

The Relationship between the Oral Microbiota and Metabolic Syndrome
 , , , and
, , , and
Abstract
:1. Background
2. Methods and Materials
2.1. Study Subjects and Sample Collection
2.2. Sample Collection
2.3. Metabolic Syndrome Classification
2.4. Collection of Plaque Samples
2.5. DNA Extraction and Quality Control
2.6. Metagenomics 16S rDNA
2.7. Library Preparation
2.8. Template Preparation, Enrichment, Sequencing, and Analysis
2.9. Statistical Analysis
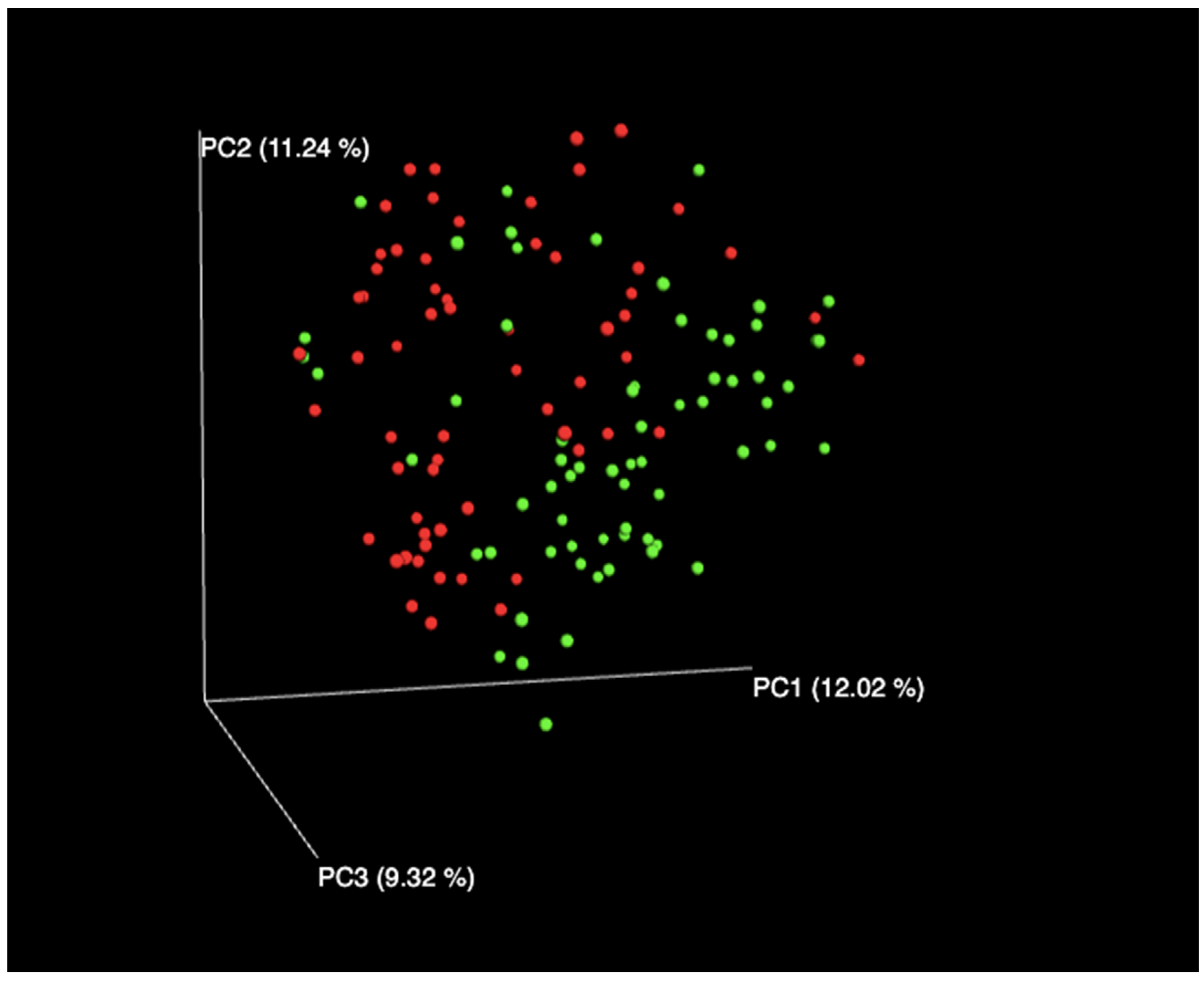
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCracken, E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the metabolic syndrome. Clin. Dermatol. 2018, 36, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the international diabetes federation task force on epidemiology and prevention; National heart, lung, and blood institute; American heart association; World heart federation; International. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Huang, S.; Xu, T.; Wang, Y.; Ju, S. Measuring Brown Fat Using MRI and Implications in the Metabolic Syndrome. J. Magn. Reson. Imaging 2020, 54, 1377–1392. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Xuan, S.; Wang, Z. Oral microbiota: A new view of body health. Food Sci. Hum. Wellness 2019, 8, 8–15. [Google Scholar] [CrossRef]
- Wade, W.G. The oral microbiome in health and disease. Pharmacol. Res. 2013, 69, 137–143. [Google Scholar] [CrossRef]
- Sharma, N.; Bhatia, S.; Sodhi, A.S.; Batra, N. Oral microbiome and health. AIMS Microbiol. 2018, 4, 42–66. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Si, J.; Lee, C.; Ko, G. Oral Microbiota: Microbial Biomarkers of Metabolic Syndrome Independent of Host Genetic Factors. Front. Cell. Infect. Microbiol. 2017, 7, 516. [Google Scholar] [CrossRef] [Green Version]
- Janket, S.-J.; Baird, A.E.; Chuang, S.-K.; Jones, J. Meta-analysis of periodontal disease and risk of coronary heart disease and stroke. Oral Surgery, Oral Med. Oral Pathol. Oral Radiol. Endodontology 2003, 95, 559–569. [Google Scholar] [CrossRef]
- Khader, Y.S.; Albashaireh, Z.S.; AlOmari, M.A. Periodontal Diseases and the Risk of Coronary Heart and Cerebrovascular Diseases: A Meta-Analysis. J. Periodontol. 2004, 75, 1046–1053. [Google Scholar] [CrossRef]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2015, 4, 4828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsha, T.; Hassan, M.; Hon, G.; Soita, D.; Kengne, A.; Erasmus, R. Derivation and validation of a waist circumference optimal cutoff for diagnosing metabolic syndrome in a South African mixed ancestry population. Int. J. Cardiol. 2013, 168, 2954–2955. [Google Scholar] [CrossRef] [PubMed]
