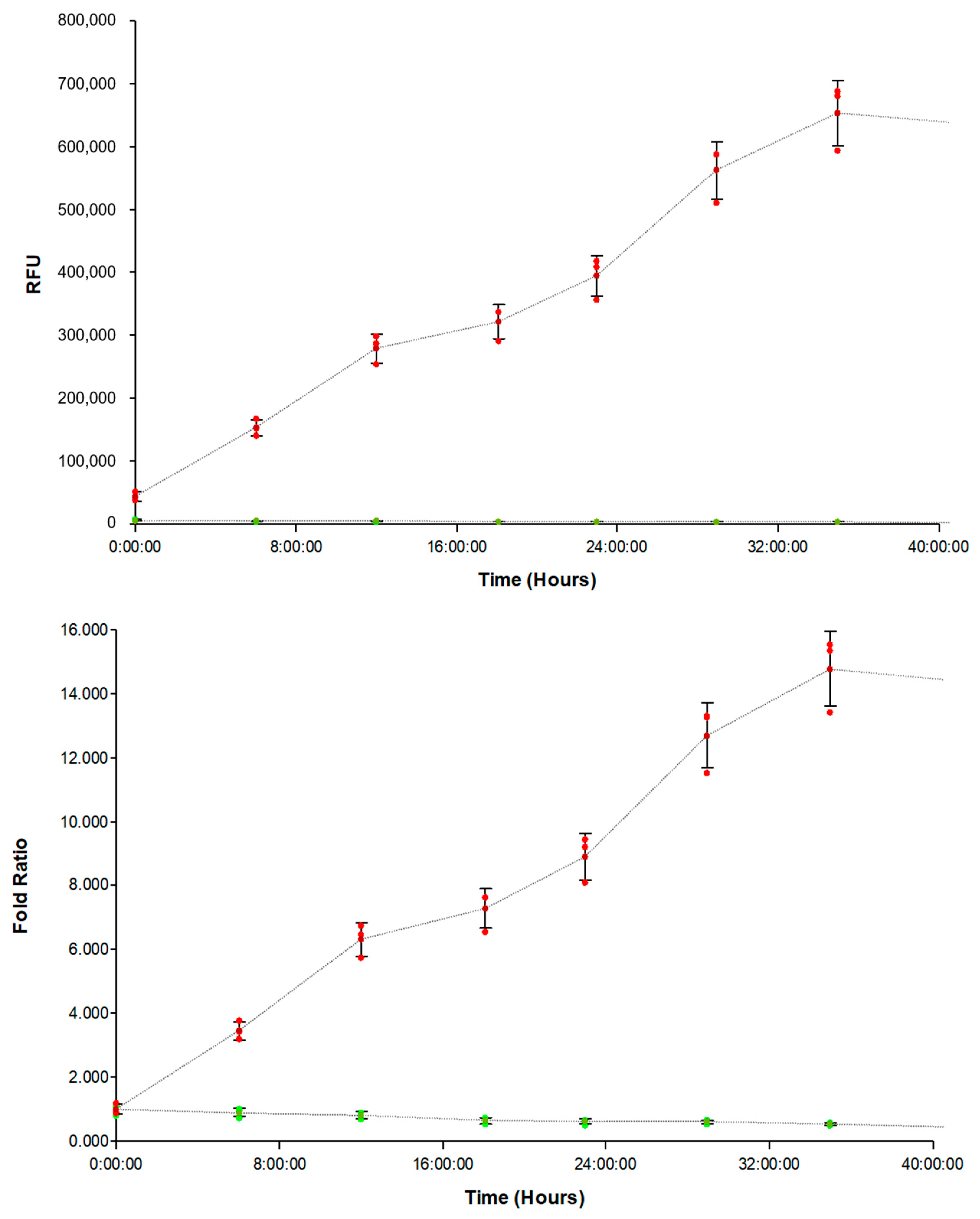
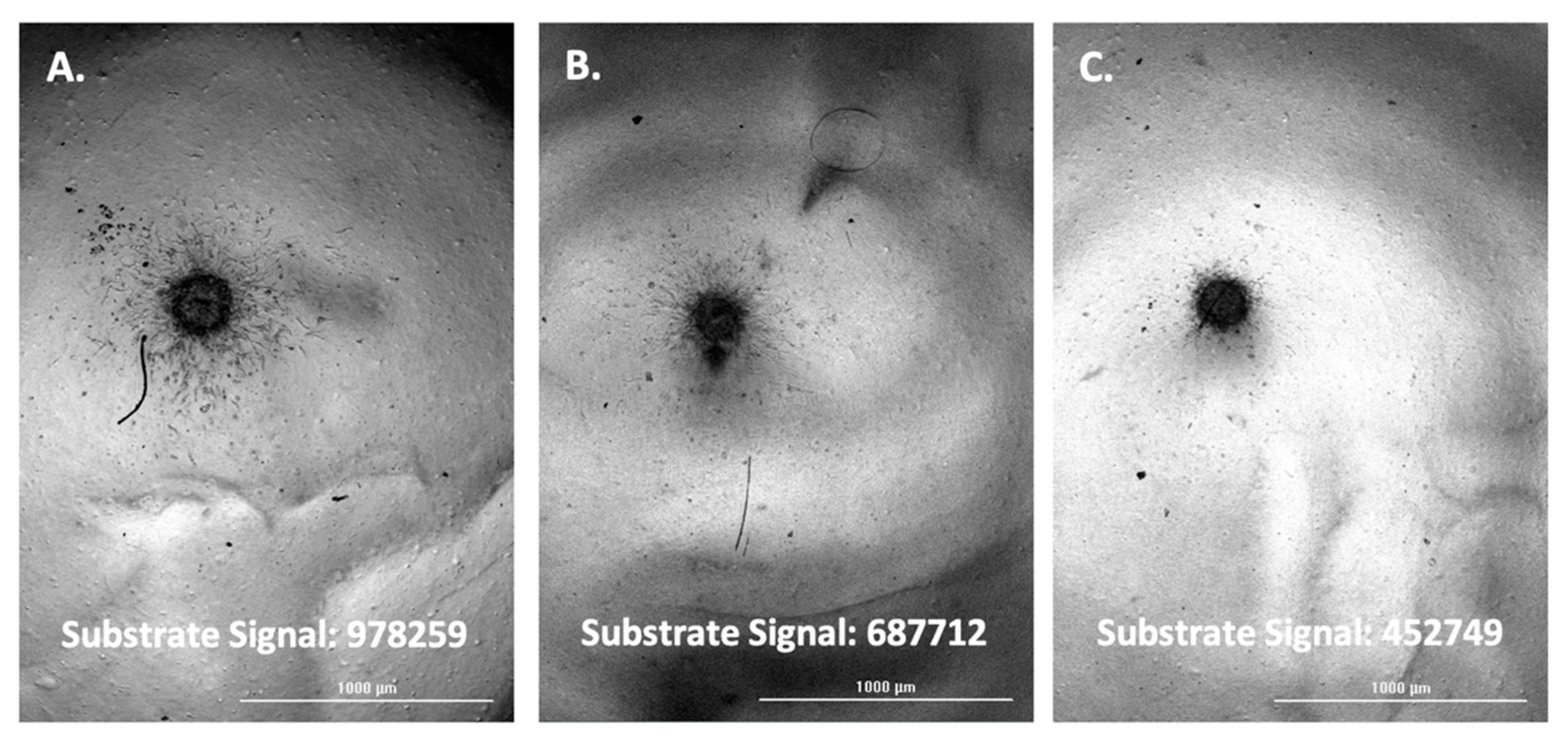
3.1. Evaluation of FAM-fTHP-9 Signal Detection
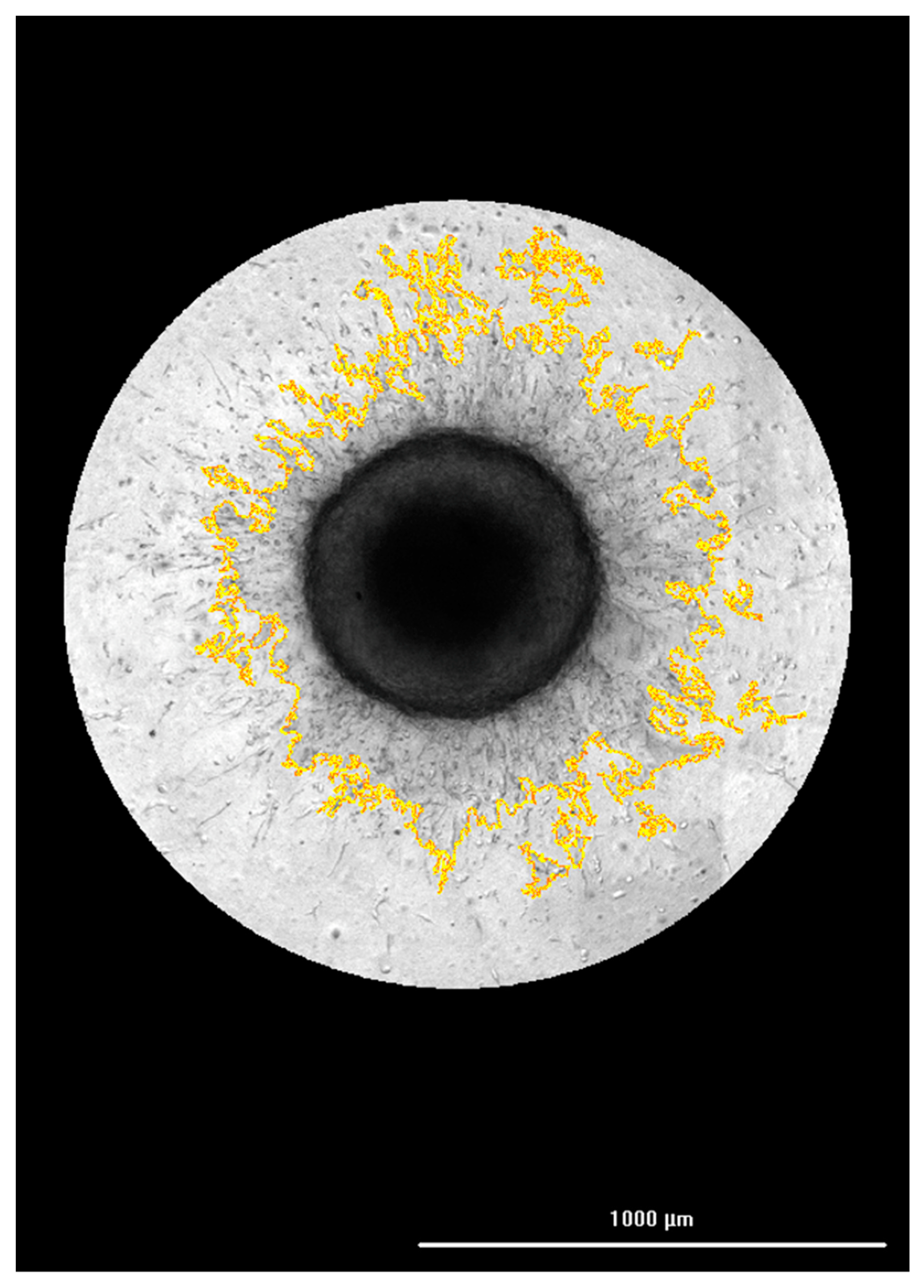
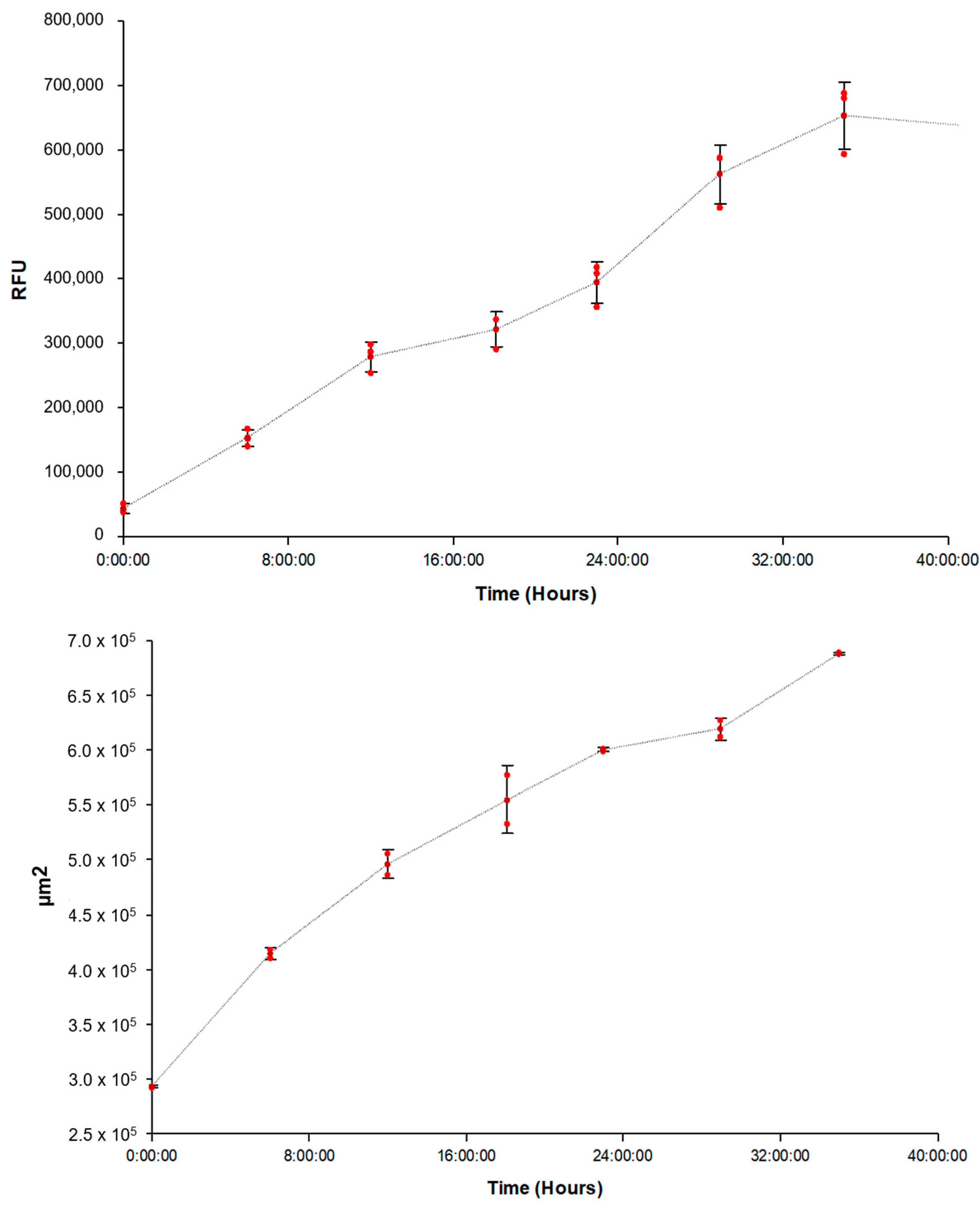
3.2. Correlation of FAM-fTHP-9 Signal with Increased Spheroid Invasion
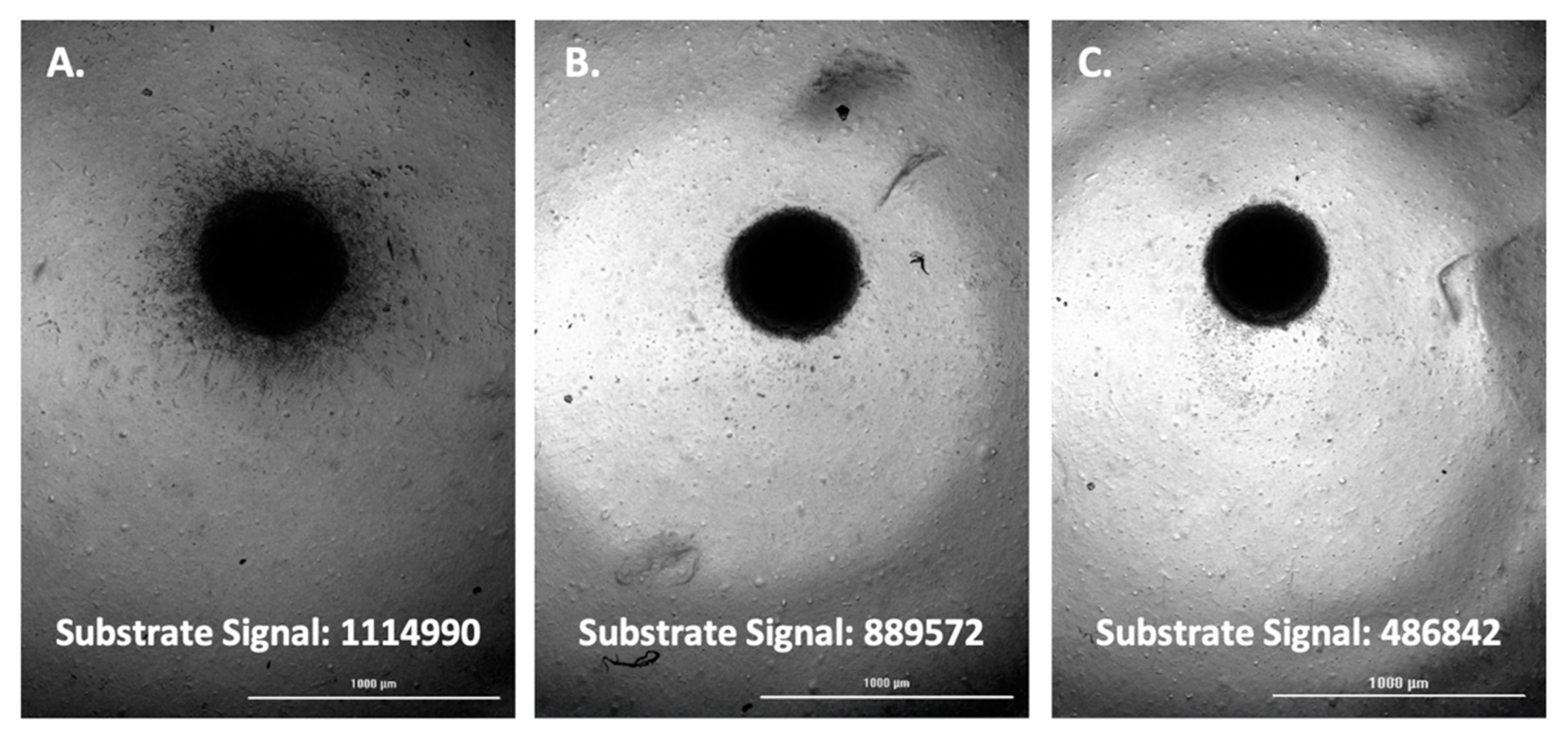
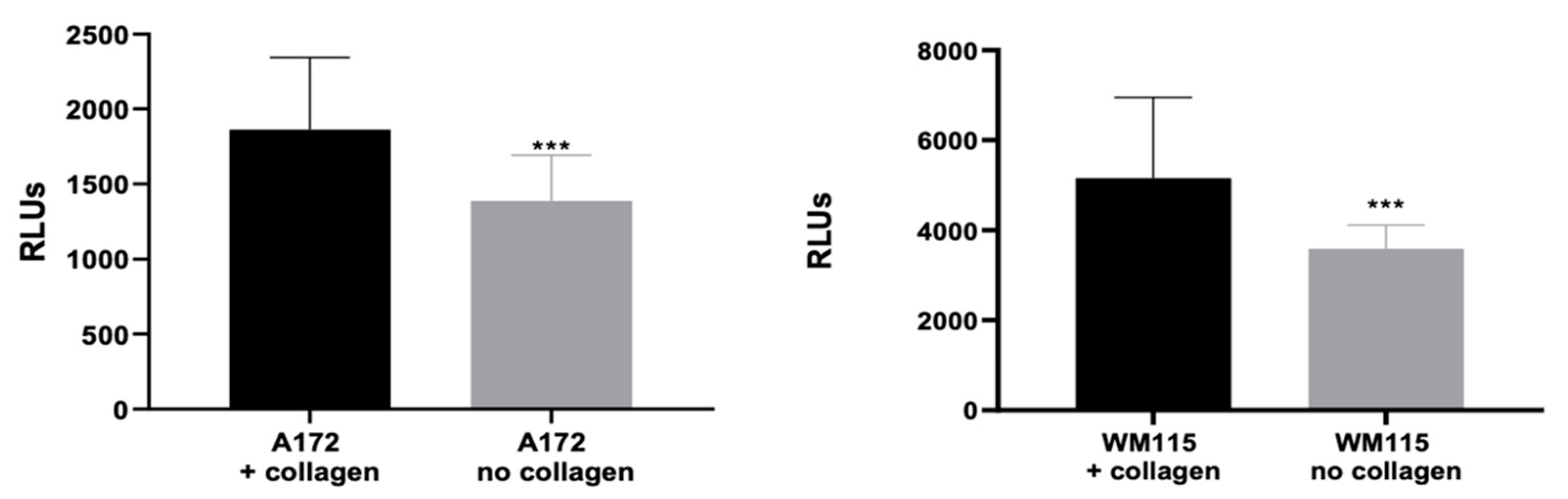
3.3. Evaluating the Origin of FAM-fTHP-9 Cleavage
3.4. Hit Pick Assay Performance Using FAM-fTHP-9 Signal to Trigger Imaging of Test Inhibitor Wells
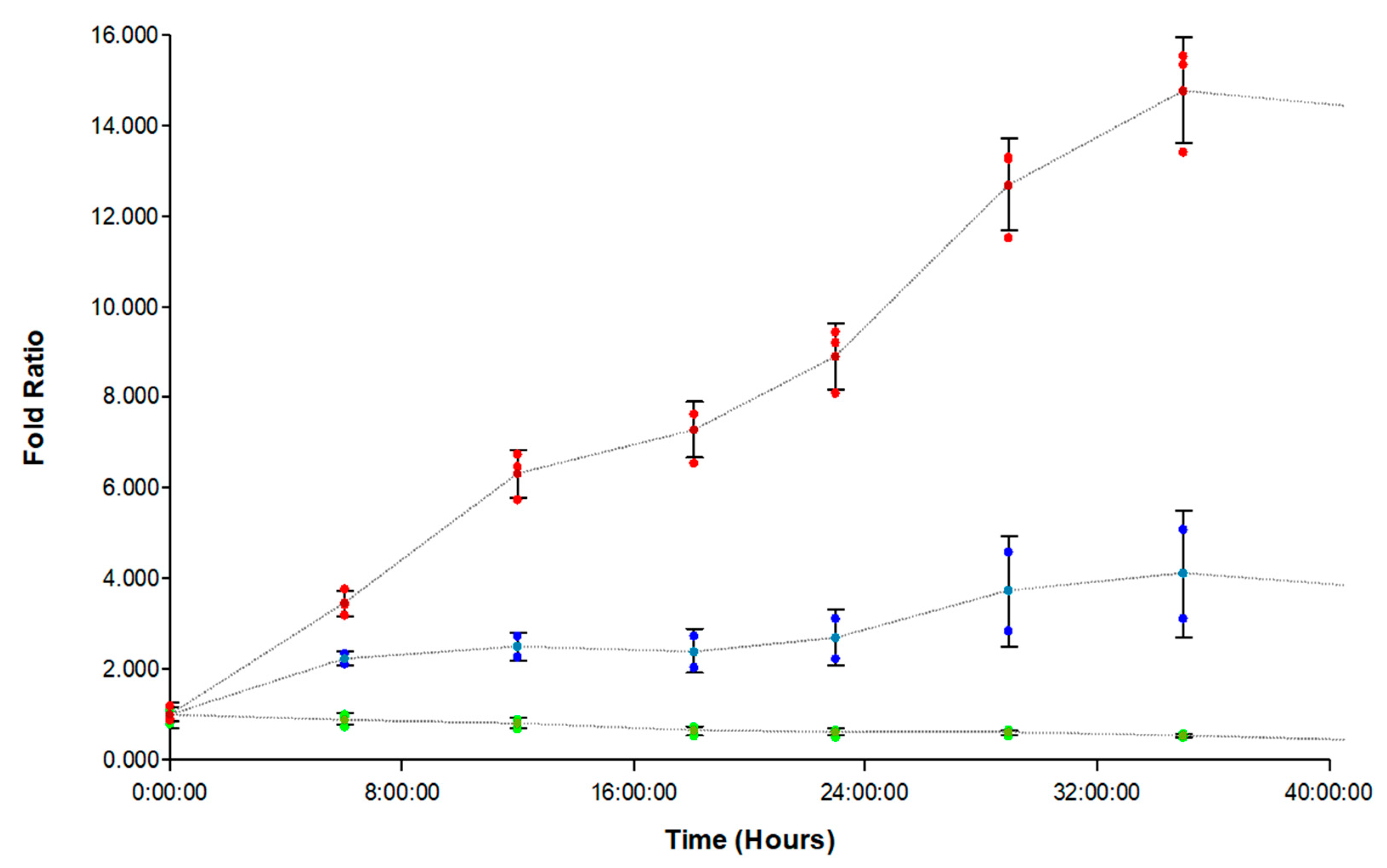
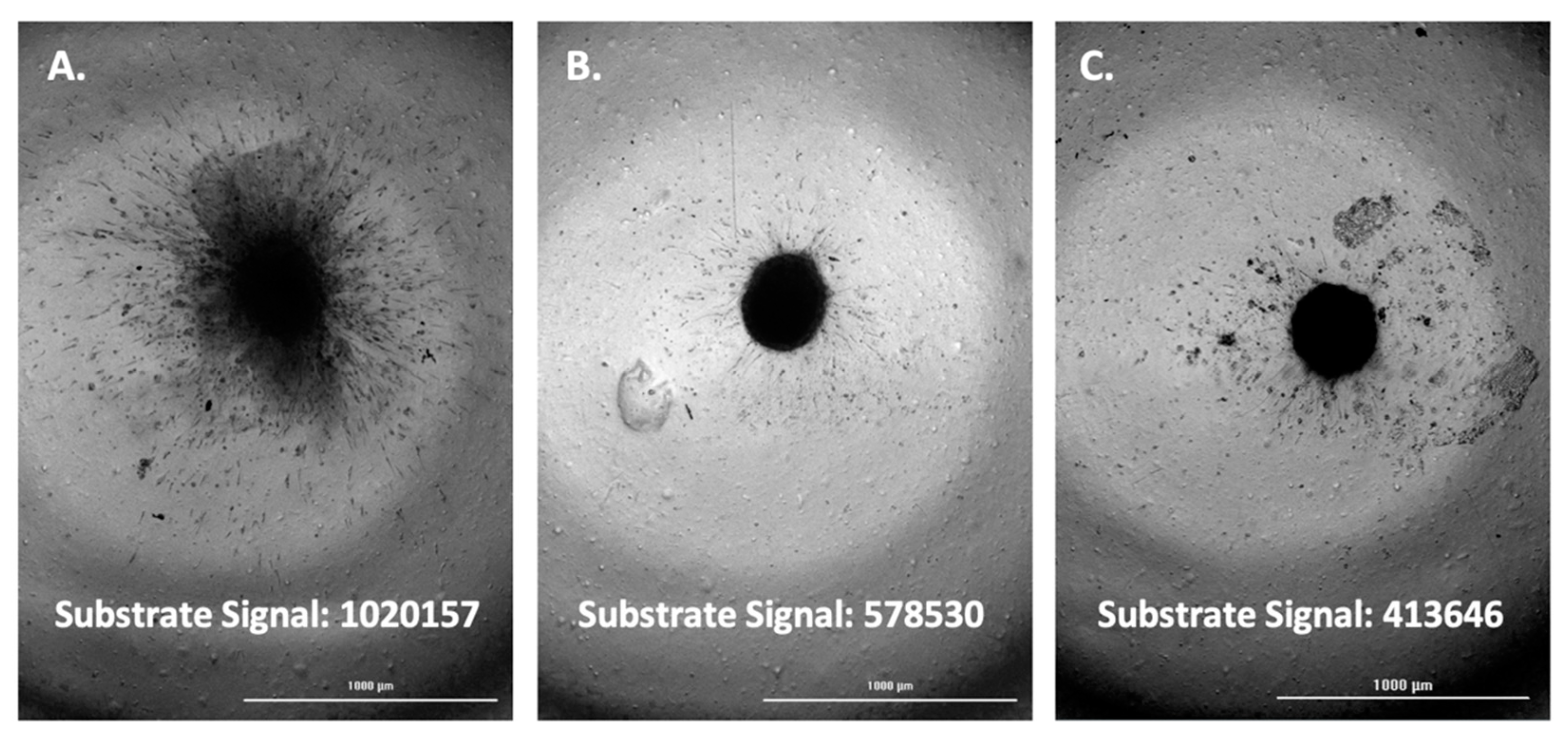
3.5. Evaluation of FAM-fTHP-9 Signal Detection and Spheroid Invasion in A172, H4, and SW 1088 Cell Lines
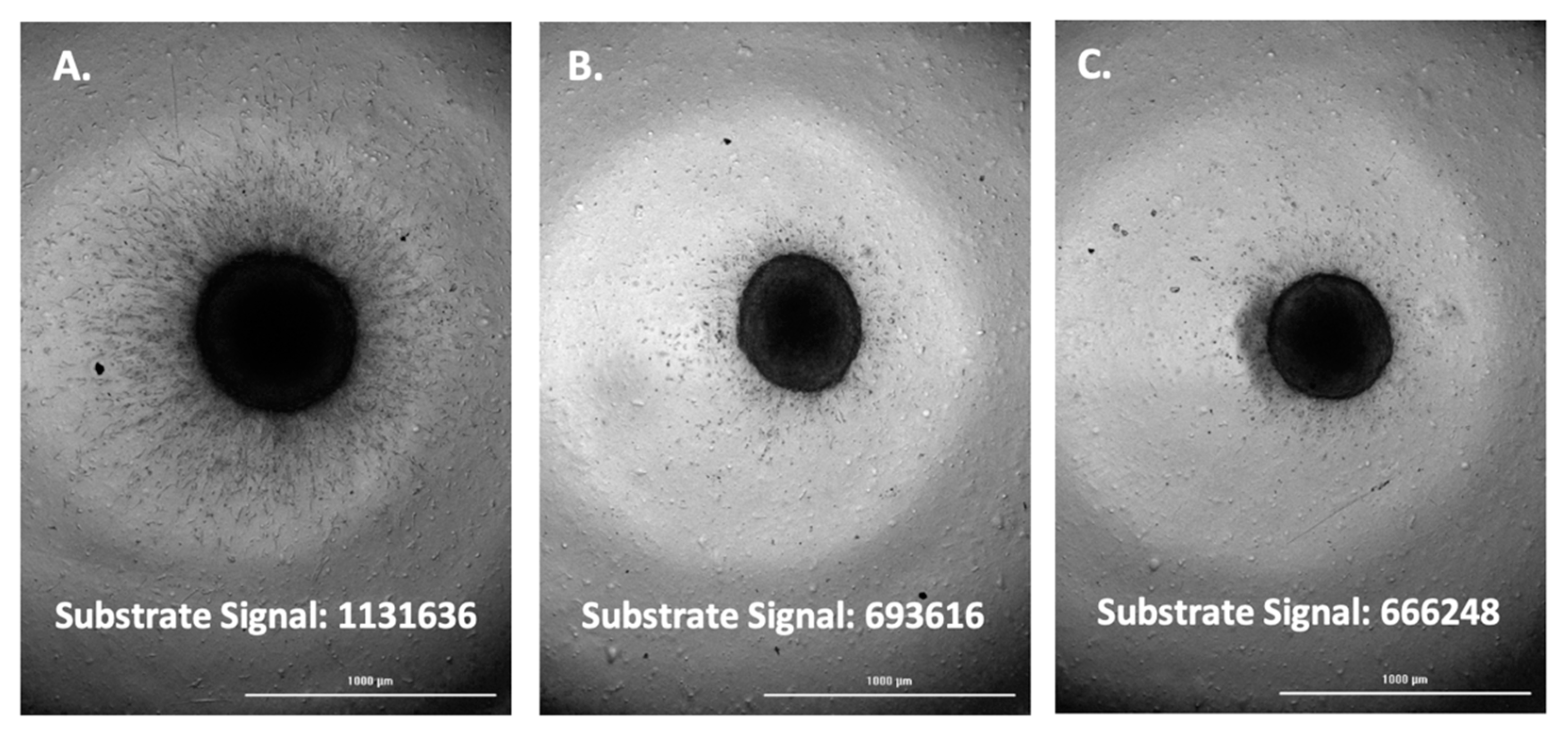
3.6. Inhibition of FAM-fTHP-9 Signal and Spheroid Invasion in A172, H4, and SW 1088 Cell Lines
3.7. Western Blot Analysis
3.8. Cell Viability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Association of Neurological Surgeons. Brain Tumors—Classifications, Symptoms, Diagnosis and Treatments. Available online: https://www.aans.org/en/Patients/Neurosurgical-Conditions-and-Treatments/Brain-Tumors (accessed on 24 October 2021).
- Ostrom, Q.; Gittleman, H.; Farah, P.; Ondracek, A.; Chen, Y.; Wolinsky, Y.; Stroup, N.E.; Kruchko, C.; Barnholtz-Sloan, J. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2006–2010. Neuro Oncol. 2013, 15, ii1–ii56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Klockow, J.L.; Zhang, M.; Lafortune, F.; Chang, E.; Jin, L.; Wu, Y.; Daldrup-Link, H.E. Glioblastoma multiforme (GBM): An overview of current therapies and mechanisms of resistance. Pharmacol. Res. 2021, 171, 105780. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Sounni, N.E.; Devy, L.; Hajitou, A.; Frankenne, F.; Munaut, C.; Gilles, C.; Deroanne, C.; Thompson, E.W.; Foidart, J.M.; Noel, A. MT1-MMP expression promotes tumor growth and angiogenesis through an up-regulation of vascular endothelial growth factor expression. FASEB J. 2002, 16, 555–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiki, M. Membrane-type 1 matrix metalloproteinase: A key enzyme for tumor invasion. Cancer Lett. 2003, 194, 1–11. [Google Scholar] [CrossRef]
- Arroyo, A.; Genis, L.; Gonzalo, P.; Matias-Roman, S.; Pollan, A.; Galvez, B.G. Matrix Metalloproteinases: New Routes to the Use of MT1-MMP As a Therapeutic Target in Angiogenesis-Related Disease. Curr. Pharm. Des. 2007, 13, 1787–1802. [Google Scholar] [CrossRef]
- Itoh, Y.; Ito, N.; Nagase, H.; Evans, R.D.; Bird, S.A.; Seiki, M. Cell Surface Collagenolysis Requires Homodimerization of the Membrane-bound Collagenase MT1-MMP. Mol. Biol. Cell 2006, 17, 5390–5399. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Seiki, M. MT1-MMP: A potent modifier of pericellular microenvironment. J. Cell. Physiol. 2006, 206, 1–8. [Google Scholar] [CrossRef]
