
High Glucose Induces in HK2 Kidney Cells an IFN–Dependent ZIKV Antiviral Status Fueled by Viperin

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus, Plasmids, and Reagents
2.2. RT-qPCR
2.3. Transfection Experiment
2.4. Immunofluorescence Assay
2.5. Flow Cytometry Analysis
2.6. MTT Assay
2.7. LDH Assay
2.8. Western Blot Assay
2.9. Quantification of Type I IFN by SEAP Activity
2.10. ELISA Assay
2.11. Statistical Analysis
3. Results
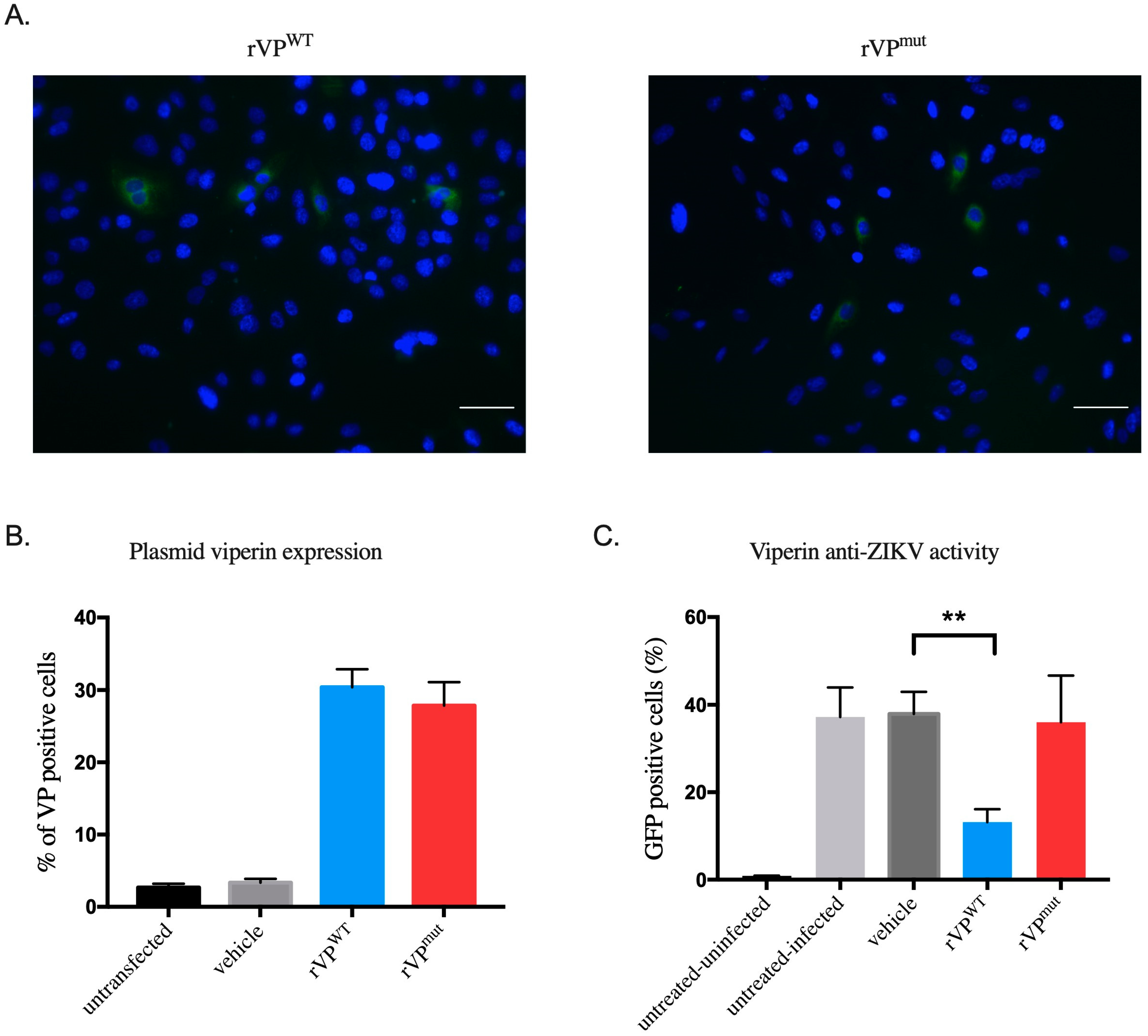
3.1. HK-2HGC Cells Display a High VP Protein Expression Level
3.2. Overexpression of Viperin in HK-2NGC Cells Inhibits ZIKV Infection
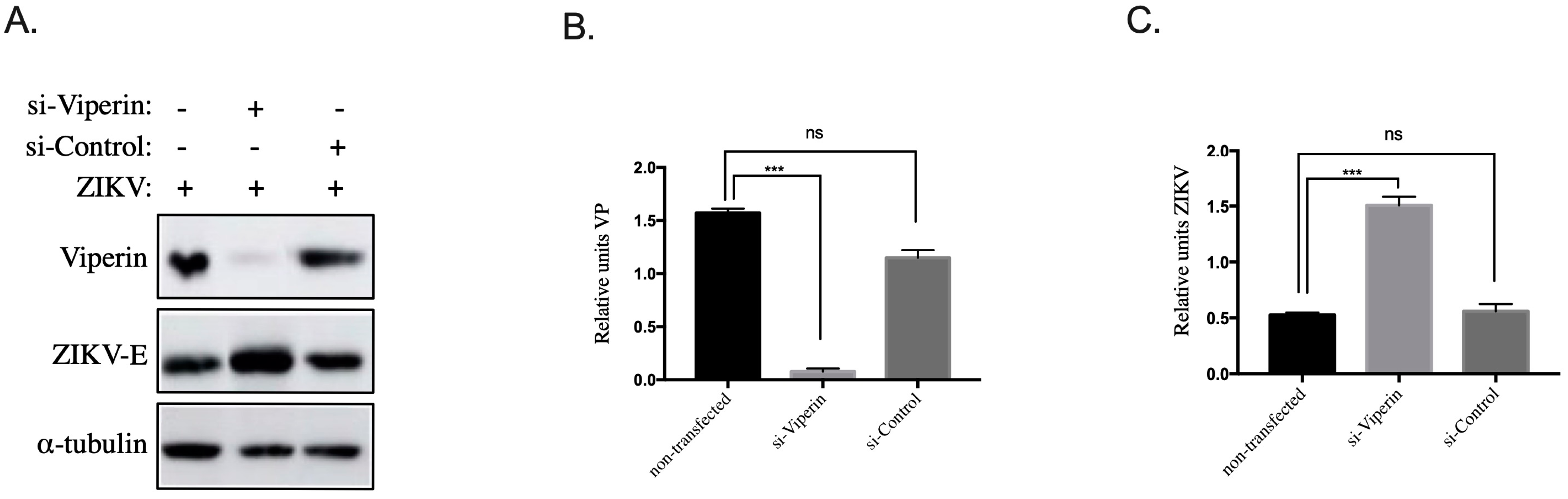
3.3. Viperin Knockdown in HK-2HGC Cells Rescues ZIKV Replication
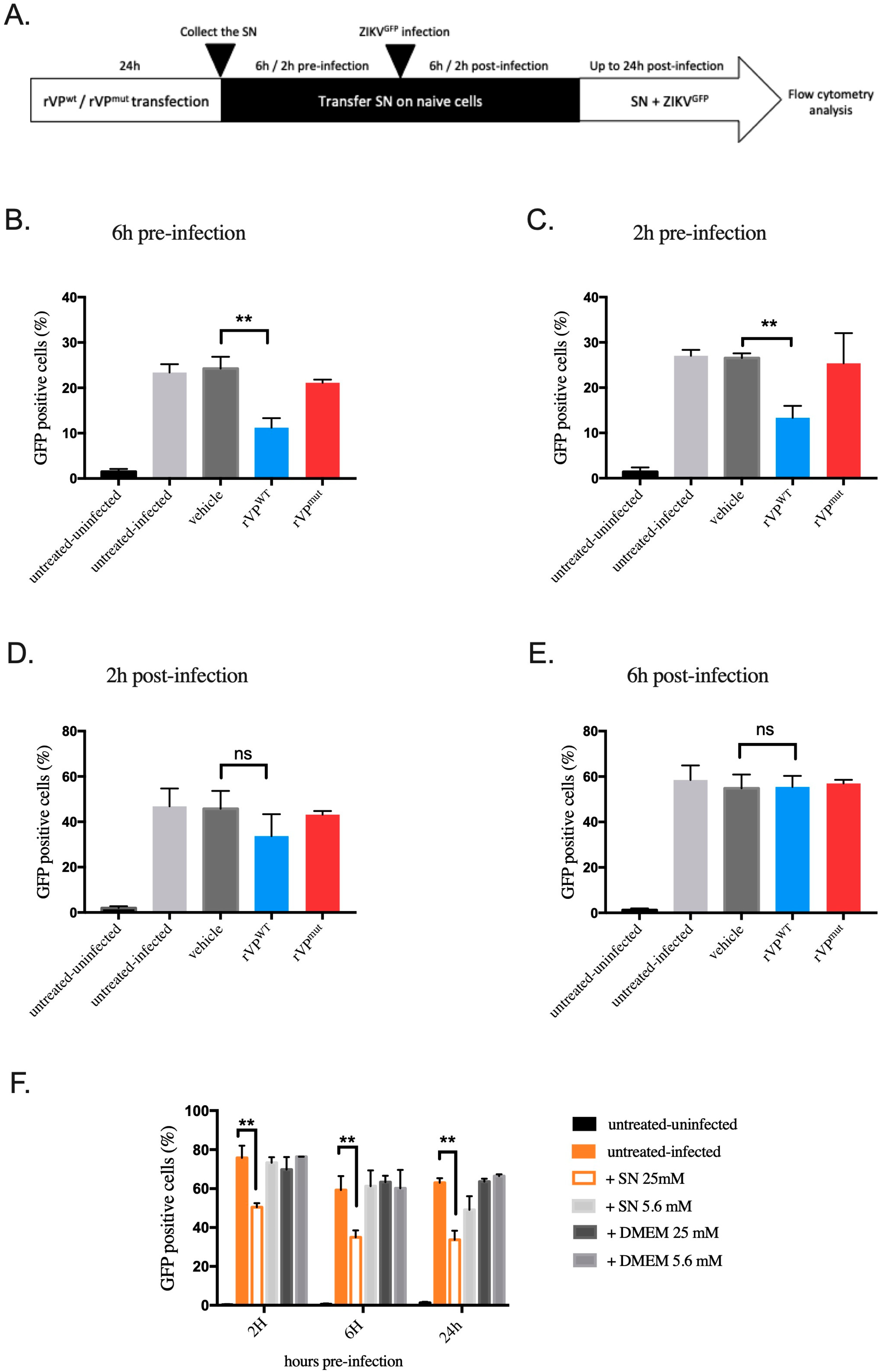
3.4. Paracrine Antiviral Effect of Viperin in HK-2HGC Cells
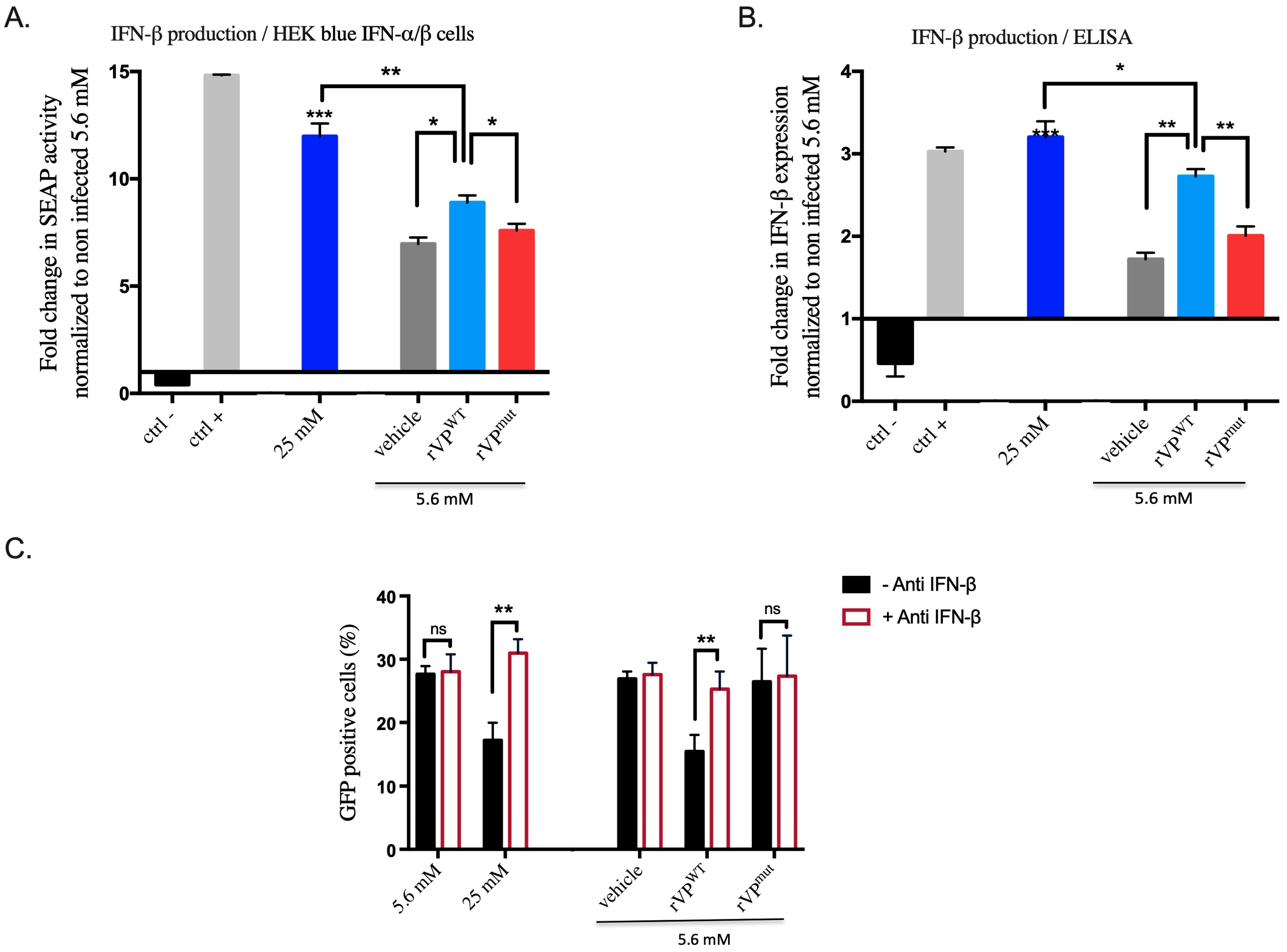
3.5. Viperin Has a Paracrine Function through IFN-β in HK-2 Cells under High Glucose Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rivera-Serrano, E.E.; Gizzi, A.S.; Arnold, J.J.; Grove, T.L.; Almo, S.C.; Cameron, C.E. Viperin Reveals Its True Function. Annu. Rev. Virol. 2020, 7, 421–446. [Google Scholar] [CrossRef]
- Boudinot, P.; Riffault, S.; Salhi, S.; Carrat, C.; Sedlik, C.; Mahmoudi, N.; Charley, B.; Benmansour, A. Vesicular Stomatitis Virus and Pseudorabies Virus Induce a Vig1/Cig5 Homologue in Mouse Dendritic Cells via Different Pathways. J. Gen. Virol. 2000, 81, 2675–2682. [Google Scholar] [CrossRef] [PubMed]
- Rivieccio, M.A.; Suh, H.-S.; Zhao, Y.; Zhao, M.-L.; Chin, K.C.; Lee, S.C.; Brosnan, C.F. TLR3 Ligation Activates an Antiviral Response in Human Fetal Astrocytes: A Role for Viperin/Cig5. J. Immunol. 