- Keijser, B.J.F.; Zaura, E.; Huse, S.M.; Van Der Vossen, J.M.B.M.; Schuren, F.H.J.; Montijn, R.C.; ten Cate, J.M.; Crielaard, W. Pyrosequencing analysis of the Oral Microflora of healthy adults. J. Dent. Res. 2008, 87, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Saeb, A.T.; Al-Rubeaan, K.A.; Aldosary, K.; Raja, G.U.; Mani, B.; Abouelhoda, M.; Tayeb, H.T. Relative reduction of biological and phylogenetic diversity of the oral microbiota of diabetes and pre-diabetes patients. Microb. Pathog. 2019, 128, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Vielkind, P.; Jentsch, H.; Eschrich, K.; Rodloff, A.C.; Stingu, C.-S. Prevalence of Actinomyces spp. in patients with chronic periodontitis. Int. J. Med. Microbiol. 2015, 305, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Beutler, J.; Jentsch, H.F.R.; Rodloff, A.C.; Stingu, C.-S. Bacteremia after professional mechanical plaque removal in patients with chronic periodontitis. Oral Dis. 2019, 25, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Watanabe, K.; Kumada, H.; Toyama, T.; Tani-Ishii, N.; Hamada, N. Peptidoglycan of Actinomyces naeslundii induces inflammatory cytokine production and stimulates osteoclastogenesis in alveolar bone resorption. Arch. Oral Biol. 2012, 57, 1522–1528. [Google Scholar] [CrossRef]
- Linden, G.J.; Lyons, A.; Scannapieco, F.A. Periodontal systemic associations: Review of the evidence. J. Periodontol. 2013, 84, S8–S19. [Google Scholar] [CrossRef]
- Lamster, I.B.; Pagan, M. Periodontal disease and the metabolic syndrome. Int. Dent. J. 2017, 67, 67–77. [Google Scholar] [CrossRef]
- Valour, F.; Sénéchal, A.; Dupieux, C.; Karsenty, J.; Lustig, S.; Breton, P.; Gleizal, A.; Boussel, L.; Laurent, F.; Braun, E.; et al. Actinomycosis: Etiology, clinical features, diagnosis, treatment, and management. Infect. Drug Resist. 2014, 7, 183–197. [Google Scholar] [CrossRef]
- Zhong, E.F.; Chang, A.; Stucky, A.; Chen, X.; Mundluru, T.; Khalifeh, M.; Sedghizadeh, P.P. Genomic Analysis of Oral Lichen Planus and Related Oral Microbiome Pathogens. Pathogens 2020, 9, 952. [Google Scholar] [CrossRef] [PubMed]
- Tefiku, U.; Popovska, M.; Cana, A.; Zendeli-Bedxeti, L.; Recica, B.; Spasovska-Gjorgovska, A.; Spasovski, S. Determination of the Role of Fusobacterium Nucleatum in the Pathogenesis in and Out the Mouth. Prilozi 2020, 41, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Krisanaprakornkit, S.; Kimball, J.R.; Weinberg, A.; Darveau, R.P.; Bainbridge, B.W.; Dale, B.A. Inducible expression of human β-defensin 2 by Fusobacterium nucleatum in oral epithelial cells: Multiple signaling pathways and role of commensal bacteria in innate immunity and the epithelial barrier. Infect. Immun. 2000, 68, 2907–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.; Kim, Y.; Min, B.-M.; Han, S.H.; Choi, Y. Innate immune responses of gingival epithelial cells to nonperiodontopathic and periodontopathic bacteria. J. Periodontal. Res. 2007, 42, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kho, S.