- Markovic, D.S.; Vinnakota, K.; Chirasani, S.; Synowitz, M.; Raguet, H.; Stock, K.; Sliwa, M.; Lehmann, S.; Kälin, R.; van Rooijen, N.; et al. Gliomas induce and exploit microglial MT1-MMP expression for tumor expansion. Proc. Natl. Acad. Sci. USA 2009, 106, 12530–12535. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Xue, Y.-X.; Liu, L.-B.; Wang, P.; Liu, Y.-H.; Ying, H.-Q. Expressions of matrix metalloproteinase-7 and matrix metalloproteinase-14 associated with the activation of extracellular signal-regulated kinase1/2 in human brain gliomas of different pathological grades. Med. Oncol. 2011, 28, S433–S438. [Google Scholar] [CrossRef]
- Wang, L.; Yuan, J.; Tu, Y.; Mao, X.; He, S.; Fu, G.; Zong, J.; Zhang, Y. Co-expression of MMP-14 and MMP-19 predicts poor survival in human glioma. Clin. Transl. Oncol. 2013, 15, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, S.; Zhou, L.; Yang, L.; Wu, X.; Tang, B.; Xie, S.; Fang, L.; Zheng, S.; Hong, T. Immune Infiltration of MMP14 in Pan Cancer and Its Prognostic Effect on Tumors. Front. Oncol. 2021, 11, 717606. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Sang, W.; Su, L.; Shen, Y.; Hu, Y.; Zhang, W. Analysis of the expression and prognostic value of MT1-MMP, β1-integrin and YAP1 in glioma. Open Med. 2022, 17, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Takino, T.; Okada, Y.; Cao, J.; Shinagawa, A.; Yamamoto, E.; Seiki, M. A matrix metalloproteinase expressed on the surface of invasive tumour cells. Nature 1994, 370, 61–65. [Google Scholar] [CrossRef]
- VanMeter, T.E.; Rooprai, H.K.; Kibble, M.M.; Fillmore, H.L.; Broaddus, W.C.; Pilkington, G.J. The role of matrix metalloproteinase genes in glioma invasion: Co-dependent and interactive proteolysis. J. Neuro Oncol. 2001, 53, 213–235. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Hsieh, I.-Y.; Huang, X.; Li, J.; Zhao, W. Glioblastoma Stem-Like Cells: Characteristics, Microenvironment, and Therapy. Front. Pharmacol. 2016, 7, 477. [Google Scholar] [CrossRef] [Green Version]
- Tam, E.M.; Wu, Y.I.; Butler, G.S.; Stack, M.S.; Overall, C.M. Collagen Binding Properties of the Membrane Type-1 Matrix Metalloproteinase (MT1-MMP) Hemopexin C Domain. J. Biol. Chem. 2002, 277, 39005–39014. [Google Scholar] [CrossRef] [Green Version]
- Rape, A.; Ananthanarayanan, B.; Kumar, S. Engineering strategies to mimic the glioblastoma microenvironment. Adv. Drug Deliv. Rev. 2014, 79-80, 172–183. [Google Scholar] [CrossRef]
- Bolteus, A.J.; Berens, M.E.; Pilkington, G.J. Migration and invasion in brain neoplasms. Curr. Neurol. Neurosci. Rep. 2001, 1, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Griffith, L.G.; Swartz, M.A. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.M.; Cukierman, E. Modeling Tissue Morphogenesis and Cancer in 3D. Cell 2007, 130, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubert, C.G.; Rivera, M.; Spangler, L.C.; Wu, Q.; Mack, S.C.; Prager, B.C.; Couce, M.; McLendon, R.E.; Sloan, A.E.; Rich, J.N. A Three-Dimensional Organoid Culture System Derived from Human Glioblastomas Recapitulates the Hypoxic Gradients and Cancer Stem Cell Heterogeneity of Tumors Found In Vivo. Cancer Res. 2016, 76, 2465–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, W.; Zhu, H.; Zhao, M.; Zhou, Q.; Yin, W.; Liu, W.; Wang, L.; Xiao, Z.; Jiang, X.; Dai, J.; et al. Potential mechanisms underlying the promoting effects of 3D collagen scaffold culture on stemness and drug resistance of glioma cells. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166522. [Google Scholar] [CrossRef] [PubMed]