2006, 177, 4735–4741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severa, M.; Coccia, E.M.; Fitzgerald, K.A. Toll-like Receptor-Dependent and -Independent Viperin Gene Expression and Counter-Regulation by PRDI-Binding Factor-1/BLIMP1. J. Biol. Chem. 2006, 281, 26188–26195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandvaux, N.; Servant, M.J.; TenOever, B.; Sen, G.C.; Balachandran, S.; Barber, G.N.; Lin, R.; Hiscott, J. Transcriptional Profiling of Interferon Regulatory Factor 3 Target Genes: Direct Involvement in the Regulation of Interferon-Stimulated Genes. J. Virol. 2002, 76, 5532–5539. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.-L.; Chang, T.-H.; Liao, C.-L.; Lin, Y.-L. The Cellular Antiviral Protein Viperin Is Attenuated by Proteasome-Mediated Protein Degradation in Japanese Encephalitis Virus-Infected Cells. J. Virol. 2008, 82, 10455–10464. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.-C.; Cresswell, P. Viperin (Cig5), an IFN-Inducible Antiviral Protein Directly Induced by Human Cytomegalovirus. Proc. Natl. Acad. Sci. USA 2001, 98, 15125–15130. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Cong, J.-P.; Shenk, T. Use of Differential Display Analysis to Assess the Effect of Human Cytomegalovirus Infection on the Accumulation of Cellular RNAs: Induction of Interferon-Responsive RNAs. Proc. Natl. Acad. Sci. USA 1997, 94, 13985–13990. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hinson, E.R.; Cresswell, P. The Interferon-Inducible Protein Viperin Inhibits Influenza Virus Release by Perturbing Lipid Rafts. Cell Host Microbe 2007, 2, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Helbig, K.J.; Eyre, N.S.; Yip, E.; Narayana, S.; Li, K.; Fiches, G.; McCartney, E.M.; Jangra, R.K.; Lemon, S.M.; Beard, M.R. The Antiviral Protein Viperin Inhibits Hepatitis C Virus Replication via Interaction with Nonstructural Protein 5A. Hepatology 2011, 54, 1506–1517. [Google Scholar] [CrossRef]
- Zhang, Y.; Burke, C.W.; Ryman, K.D.; Klimstra, W.B. Identification and Characterization of Interferon-Induced Proteins That Inhibit Alphavirus Replication. J. Virol. 2007, 81, 11246–11255. [Google Scholar] [CrossRef] [Green Version]
