
The Neurobiological Correlates of Gaze Perception in Healthy Individuals and Neurologic Patients

 ,
, 
Abstract
:1. Introduction
2. Neural Correlates of Gaze Direction: The Pivotal Role of the STS
3. Difference between Gaze Cues and Other Symbolic Cues
4. Conjoint Processing of Emotional Expressions and Gaze Direction in the Amygdala and STS?
4.1. The Time-Course of Processing of Emotional Expressions and Gaze Direction in the Amygdala and STS
4.2. Transient or Sustained Presentation of Visual Stimuli
4.3. Gaze Direction and Emotional Expression: Low-Level Visual Features
5. Subliminal vs. Supraliminal Cues
6. Brain-Damaged Patients and Transcranial Magnetic Stimulation (TMS) Evidence
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Argyle, M.; Cook, M. Gaze and Mutual Gaze; Cambridge U Press: Cambridge, UK, 1976. [Google Scholar]
- Emery, N.J. The Eyes Have It: The Neuroethology, Function and Evolution of Social Gaze. Neurosci. Biobehav. Rev. 2000, 24, 581–604. [Google Scholar] [CrossRef]
- Kleinke, C.L. Gaze and Eye Contact. A Research Review. Psychol. Bull. 1986, 100, 78–100. [Google Scholar] [CrossRef] [PubMed]
- Leopold, D.; Rhodes, G. A Comparative View of Face Perception. J. Comp. Psychol. 2010, 124, 233–251. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Song, H.; Guan, L.; Wu, X. The Neural Basis of Intergroup Threat Effect on Social Attention. Sci. Rep. 2017, 7, 41062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayliss, A.P.; Tipper, S.P. Predictive Gaze Cues and Personality Judgments: Should Eye Trust You? Psychol. Sci. 2006, 17, 514–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deaner, R.O.; Platt, M.L. Reflexive Social Attention in Monkeys and Humans. Curr. Biol. 2003, 13, 1609–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, T.; Sato, W.; Kubota, Y.; Usui, K.; Inoue, Y.; Murai, T.; Hayashi, T.; Toichi, M. Involvement of Medial Temporal Structures in Reflexive Attentional Shift by Gaze. Soc. Cogn. Affect. Neurosci. 2008, 3, 80–88. [Google Scholar] [CrossRef]
- Driver, J.; Davis, G.; Ricciardelli, P.; Kidd, P.; Maxwell, E.; Baron-Cohen, S. Gaze Perception Triggers Reflexive Visuospatial Orienting. Vis. Cogn. 1999, 6, 509–540. [Google Scholar] [CrossRef]
- Deaner, R.O.; Shepherd, S.V.; Platt, M.L. Familiarity Accentuates Gaze Cuing in Women but Not Men. Biol. Lett. 2007, 3, 64–67. [Google Scholar] [CrossRef] [Green Version]
- Langton, S.R.H.; Bruce, V. Reflexive Visual Orienting in Response to the Social Attention of Others. Vis. Cogn. 1999, 6, 541–567. [Google Scholar] [CrossRef]
- Frischen, A.; Bayliss, A.P.; Tipper, S.P. Gaze Cueing of Attention Visual Attention, Social Cognition, and Individual Differences. Psychol. Bull. 2007, 133, 694–724. [Google Scholar] [CrossRef] [PubMed]
- Hadders-Algra, M. Human Face and Gaze Perception Is Highly Context Specific and Involves Bottom-up and Top-down Neural Processing. Neurosci. Biobehav. Rev. 2022, 132, 304–323. [Google Scholar] [CrossRef] [PubMed]
- Friesen, C.K.; Kingstone, A. The Eyes Have It! Reflexive Orienting Is Triggered by Nonpredictive Gaze. Psychon. Bull. Rev. 1998, 5, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Dalmaso, M.; Castelli, L.; Galfano, G. Social Modulators of Gaze-Mediated Orienting of Attention: A Review Social Modulators of Gaze-Mediated Orienting of Attention: A Review. Psychon. Bull. Rev. 2020, 27, 833–855. [Google Scholar] [CrossRef]
- Mathews, A.; Fox, E.; Yiend, J.; Calder, A. The Face of Fear: Effects of Eye Gaze and Emotion on Visual Attention. Vis. Cogn. 2003, 10, 823–835. [Google Scholar] [CrossRef] [Green Version]
- Tipples, J. Fear and Fearfulness Potentiate Automatic Orienting to Eye Gaze. Cogn. Emot. 2006, 20, 309–320. [Google Scholar] [CrossRef]
- Pecchinenda, A.; Petrucci, M. Emotion Unchained: Facial Expression Modulates Gaze Cueing under Cognitive Load. PLoS ONE 2016, 11, e0168111. [Google Scholar] [CrossRef]
- McKay, K.; Grainger, S.; Coundouris, S.; Skorich, D.; Phillips, L.; Henry, J. Social Orienting of Attention: A Meta-Analytic Review of the Gaze-Cueing Effect. J. Vis. 2021, 21, 1977. [Google Scholar] [CrossRef]
- Jones, B.C.; DeBruine, L.M.; Main, J.C.; Little, A.C.; Welling, L.L.M.; Feinberg, D.R.; Tiddeman, B.R. Facial Cues of Dominance Modulate the Short-Term Gaze-Cuing Effect in Human Observers. Proc. Biol. Sci. 2010, 277, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Masulli, P.; Galazka, M.; Eberhard, D.; Gillberg, C.; Billstedt, E.; Hadjikhani, N.; Andersen, T.S. Data-Driven Analysis of Gaze Patterns in Face Perception: Methodological and Clinical Contributions. Sci. Direct 2021, 7, 9–23. [Google Scholar] [CrossRef]