-A.; Choi, Y.S.; Kim, Y.C.; Rhyu, I.-C.; Choi, Y. Antibody and T Cell Responses to Fusobacterium nucleatum and Treponema denticola in Health and Chronic Periodontitis. PLoS ONE 2013, 8, e53703. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.J.; Murphy, K.E.; Fernandez, M.L. Impact of Obesity and Metabolic Syndrome on Immunity. Adv. Nutr. Int. Rev. J. 2016, 7, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Ferroni, P.; Roselli, M.; Riondino, S.; Cavaliere, F.; Guadagni, F. Insulin resistance as a predictor of venous thromboembolism in breast cancer. Endocr.-Relat. Cancer 2016, 23, L25–L28. [Google Scholar] [CrossRef] [Green Version]
- Feres FTeles RTeles LC Figueiredo, M.F. The subgingival periodontal microbiota in the aging mouth Magda. Physiol. Behav. 2017, 176, 139–148. [Google Scholar]
- Preza, D.; Olsen, I.; Willumsen, T.; Boches, S.K.; Cotton, S.L.; Grinde, B.; Paster, B.J. Microarray analysis of the microflora of root caries in elderly. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 28, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Macuch, P.; Tanner, A. Campylobacter Species in Health, Gingivitis, and Periodontitis. J. Dent. Res. 2000, 79, 785–792. [Google Scholar] [CrossRef]
- Haririan, H.; Andrukhov, O.; Bertl, K.; Lettner, S.; Kierstein, S.; Moritz, A.; Rausch-Fan, X. Microbial Analysis of Subgingival Plaque Samples Compared to That of Whole Saliva in Patients with Periodontitis. J. Periodontol. 2014, 85, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Henne, K.; Fuchs, F.; Kruth, S.; Horz, H.-P.; Conrads, G. Shifts in Campylobacter species abundance may reflect general microbial community shifts in periodontitis progression. J. Oral Microbiol. 2014, 6, 25874. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Zheng, D.; Lucas, A.R.; Kesavalu, L. Polymicrobial Oral Infection with Four Periodontal Bacteria Orchestrates a Distinct Inflammatory Response and Atherosclerosis in ApoEnull Mice. PLoS ONE 2015, 10, e0143291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yost, S.; Duran-Pinedo, A.E.; Teles, R.; Krishnan, K.; Frias-Lopez, J. Functional signatures of oral dysbiosis during periodontitis progression revealed by microbial metatranscriptome analysis. Genome Med. 2015, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Links between atherosclerotic and periodontal disease. Exp. Mol. Pathol. 2016, 100, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Uchida, F.; Oh, S.; Shida, T.; Suzuki, H.; Yamagata, K.; Mizokami, Y.; Bukawa, H.; Tanaka, K.; Shoda, J. Effects of Exercise on the Oral Microbiota and Saliva of Patients with Non-Alcoholic Fatty Liver Disease. Int. J. Environ. Res. Public Health 2021, 18, 3470. [Google Scholar] [CrossRef]


{kind=link}
| JIS = No | JIS = Yes | p-Value | |
|---|---|---|---|
| (n = 66) | (n = 62) | ||