- Nasr, N.; Maddocks, S.; Turville, S.G.; Harman, A.N.; Woolger, N.; Helbig, K.J.; Wilkinson, J.; Bye, C.R.; Wright, T.K.; Rambukwelle, D.; et al. HIV-1 Infection of Human Macrophages Directly Induces Viperin Which Inhibits Viral Production. Blood 2012, 120, 778–788. [Google Scholar] [CrossRef]
- Jiang, D.; Weidner, J.M.; Qing, M.; Pan, X.-B.; Guo, H.; Xu, C.; Zhang, X.; Birk, A.; Chang, J.; Shi, P.-Y.; et al. Identification of Five Interferon-Induced Cellular Proteins That Inhibit West Nile Virus and Dengue Virus Infections. J. Virol. 2010, 84, 8332–8341. [Google Scholar] [CrossRef] [Green Version]
- Noppakunmongkolchai, W.; Poyomtip, T.; Jittawuttipoka, T.; Luplertlop, N.; Sakuntabhai, A.; Chimnaronk, S.; Jirawatnotai, S.; Tohtong, R. Inhibition of Protein Kinase C Promotes Dengue Virus Replication. Virol. J. 2016, 13, 35. [Google Scholar] [CrossRef] [Green Version]
- Van der Hoek, K.H.; Eyre, N.S.; Shue, B.; Khantisitthiporn, O.; Glab-Ampi, K.; Carr, J.M.; Gartner, M.J.; Jolly, L.A.; Thomas, P.Q.; Adikusuma, F.; et al. Viperin Is an Important Host Restriction Factor in Control of Zika Virus Infection. Sci. Rep. 2017, 7, 4475. [Google Scholar] [CrossRef] [Green Version]
- Panayiotou, C.; Lindqvist, R.; Kurhade, C.; Vonderstein, K.; Pasto, J.; Edlund, K.; Upadhyay, A.S.; Överby, A.K. Viperin Restricts Zika Virus and Tick-Borne Encephalitis Virus Replication by Targeting NS3 for Proteasomal Degradation. J. Virol. 2018, 92, e02054-17. [Google Scholar] [CrossRef] [Green Version]
- Vanwalscappel, B.; Gadea, G.; Desprès, P. A Viperin Mutant Bearing the K358R Substitution Lost Its Anti-ZIKA Virus Activity. Int. J. Mol. Sci. 2019, 20, 1574. [Google Scholar] [CrossRef] [Green Version]
- Gizzi, A.S.; Grove, T.L.; Arnold, J.J.; Jose, J.; Jangra, R.K.; Garforth, S.J.; Du, Q.; Cahill, S.M.; Dulyaninova, N.G.; Love, J.D.; et al. A Naturally Occurring Antiviral Ribonucleotide Encoded by the Human Genome. Nature 2018, 558, 610–614. [Google Scholar] [CrossRef]