- Pedron, S.; Becka, E.; Harley, B.A. Regulation of glioma cell phenotype in 3D matrices by hyaluronic acid. Biomaterials 2013, 34, 7408–7417. [Google Scholar] [CrossRef]
- Pedron, S.; Becka, E.; Harley, B.A. Spatially Gradated Hydrogel Platform as a 3D Engineered Tumor Microenvironment. Adv. Mater. 2015, 27, 1567–1572. [Google Scholar] [CrossRef]
- Jiglaire, C.J.; Baeza-Kallee, N.; Denicolaï, E.; Barets, D.; Metellus, P.; Padovani, L.; Chinot, O.; Figarella-Branger, D.; Fernandez, C. Ex vivo cultures of glioblastoma in three-dimensional hydrogel maintain the original tumor growth behavior and are suitable for preclinical drug and radiation sensitivity screening. Exp. Cell Res. 2014, 321, 99–108. [Google Scholar] [CrossRef]
- Minond, D.; Lauer-Fields, J.L.; Nagase, H.; Fields, G.B. Matrix Metalloproteinase Triple-Helical Peptidase Activities Are Differentially Regulated by Substrate Stability. Biochemistry 2004, 43, 11474–11481. [Google Scholar] [CrossRef]
- Tokmina-Roszyk, M.; Tokmina-Roszyk, D.; Fields, G.B. The synthesis and application of Fmoc-Lys(5-Fam) building blocks. Biopolymers 2013, 100, 347–355. [Google Scholar] [CrossRef]
- Triglia, P.; Linscott, W.D. Titers of nine complement components, conglutinin and C3b-inactivator in adult and fetal bovine sera. Mol. Immunol. 1980, 17, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Larson, B.; Knapinska, A.; Drotleff, G.; Fields, G. Combination of a Fluorescent Substrate-Based MMP Activity Assay and Hit Pick Reading/Imaging Procedure: Efficiently screen for inhibitors of MT1-MMP activity and 3D glioma tumoroid invasion. In Agilent Application Note: Cell Migration and Invasion; Agilent: Raleigh, NC, USA, 2022; 11p. [Google Scholar]
- Vandenbroucke, R.E.; Libert, C. Is there new hope for therapeutic matrix metalloproteinase inhibition? Nat. Rev. Drug Discov. 2014, 13, 904–927. [Google Scholar] [CrossRef] [PubMed]
- Pei, D.; Weiss, S.J. Transmembrane-deletion Mutants of the Membrane-type Matrix Metalloproteinase-1 Process Progelatinase A and Express Intrinsic Matrix-degrading Activity. J. Biol. Chem. 1996, 271, 9135–9140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Kinoshita, T.; Takino, T.; Nakayama, K.; Seiki, M. Activation of a recombinant membrane type 1-matrix metalloproteinase (MT1-MMP) by furin and its interaction with tissue inhibitor of metalloproteinases (TIMP)-2. FEBS Lett. 1996, 393, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Okumura, Y.; Sato, H.; Seiki, M.; Kido, H. Proteolytic activation of the precursor of membrane type 1 matrix metalloproteinase by human plasmin: A possible cell surface activator. FEBS Lett. 1997, 402, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Qiu, Y.; Wan, H.; Sohn, M.R.; Sohn, H.D.; Jin, B.R. Antifibrinolytic Role of a Bee Venom Serine Protease Inhibitor That Acts as a Plasmin Inhibitor. PLoS ONE 2012, 7, e32269. [Google Scholar] [CrossRef] [Green Version]