| Age (years) | 44.24 (14.55) | 49.98 (10.45) | 0.012 |
| BMI (kg/m2) | 25.55 (6.12) | 37.82 (8.72) | <0.001 |
| Waist-C (cm) | 80.70 (12.63) | 110.5 (14.69) | <0.001 |
| Hip (cm) | 92.07 (13.00) | 119.3 (18.03) | <0.001 |
| SBP (mmHg) | 126.1 (23.46) | 133.3 (22.41) | 0.079 |
| DBP (mmHg) | 81.80 (14.60) | 87.24 (12.32) | 0.025 |
| Glucose Fasting Blood (mmol/L) | 4.90 (4.50; 5.40) | 7.30 (6.15; 11.05) | <0.001 |
| Glucose2HRsPostPrandial (mmol/L) | 6.95 (5.20; 8.80) | 11.80 (9.90; 15.50) | <0.001 |
| GlycatedHBHbA1cTrial (%) | 5.50 (5.28; 5.93) | 7.30 (6.58; 9.30) | <0.001 |
| Insulin Fasting (mIU/L) | 5.00 (3.40; 8.40) | 15.90 (8.15; 22.40) | <0.001 |
| Insulin120Minutes (mIU/L) | 32.10 (19.18; 64.15) | 67.50 (39.10; 110.30) | 0.003 |
| TriglyceridesS (mmol/L) | 1.03 (0.77; 1.32) | 1.63 (1.25; 2.19) | <0.001 |
| LDL Cholesterol (mmol/L) | 2.65 (2.20; 3.23) | 3.40 (3.00; 4.15) | <0.001 |
| Cholesterol HDLS (mmol/L) | 1.50 (1.20; 1.80) | 5.20 (4.70; 6.13) | <0.001 |
| CholesterolS (mmol/L) | 4.90 (4.10; 5.53) | 4.90 (4.10; 5.53) | 0.059 |
| CRP (mg/L) | 2.24 (1.38; 5.72) | 7.19 (3.70; 15.18) | <0.001 |
| Gamma GTS (IU/L) | 32.50 (21.00; 54.00) | 34.00 (25.75; 65.25) | 0.228 |
| Gender | 0.049 | ||
| Female (n = 93) | 43 (65%) | 50 (81%) | |
| Male (n = 35) | 23 (35%) | 12 (19%) | |
| Gingival bleeding | 0.964 | ||
| No (n = 44) | 23 (35%) | 21 (35%) | |
| Yes (n = 81) | 42 (65%) | 39 (65) | |
| Periodontitis | 0.499 | ||
| No (n = 56) | 31 (48%) | 25 (42%) | |
| Yes (n = 69) | 34 (52%) | 35 (58%) |
| Metabolic Syndrome | ||
|---|---|---|
| No | Yes | |
| Number of taxa | 253 | 275 |
| Shannon | 4.207 | 4.239 |
| Chao1 | 253 | 275 |
| Simpson | 0.0296 | 0.0297 |
| MetS | Periodontitis | Bleeding | |||||||
|---|---|---|---|---|---|---|---|---|---|
| No | Yes | p-Value | No | Yes | p-Value | No | Yes | p-Value | |
| Actinomyces dentalis | 1.59 | 2.80 | <0.001 | 1.79 | 2.54 | 0.117 | 2.07 | 2.28 | 0.513 |
| Actinomyces naeslundii | 1.48 | 4.03 | <0.001 | 2.58 | 2.15 | 0.166 | 2.93 | 1.99 | 0.056 |
| Actinomyces odontolyticus | 1.04 | 0.67 | 0.005 | ||||||
| Actinomyces viscosus | 0.27 | 1.58 | 0.021 | ||||||
| Aggregatibacter segnis | 2.52 | 1.81 | 0.174 | 2.13 | 2.08 | 0.438 | 1.33 | 2.56 | 0.152 |
| Campylobacter gracilis | 3.71 | 2.22 | 0.002 | 2.94 | 3.07 | 0.984 | 3.35 | 2.81 | 0.123 |
| Capnocytophaga leadbetteri | 1.34 | 1.01 | 0.219 | 1.02 | 1.23 | 0.607 | 1.12 | 1.14 | 0.802 |
| Corynebacterium matruchotii | 2.97 | 7.91 | <0.001 | 5.94 | 5.07 | 0.281 | 5.72 | 5.31 | 0.706 |
| Fusobacterium canifelinum | 1.22 | 0.72 | 0.001 | 1.13 | 0.81 | 0.187 | |||
| Fusobacterium nucleatum | 7.09 | 4.62 | <0.001 | 6.23 | 5.68 | 0.447 | 4.49 | 6.78 | 0.001 |
| Fusobacterium periodonticum | 1.68 | 0.38 | 0.022 | 0.95 | 1.17 | 0.328 | 0.72 | 1.27 | 0.249 |