- Vonderstein, K.; Nilsson, E.; Hubel, P.; Nygård, S.L.; Upadhyay, A.; Pasto, J.; Pichlmair, A.; Lundmark, R.; Överby, A.K. Viperin Targets Flavivirus Virulence by Inducing Assembly of Noninfectious Capsid Particles. J. Virol. 2018, 92, e01751-17. [Google Scholar] [CrossRef] [Green Version]
- Eom, J.; Kim, J.J.; Yoon, S.G.; Jeong, H.; Son, S.; Lee, J.B.; Yoo, J.; Seo, H.J.; Cho, Y.; Kim, K.S.; et al. Intrinsic Expression of Viperin Regulates Thermogenesis in Adipose Tissues. Proc. Natl. Acad. Sci. USA 2019, 116, 17419–17428. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.-Y.; Cresswell, P. Viperin Regulates Cellular Lipid Metabolism during Human Cytomegalovirus Infection. PLoS Pathog. 2013, 9, e1003497. [Google Scholar] [CrossRef]
- Ghosh, S.; Marsh, E.N.G. Viperin: An Ancient Radical SAM Enzyme Finds Its Place in Modern Cellular Metabolism and Innate Immunity. J. Biol. Chem. 2020, 295, 11513–11528. [Google Scholar] [CrossRef]
- Reslan, A.; Haddad, J.G.; Moukendza Koundi, L.; Desprès, P.; Bascands, J.-L.; Gadea, G. Zika Virus Growth in Human Kidney Cells Is Restricted by an Elevated Glucose Level. Int. J. Mol. Sci. 2021, 22, 2495. [Google Scholar] [CrossRef]
- Ali, Q.; Sabuhi, R.; Hussain, T. High Glucose Up-Regulates Angiotensin II Subtype 2 Receptors via Interferon Regulatory Factor-1 in Proximal Tubule Epithelial Cells. Mol. Cell. Biochem. 2010, 344, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Chien, A.; Jialal, I. Hyperglycemia Induces Toll like Receptor 4 Expression and Activity in Mouse Mesangial Cells: Relevance to Diabetic Nephropathy. Am. J. Physiol.-Ren. Physiol. 2012, 303, F1145–F1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hills, C.E.; Siamantouras, E.; Smith, S.W.; Cockwell, P.; Liu, K.