- Molloy, S.; Bresnahan, P.; Leppla, S.; Klimpel, K.; Thomas, G. Human furin is a calcium-dependent serine endoprotease that recognizes the sequence Arg-X-X-Arg and efficiently cleaves anthrax toxin protective antigen. J. Biol. Chem. 1992, 267, 16396–16402. [Google Scholar] [CrossRef]
- Shimomura, T.; Ochiai, M.; Kondo, J.; Morimoto, Y. A novel protease obtained from FBS-containing culture supernatant, that processes single chain form hepatocyte growth factor to two chain form in serum-free culture. Cytotechnology 1992, 8, 219–229. [Google Scholar] [CrossRef]
- Remacle, A.G.; Golubkov, V.S.; Shiryaev, S.A.; Dahl, R.; Stebbins, J.L.; Chernov, A.V.; Cheltsov, A.V.; Pellecchia, M.; Strongin, A.Y. Novel MT1-MMP Small-Molecule Inhibitors Based on Insights into Hemopexin Domain Function in Tumor Growth. Cancer Res. 2012, 72, 2339–2349. [Google Scholar] [CrossRef] [Green Version]
- Nam, D.H.; Rodriguez, C.; Remacle, A.G.; Strongin, A.Y.; Ge, X. Active-site MMP-selective antibody inhibitors discovered from convex paratope synthetic libraries. Proc. Natl. Acad. Sci. USA 2016, 113, 14970–14975. [Google Scholar] [CrossRef]
- Hur, J.H.; Park, M.J.; Park, I.C.; Yi, D.H.; Rhee, C.H.; Hong, S.I.; Lee, S.H. Matrix metalloproteinases in human gliomas: Activation of matrix metalloproteinase-2 (MMP-2) may be correlated with membrane-type-1 matrix metalloproteinase (MT1-MMP) expression. J. Korean Med. Sci. 2000, 15, 309–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proulx-Bonneau, S.; Pratt, J.; Annabi, B. A role for MT1-MMP as a cell death sensor/effector through the regulation of endoplasmic reticulum stress in U87 glioblastoma cells. J. Neuro Oncol. 2011, 104, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Duran, C.L.; Lee, D.W.; Jung, J.-U.; Ravi, S.; Pogue, C.B.; Toussaint, L.G.; Bayless, K.J.; Sitcheran, R. NIK regulates MT1-MMP activity and promotes glioma cell invasion independently of the canonical NF-κB pathway. Oncogenesis 2016, 5, e231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thome, I.; Lacle, R.; Voß, A.; Bortolussi, G.; Pantazis, G.; Schmidt, A.; Conrad, C.; Jacob, R.; Timmesfeld, N.; Bartsch, J.W.; et al. Neoplastic Cells are the Major Source of MT-MMPs in IDH1-Mutant Glioma, Thus Enhancing Tumor-Cell Intrinsic Brain Infiltration. Cancers 2020, 12, 2456. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, T.; Theissen, S.M.; Morton, M.J.; Caplan, M.J. The Cytoplasmic Tail Dileucine Motif LL572 Determines the Glycosylation Pattern of Membrane-type 1 Matrix Metalloproteinase. J. Biol. Chem. 2008, 283, 35410–35418. [Google Scholar] [CrossRef] [Green Version]