| Granulicatella adiacens | 1.13 | 0.69 | 0.016 | ||||||
| Haemophilus parainfluenzae | 10.10 | 3.94 | <0.001 | 7.78 | 6.75 | 0.712 | 9.08 | 6.11 | 0.051 |
| Leptotrichia buccalis | 1.45 | 2.51 | 0.007 | 1.52 | 2.38 | 0.356 | 2.41 | 1.75 | 0.592 |
| Leptotrichia genomosp. | 0.97 | 1.84 | 0.183 | 1.12 | 1.63 | 0.538 | 1.39 | 1.41 | 0.329 |
| Leptotrichia hofstadii | 0.49 | 1.01 | 0.184 | 1.06 | 0.58 | 0.343 | |||
| Leptotrichia hongkongensis | 1.33 | 0.47 | 0.261 | 1.47 | 0.49 | 0.236 | |||
| Leptotrichia wadei | 1.23 | 1.43 | 0.327 | 1.18 | 1.49 | 0.180 | 1.24 | 1.42 | 0.495 |
| Mannheimia varigena | 3.54 | 2.03 | 0.075 | 2.86 | 2.80 | 0.150 | 2.99 | 2.74 | 0.462 |
| Neisseria flavescens | 1.19 | 0.57 | 0.835 | 0.57 | 1.02 | 0.036 | |||
| Other * | 38.22 | 40.17 | 0.118 | 41.21 | 41.56 | 0.646 | 38.50 | 40.62 | 0.528 |
| Prevotella histicola | 0.39 | 1.00 | 0.225 | ||||||
| Prevotella maculosa | 1.00 | 0.91 | 0.763 | 1.03 | 0.90 | 0.197 | |||
| Prevotella melaninogenica | 3.71 | 4.13 | 0.405 | 4.09 | 3.92 | 0.514 | 2.94 | 4.62 | 0.124 |
| Prevotella oris | 0.73 | 1.34 | 0.160 | 1.09 | 1.02 | 0.227 | 1.37 | 0.86 | 0.059 |
| Prevotella oulorum | 1.32 | 0.75 | 0.069 | 1.06 | 1.06 | 0.255 | 1.50 | 0.80 | 0.326 |
| Prevotella pallens | 1.39 | 0.90 | 0.078 | 1.11 | 1.18 | 0.983 | 0.99 | 1.24 | 0.722 |
| Prevotella veroralis | 1.17 | 1.09 | 0.818 | 1.23 | 1.08 | 0.787 | 0.68 | 1.43 | 0.084 |
| Selenomonas noxia | 1.19 | 1.04 | 0.534 | 0.92 | 1.30 | 0.273 | 1.55 | 0.88 | 0.001 |
| Streptococcus gordonii | 1.03 | 0.67 | 0.339 | ||||||
| Streptococcus mutans | 0.33 | 1.39 | 0.269 | 1.08 | 0.81 | 0.836 | |||
| Streptococcus sanguinis | 0.43 | 1.56 | <0.001 | 1.35 | 0.65 | 0.209 | 1.76 | 0.50 | 0.007 |
| Veillonella alcalescens | 3.96 | 4.37 | 0.954 | 3.94 | 4.43 | 0.414 | 3.62 | 4.56 | 0.541 |
| Veillonella parvula | 1.08 | 0.94 | 0.140 | 1.03 | 1.02 | 0.933 | 0.92 | 1.08 | 0.919 |
| Veillonella rogosae | 1.37 | 0.89 | 0.007 | 1.13 | 1.16 | 0.757 | 0.87 | 1.31 | 0.037 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prince, Y.; Davison, G.M.; Davids, S.F.G.; Erasmus, R.T.; Kengne, A.P.; Graham, L.M.; Raghubeer, S.; Matsha, T.E. The Relationship between the Oral Microbiota and Metabolic Syndrome. Biomedicines 2023, 11, 3. https://doi.org/10.3390/biomedicines11010003
Prince Y, Davison GM, Davids SFG, Erasmus RT, Kengne AP, Graham LM, Raghubeer S, Matsha TE. The Relationship between the Oral Microbiota and Metabolic Syndrome. Biomedicines. 2023; 11(1):3. https://doi.org/10.3390/biomedicines11010003
Chicago/Turabian StylePrince, Yvonne, Glenda M. Davison, Saarah F. G. Davids, Rajiv T. Erasmus, Andre P. Kengne, Lisa M. Graham, Shanel Raghubeer, and Tandi E. Matsha. 2023. "The Relationship between the Oral Microbiota and Metabolic Syndrome" Biomedicines 11, no. 1: 3. https://doi.org/10.3390/biomedicines11010003