-K.; Squires, P.E. TGFβ Modulates Cell-to-Cell Communication in Early Epithelial-to-Mesenchymal Transition. Diabetologia 2012, 55, 812–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaghobian, D.; Don, A.S.; Yaghobian, S.; Chen, X.; Pollock, C.A.; Saad, S. Increased Sphingosine 1-Phosphate Mediates Inflammation and Fibrosis in Tubular Injury in Diabetic Nephropathy. Clin. Exp. Pharmacol. Physiol. 2016, 43, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Pontrelli, P.; Conserva, F.; Papale, M.; Oranger, A.; Barozzino, M.; Vocino, G.; Rocchetti, M.T.; Gigante, M.; Castellano, G.; Rossini, M.; et al. Lysine 63 Ubiquitination Is Involved in the Progression of Tubular Damage in Diabetic Nephropathy. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Yan, R.; Wan, Q.; Lv, B.; Yang, Y.; Lv, T.; Xin, W. Inhibitor of Growth 2 Regulates the High Glucose-Induced Cell Cycle Arrest and Epithelial-to-Mesenchymal Transition in Renal Proximal Tubular Cells. J. Physiol. Biochem. 2020, 76, 373–382. [Google Scholar] [CrossRef]
- Wei, C.; Zheng, C.; Sun, J.; Luo, D.; Tang, Y.; Zhang, Y.; Ke, X.; Liu, Y.; Zheng, Z.; Wang, H. Viperin Inhibits Enterovirus A71 Replication by Interacting with Viral 2C Protein. Viruses 2018, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Gadea, G.; Bos, S.; Krejbich-Trotot, P.; Clain, E.; Viranaicken, W.; El-Kalamouni, C.; Mavingui, P.; Desprès, P. A Robust Method for the Rapid Generation of Recombinant Zika Virus Expressing the GFP Reporter Gene. Virology 2016, 497, 157–162. [Google Scholar] [CrossRef]
- Yuan, Y.; Miao, Y.; Qian, L.; Zhang, Y.; Liu, C.; Liu, J.; Zuo, Y.; Feng, Q.; Guo, T.; Zhang, L.; et al. Targeting UBE4A Revives Viperin Protein in Epithelium to Enhance Host Antiviral Defense. Mol. Cell 2020, 77, 734–747.e7. [Google Scholar] [CrossRef]
- Kumar, A.; Hou, S.; Airo, A.M.; Limonta, D.; Mancinelli, V.; Branton, W.; Power, C.; Hobman, T.C. Zika Virus Inhibits Type-I Interferon Production and Downstream Signaling. EMBO Rep. 2016, 17, 1766–1775. [Google Scholar] [CrossRef] [Green Version]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef]