- Knapinska, A.M.; Estrada, C.-A.; Fields, G.B. The roles of matrix metalloproteinases in pancreatic cancer. In Progress in Molecular Biology and Translational Science; (Matrix Metalloproteinases and Tissue Remodeling in Health and Disease: Target Tissues and Therapy); Khalil, R.A., Ed.; Academic Press: Cambridge, MA, USA; Elsevier, Inc.: London, UK, 2017; Volume 148, pp. 339–354. [Google Scholar]
- Pahwa, S.; Stawikowski, M.J.; Fields, G.B. Monitoring and Inhibiting MT1-MMP during Cancer Initiation and Progression. Cancers 2014, 6, 416–435. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Holtzinger, A.; Jagan, I.; BeGora, M.; Lohse, I.; Ngai, N.; Nostro, C.; Wang, R.; Muthuswamy, L.B.; Crawford, H.C.; et al. Ductal pancreatic cancer modeling and drug screening using human pluripotent stem cell-and patient-derived tumor organoids. Nat. Med. 2015, 21, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Hesley, J.; Sirenko, O. Screening for Cancer Therapeutics Using Spheroids. Genet. Eng. Biotechnol. News 2016, 36, 14–15. [Google Scholar] [CrossRef]
- Pahwa, S.; Bhowmick, M.; Amar, S.; Cao, J.; Strongin, A.Y.; Fridman, R.; Weiss, S.J.; Fields, G.B. Characterization and regulation of MT1-MMP cell surface-associated activity. Chem. Biol. Drug Des. 2019, 93, 1251–1264. [Google Scholar] [CrossRef]
- Ouyang, M.; Huang, H.; Shaner, N.C.; Remacle, A.G.; Shiryaev, S.A.; Strongin, A.Y.; Tsien, R.Y.; Wang, Y. Simultaneous Visualization of Protumorigenic Src and MT1-MMP Activities with Fluorescence Resonance Energy Transfer. Cancer Res 2010, 70, 2204–2212. [Google Scholar] [CrossRef]
- Ouyang, M.; Lu, S.; Li, X.-Y.; Xu, J.; Seong, J.; Giepmans, B.; Shyy, J.Y.-J.; Weiss, S.J.; Wang, Y. Visualization of Polarized Membrane Type 1 Matrix Metalloproteinase Activity in Live Cells by Fluorescence Resonance Energy Transfer Imaging. J. Biol. Chem. 2008, 283, 17740–17748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabaiah, A.; Daugherty, P.S. Directed Evolution of Protease Beacons that Enable Sensitive Detection of Endogenous MT1-MMP Activity in Tumor Cell Lines. Chem. Biol. 2011, 18, 392–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, Y.; Huang, H.; Pan, Y.; Chaney, E.J.; Boppart, S.; Ozer, H.; Strongin, A.Y.; Wang, Y. Quantitative FRET Imaging to Visualize the Invasiveness of Live Breast Cancer Cells. PLoS ONE 2013, 8, e58569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limsakul, P.; Peng, Q.; Wu, Y.; Allen, M.E.; Liang, J.; Remacle, A.G.; Lopez, T.; Ge, X.; Kay, B.K.; Zhao, H.; et al. Directed Evolution to Engineer Monobody for FRET Biosensor Assembly and Imaging at Live-Cell Surface. Cell Chem. Biol. 2018, 25, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Leight, J.L.; Alge, D.L.; Maier, A.J.; Anseth, K.S. Direct measurement of matrix metalloproteinase activity in 3D cellular microenvironments using a fluorogenic peptide substrate. Biomaterials 2013, 34, 7344–7352. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Chen, Y.; Wang, W.; Zhang, J.; Jiang, T.; Lu, Q. Noninvasive Bioluminescence Imaging of Matrix Metalloproteinase-14 Activity in Lung Cancer Using a Membrane-Bound Biosensor. Anal. Chem. 2021, 93, 8739–8745. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, F.; Ma, Y.; Liu, G.; Kim, K.; Fang, X.; Lee, S.; Chen, X. In Vivo Optical Imaging of Membrane-Type Matrix Metalloproteinase (MT-MMP) Activity. Mol. Pharm. 2011, 8, 2331–2338. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.-D.; Kang, K.A. Chapter 54: MMP-14 Triggered Fluorescence Contrast Agent. In Oxygen Transport to Tissue XXXVIII: Advances in Experimental Medicine and Biology 923; Luo, Q., Li, L.Z., Harrison, D.K., Shi, H., Bruley, D.F., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 413–419. [Google Scholar]
- Yamamoto, M.; Mohanam, S.; Sawaya, R.; Fuller, G.; Seiki, M.; Sato, H.; Gokaslan, Z.L.; Liotta, L.; Nicolson, G.L.; Rao, J.S. Differential expression of membrane-type matrix metalloproteinase and its correlation with gelatinase A activation in human malignant brain tumors in vivo and in vitro. Cancer Res 1996, 56, 384–392. [Google Scholar]
- Nuttall, R.K.; Pennington, C.J.; Taplin, J.; Wheal, A.; Yong, V.W.; Forsyth, P.; Edwards, D.R. Elevated membrane-type matrix metalloproteinases in gliomas revealed by profiling proteases and inhibitors in human cancer cells. Mol. Cancer Res. 2003, 1, 333–345. [Google Scholar]
- Liu, G.; Atteridge, C.L.; Wang, X.; Lundgren, A.D.; Wu, J.D. Cutting Edge: The Membrane Type Matrix Metalloproteinase MMP14 Mediates Constitutive Shedding of MHC Class I Chain-Related Molecule a Independent of a Disintegrin and Metalloproteinases. J. Immunol. 2010, 184, 3346–3350. [Google Scholar] [CrossRef] [Green Version]
- De Andrade, L.F.; Tay, R.E.; Pan, D.; Luoma, A.M.; Ito, Y.; Badrinath, S.; Tsoucas, D.; Franz, B.; May, K.F., Jr.; Harvey, C.J.; et al. Antibody-mediated inhibition of MICA and MICB shedding promotes NK cell–driven tumor immunity. Science 2018, 359, 1537–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, G.B. Protease-activated delivery and imaging systems. In The Cancer Degradome—Proteases in Cancer Biology; Edwards, D., Hoyer-Hansen, G., Blasi, F., Sloane, B., Eds.; Springer: New York, NY, USA, 2008; pp. 827–851. [Google Scholar]
- Atkinson, J.M.; Falconer, R.A.; Edwards, D.R.; Pennington, C.J.; Siller, C.S.; Shnyder, S.D.; Bibby, M.C.; Patterson, L.H.; Loadman, P.M.; Gill, J.H. Development of a Novel Tumor-Targeted Vascular Disrupting Agent Activated by Membrane-Type Matrix Metalloproteinases. Cancer Res 2010, 70, 6902–6912. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.H.; Loadman, P.M.; Shnyder, S.D.; Cooper, P.; Atkinson, J.M.; Morais, G.R.; Patterson, L.H.; Falconer, R.A. Tumor-Targeted Prodrug ICT2588 Demonstrates Therapeutic Activity against Solid Tumors and Reduced Potential for Cardiovascular Toxicity. Mol. Pharm. 2014, 11, 1294–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnieh, F.M.; Morais, G.R.; Garland, H.; Loadman, P.M.; Falconer, R.A. Targeted delivery of a colchicine analogue provides synergy with ATR inhibition in cancer cells. Biochem. Pharmacol. 2022, 201, 115095. [Google Scholar] [CrossRef]