- DeFilippis, V.R.; Robinson, B.; Keck, T.M.; Hansen, S.G.; Nelson, J.A.; Früh, K.J. Interferon Regulatory Factor 3 Is Necessary for Induction of Antiviral Genes during Human Cytomegalovirus Infection. J. Virol. 2006, 80, 1032–1037. [Google Scholar] [CrossRef] [Green Version]
- Stirnweiss, A.; Ksienzyk, A.; Klages, K.; Rand, U.; Grashoff, M.; Hauser, H.; Kröger, A. IFN Regulatory Factor-1 Bypasses IFN-Mediated Antiviral Effects through Viperin Gene Induction. J. Immunol. 2010, 184, 5179–5185. [Google Scholar] [CrossRef]
- Dumbrepatil, A.B.; Ghosh, S.; Zegalia, K.A.; Malec, P.A.; Hoff, J.D.; Kennedy, R.T.; Marsh, E.N.G. Viperin Interacts with the Kinase IRAK1 and the E3 Ubiquitin Ligase TRAF6, Coupling Innate Immune Signaling to Antiviral Ribonucleotide Synthesis. J. Biol. Chem. 2019, 294, 6888–6898. [Google Scholar] [CrossRef]
- Saitoh, T.; Satoh, T.; Yamamoto, N.; Uematsu, S.; Takeuchi, O.; Kawai, T.; Akira, S. Antiviral Protein Viperin Promotes Toll-like Receptor 7- and Toll-like Receptor 9-Mediated Type I Interferon Production in Plasmacytoid Dendritic Cells. Immunity 2011, 34, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Moschonas, A.; Ioannou, M.; Eliopoulos, A.G. CD40 Stimulates a “Feed-Forward” NF-ΚB–Driven Molecular Pathway That Regulates IFN-β Expression in Carcinoma Cells. J. Immunol. 2012, 188, 5521–5527. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Wang, H.; Li, Y.; Leng, X.; Huang, W.; Ma, Y.; Xu, T.; Qi, X. Transfection Reagent Lipofectamine Triggers Type I Interferon Signaling Activation in Macrophages. Immunol. Cell Biol. 2019, 97, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Alcendor, D.J. Zika Virus Infection of the Human Glomerular Cells: Implications for Viral Reservoirs and Renal Pathogenesis. J. Infect. Dis. 2017, 216, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Alcendor, D.J. Zika Virus Infection and Implications for Kidney Disease. J. Mol. Med. 2018, 96, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Burdmann, E.A. Flaviviruses and Kidney Diseases. Adv. Chronic Kidney Dis. 2019, 26, 198–206. [Google Scholar] [CrossRef]
- Murray, K.O.; Kolodziej, S.; Ronca, S.E.; Gorchakov, R.; Navarro, P.; Nolan, M.S.; Podoll, A.; Finkel, K.; Mandayam, S. Visualization of West Nile Virus in Urine Sediment Using Electron Microscopy and Immunogold up to Nine Years Postinfection. Am. J. Trop. Med. Hyg. 2017, 97, 1913–1919. [Google Scholar] [CrossRef] [Green Version]
- Saxena, V.; Xie, G.; Li, B.; Farris, T.; Welte, T.; Gong, B.; Boor, P.; Wu, P.; Tang, S.-J.; Tesh, R.; et al. A Hamster-Derived West Nile Virus Isolate Induces Persistent Renal Infection in Mice. PLoS Negl. Trop. Dis. 2013, 7, e2275. [Google Scholar] [CrossRef]
- Thomas, E.T.A.; George, J.; Sruthi, D.; Vineetha, N.S.; Gracious, N. Clinical Course of Dengue Fever and Its Impact on Renal Function in Renal Transplant Recipients and Patients with Chronic Kidney Disease. Nephrology 2019, 24, 564–568. [Google Scholar] [CrossRef]
- Gurugama, P.; Jayarajah, U.; Wanigasuriya, K.; Wijewickrama, A.; Perera, J.; Seneviratne, S.L. Renal Manifestations of Dengue Virus Infections. J. Clin. Virol. 2018, 101, 1–6. [Google Scholar] [CrossRef]
- Benfield, T.; Jensen, J.S.; Nordestgaard, B.G. Influence of Diabetes and Hyperglycaemia on Infectious Disease Hospitalisation and Outcome. Diabetologia 2007, 50, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Saqib, M.A.N.; Rafique, I.; Bashir, S.; Salam, A.A. A Retrospective Analysis of Dengue Fever Case Management and Frequency of Co-Morbidities Associated with Deaths. BMC Res. Notes 2014, 7, 205. [Google Scholar] [CrossRef] [Green Version]