- Ansari, C.; Tikhomirov, G.; Hong, S.H.; Falconer, R.; Loadman, P.; Gill, J.; Castaneda, R.; Hazard, F.K.; Tong, L.; Lenkov, O.D.; et al. Development of Novel Tumor-Targeted Theranostic Nanoparticles Activated by Membrane-Type Matrix Metalloproteinases for Combined Cancer Magnetic Resonance Imaging and Therapy. Small 2014, 10, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Chen, Z.; Li, K.; Morais, G.R.; Klockow, J.; Yerneni, K.; Pisani, L.; Chin, F.T.; Mitra, S.; Cheshier, S.; et al. A Novel Theranostic Strategy for MMP-14–Expressing Glioblastomas Impacts Survival. Mol. Cancer Ther. 2017, 16, 1909–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daldrup-Link, H.E. Rethinking Brain Cancer Therapy: Tumor Enzyme Activatable Theranostic Nanoparticles. Mol. Imaging 2017, 16, 1536012117730950. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Klockow, J.L.; Mohanty, S.; Ku, K.S.; Aghighi, M.; Melemenidis, S.; Chen, Z.; Li, K.; Morais, G.R.; Zhao, N.; et al. Theranostic nanoparticles enhance the response of glioblastomas to radiation. Nanotheranostics 2019, 3, 299–310. [Google Scholar] [CrossRef]
- Albright, C.F.; Graciani, N.; Han, W.; Yue, E.; Stein, R.; Lai, Z.; Diamond, M.; Dowling, R.; Grimminger, L.; Zhang, S.-Y.; et al. Matrix metalloproteinase–activated doxorubicin prodrugs inhibit HT1080 xenograft growth better than doxorubicin with less toxicity. Mol. Cancer Ther. 2005, 4, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Kasten, B.B.; Jiang, K.; Cole, D.; Jani, A.; Udayakumar, N.; Gillespie, G.Y.; Lu, G.; Dai, T.; Rosenthal, E.L.; Markert, J.M.; et al. Targeting MMP-14 for dual PET and fluorescence imaging of glioma in preclinical models. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 1412–1426. [Google Scholar] [CrossRef]
- Sood, D.; Tang-Schomer, M.; Pouli, D.; Mizzoni, C.; Raia, N.; Tai, A.; Arkun, K.; Wu, J.; Black, L.D.; Scheffler, B.; et al. 3D extracellular matrix microenvironment in bioengineered tissue models of primary pediatric and adult brain tumors. Nat. Commun. 2019, 10, 4529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullen, N.; Pickford, A.R.; Perry, M.M.; Jaworski, D.M.; Loveson, K.F.; Arthur, D.J.; Holliday, J.R.; Van Meter, T.; Peckham, R.; Younas, W.; et al. Current insights into matrix metalloproteinases and glioma progression: Transcending the degradation boundary. Met. Med. 2018, 5, 13–30. [Google Scholar] [CrossRef]
- Johansson, P.; Krona, C.; Kundu, S.; Doroszko, M.; Baskaran, S.; Schmidt, L.; Vinel, C.; Almstedt, E.; Elgendy, R.; Elfineh, L.; et al. A Patient-Derived Cell Atlas Informs Precision Targeting of Glioblastoma. Cell Rep. 2020, 32, 107897. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-B.; Karpova, A.; Gritsenko, M.A.; Kyle, J.E.; Cao, S.; Li, Y.; Rykunov, D.; Colaprico, A.; Rothstein, J.H.; Hong, R.; et al. Proteogenomic and metabolomic characterization of human glioblastoma. Cancer Cell 2021, 39, 509–528.e20. [Google Scholar] [CrossRef] [PubMed]