- Htun, N.S.N.; Odermatt, P.; Eze, I.C.; Boillat-Blanco, N.; D’Acremont, V.; Probst-Hensch, N. Is Diabetes a Risk Factor for a Severe Clinical Presentation of Dengue? Review and Meta-Analysis. PLoS Negl. Trop. Dis. 2015, 9, e0003741. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Roe, K.; Nerurkar, P.V.; Namekar, M.; Orillo, B.; Verma, S.; Nerurkar, V.R. Impaired Virus Clearance, Compromised Immune Response and Increased Mortality in Type 2 Diabetic Mice Infected with West Nile Virus. PLoS ONE 2012, 7, e44682. [Google Scholar] [CrossRef] [PubMed]
- Geerling, E.; Stone, E.T.; Steffen, T.L.; Hassert, M.; Brien, J.D.; Pinto, A.K. Obesity Enhances Disease Severity in Female Mice Following West Nile Virus Infection. Front. Immunol. 2021, 12, 739025. [Google Scholar] [CrossRef] [PubMed]
- Jean, C.M.; Honarmand, S.; Louie, J.K.; Glaser, C.A. Risk Factors for West Nile Virus Neuroinvasive Disease, California, 2005. Emerg. Infect. Dis. 2007, 13, 1918–1920. [Google Scholar] [CrossRef] [PubMed]
- Chowers, M.Y.; Lang, R.; Nassar, F.; Ben-David, D.; Giladi, M.; Rubinshtein, E.; Itzhaki, A.; Mishal, J.; Siegman-Igra, Y.; Kitzes, R.; et al. Clinical Characteristics of the West Nile Fever Outbreak, Israel, 2000. Emerg. Infect. Dis. 2001, 7, 675–678. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reslan, A.; Haddad, J.G.; Desprès, P.; Bascands, J.-L.; Gadea, G. High Glucose Induces in HK2 Kidney Cells an IFN–Dependent ZIKV Antiviral Status Fueled by Viperin. Biomedicines 2022, 10, 1577. https://doi.org/10.3390/biomedicines10071577
Reslan A, Haddad JG, Desprès P, Bascands J-L, Gadea G. High Glucose Induces in HK2 Kidney Cells an IFN–Dependent ZIKV Antiviral Status Fueled by Viperin. Biomedicines. 2022; 10(7):1577. https://doi.org/10.3390/biomedicines10071577
Chicago/Turabian StyleReslan, Alawiya, Juliano G. Haddad, Philippe Desprès, Jean-Loup Bascands, and Gilles Gadea. 2022. "High Glucose Induces in HK2 Kidney Cells an IFN–Dependent ZIKV Antiviral Status Fueled by Viperin" Biomedicines 10, no. 7: 1577. https://doi.org/10.3390/biomedicines10071577