- Varn, F.S.; Johnson, K.C.; Martinek, J.; Huse, J.T.; Nasrallah, M.P.; Wesseling, P.; Cooper, L.A.; Malta, T.M.; Wade, T.E.; Sabedot, T.S.; et al. Glioma progression is shaped by genetic evolution and microenvironment interactions. Cell 2022, 185, 2184–2199.e16. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Time (h) | Substrate Signal (RFU) | Substrate Ratio | Spheroid Area (μm2) | Spheroid Ratio |
|---|---|---|---|---|---|
| A172 | 0 | 4.85 ± 0.13 × 104 | 1.00 | 8.22 ± 0.23 × 104 | 1.00 |
| 24 | 3.52 ± 0.03 × 105 | 7.26 ± 0.25 | 1.35 ± 0.05 × 105 | 1.64 ± 0.10 | |
| 48 | 5.77 ± 0.05 × 105 | 11.9 ± 0.22 | 1.8 ± 0.15 × 105 | 2.19 ± 0.12 | |
| H4 | 0 24 | 8.92 ± 0.23 × 104 5.12 ± 0.15 × 105 | 1.00 5.72 ± 0.02 | 2.93 ± 0.05 × 105 4.53 ± 0.08 × 105 | 1.00 1.55 ± 0.01 |
| 48 | 8.62 ± 0.11 × 105 | 9.66 ± 0.13 | 6.64 ± 0.02 × 105 | 2.27 ± 0.05 | |
| SW 1088 | 0 | 9.9 ± 0.2 × 104 | 1.00 | 1.64 ± 0.03 × 105 | 1.00 |
| 24 | 4.31 ± 0.32 × 105 | 4.35 ± 42 | 3.79 ± 0.17 × 105 | 2.31 ± 0.14 | |
| 48 | 7.62 ± 0.47 × 105 | 7.7 ± 0.65 | 4.84 ± 0.31 × 105 | 2.96 ± 0.25 |
| Cell Type | Time (h) | Substrate Signal (RFU) | Substrate Ratio | Spheroid Area (μm2) | Spheroid Ratio |
|---|---|---|---|---|---|
| A172 | 0 | 4.80 ± 0.26 × 104 | 1.00 | 6.06 ± 0.06 × 104 | 1.00 |
| 24 | 1.31 ± 0.03 × 105 | 2.73 ± 0.2 | 6.13 ± 0.07 × 104 | 1.01 ± 0.02 | |
| 48 | 2.0 ± 0.05 × 105 | 4.17 ± 0.31 | 5.57 ± 0.54 × 104 | 0.92 ± 0.1 | |
| H4 | 0 24 | 8.79 ± 0.42 × 104 1.33 ± 0.05 × 105 | 1.00 1.52 ± 0.02 | 2.28 ± 0.01 × 105 2.29 ± 0.02 × 105 | 1.00 1.01 ± 0.01 |
| 48 | 2.72 ± 0.15 × 105 | 3.10 ± 0.03 | 2.3 ± 0.02 × 105 | 1.01 ± 0.01 | |
| SW 1088 | 0 | 9.51 ± 0.22 × 104 | 1.00 | 1.46 ± 0.05 × 105 | 1.00 |
| 24 | 1.1 ± 0.07 × 105 | 1.16 ± 0.05 | 1.4 ± 0.1 × 105 | 0.97 ± 0.04 | |
| 48 | 2.69 ± 0.07 × 105 | 2.83 ± 0.14 | 1.34 ± 0.1 × 105 | 0.92 ± 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knapinska, A.M.; Drotleff, G.; Chai, C.; Twohill, D.; Ernce, A.; Tokmina-Roszyk, D.; Grande, I.; Rodriguez, M.; Larson, B.; Fields, G.B. Screening MT1-MMP Activity and Inhibition in Three-Dimensional Tumor Spheroids. Biomedicines 2023, 11, 562. https://doi.org/10.3390/biomedicines11020562
Knapinska AM, Drotleff G, Chai C, Twohill D, Ernce A, Tokmina-Roszyk D, Grande I, Rodriguez M, Larson B, Fields GB. Screening MT1-MMP Activity and Inhibition in Three-Dimensional Tumor Spheroids. Biomedicines. 2023; 11(2):562. https://doi.org/10.3390/biomedicines11020562
Chicago/Turabian StyleKnapinska, Anna M., Gary Drotleff, Cedric Chai, Destiny Twohill, Alexa Ernce, Dorota Tokmina-Roszyk, Isabella Grande, Michelle Rodriguez, Brad Larson, and Gregg B. Fields. 2023. "Screening MT1-MMP Activity and Inhibition in Three-Dimensional Tumor Spheroids" Biomedicines 11, no. 2: 562. https://doi.org/10.3390/biomedicines11020562