- Adams, R.B.; Kleck, R.E. Effects of Direct and Averted Gaze on the Perception of Facially Communicated Emotion. Emotion 2005, 5, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebert, A.; Chaby, L.; Guillin, A.; Chekroun, S.; Vergilino-Perez, D. Are You “Gazing” at Me? How Others’ Gaze Direction and Facial Expression Influence Gaze Perception and Postural Control. Front. Psychol. 2021, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Narganes, C.; Ana, P.; Juan, B.C.; Andrea, L. Explicit vs. Implicit Spatial Processing in Arrow vs. Eye-Gaze Spatial Congruency Effects. Psychol. Res. 2022, 0123456789. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.A.; Haxby, J.V. Distinct Representations of Eye Gaze and Identity in the Distributed Human Neural System for Face Perception. Nat. Neurosci. 2000, 3, 80–84. [Google Scholar] [CrossRef]
- Adolphs, R.; Spezio, M. Role of the Amygdala in Processing Visual Social Stimuli. Prog. Brain Res. 2006, 156, 363–378. [Google Scholar] [CrossRef]
- Wang, S.; Yu, R.; Tyszka, J.M.; Zhen, S.; Kovach, C.; Sun, S.; Huang, Y.; Hurlemann, R.; Ross, I.B.; Chung, J.M.; et al. The Human Amygdala Parametrically Encodes the Intensity of Specific Facial Emotions and Their Categorical Ambiguity. Nat. Commun. 2017, 8, 14821. [Google Scholar] [CrossRef] [Green Version]
- Diano, M.; Tamietto, M.; Celeghin, A.; Weiskrantz, L.; Tatu, M.K.; Bagnis, A.; Duca, S.; Geminiani, G.; Cauda, F.; Costa, T. Dynamic Changes in Amygdala Psychophysiological Connectivity Reveal Distinct Neural Networks for Facial Expressions of Basic Emotions. Sci. Rep. 2017, 7, 45260. [Google Scholar] [CrossRef]
- Pelphrey, K.A.; Viola, R.J.; McCarthy, G. When Strangers Pass: Processing of Mutual and Averted Social Gaze in the Superior Temporal Sulcus. Psychol. Sci. 2004, 15, 598–603. [Google Scholar] [CrossRef]
- George, N.; Conty, L. Facing the Gaze of Others. Neurophysiol. Clin. 2008, 38, 197–207. [Google Scholar] [CrossRef]
- Akiyama, T.; Kato, M.; Muramatsu, T.; Umeda, S.; Saito, F.; Kashima, H. Unilateral Amygdala Lesions Hamper Attentional Orienting Triggered by Gaze Direction. Cereb. Cortex 2007, 17, 2593–2600. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.B.; Gordon, H.L.; Baird, A.A.; Ambady, N.; Kleck, R.E. Effects of Gaze on Amygdala Sensitivity to Anger and Fear Faces. Science 2003, 300, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjikhani, N.; Hoge, R.; Snyder, J.; de Gelder, B. Pointing with the Eyes: The Role of Gaze in Communicating Danger. Brain Cogn. 2008, 68, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardee, J.E.; Thompson, J.C.; Puce, A. The Left Amygdala Knows Fear: Laterality in the Amygdala Response to Fearful Eyes. Soc. Cogn. Affect. Neurosci. 2008, 3, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fichtenholtz, H.M.; Hopfinger, J.B.; Graham, R.; Detwiler, J.M.; LaBar, K.S. Event-Related Potentials Reveal Temporal Staging of Dynamic Facial Expression and Gaze Shift Effects on Attentional Orienting. Soc. Neurosci. 2009, 4, 317–331. [Google Scholar] [CrossRef]
- Kennedy, D.P.; Adolphs, R. The Social Brain in Psychiatric and Neurological Disorders. Trends Cogn. Sci. 2012, 16, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.L.; Wu, R.M. Social Brain Dysfunctions in Patients with Parkinson’s Disease: A Review of Theory of Mind Studies. Transl. Neurodegener. 2013, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, S.; Serio, G.; Scarpazza, C.; D’Ausilio, A.; Borgomaneri, S. Frozen in (e)Motion: How Reactive Motor Inhibition Is Influenced by the Emotional Content of Stimuli in Healthy and Psychiatric Populations. Behav. Res. Ther. 2021, 146, 103963. [Google Scholar] [CrossRef]
- Spekker, E.; Tanaka, M.; Szabó, Á.; Vécsei, L. Neurogenic Inflammation: The Participant in Migraine and Recent Advancements in Translational Research. Biomedicines 2022, 10, 76. [Google Scholar] [CrossRef]
- Tanaka, M.; Vécsei, L. Editorial of Special Issue “Crosstalk between Depression, Anxiety, and Dementia: Comorbidity in Behavioral Neurology and Neuropsychiatry”. Biomedicines 2021, 9, 517. [Google Scholar] [CrossRef]
- Mormann, F.; Niediek, J.; Tudusciuc, O.; Quesada, C.M.; Coenen, V.; Elger, C.; Adolphs, R. Neurons in the Human Amygdala Encode Face Identity but Not Gaze Direction. Nat. Neurosci. 2016, 18, 1568–1570. [Google Scholar] [CrossRef]
- Hooker, C.I.; Paller, K.A.; Gitelman, D.R.; Parrish, T.B.; Mesulam, M.-M.; Reber, P.J. Brain Networks for Analyzing Eye Gaze. Brain Res. Cogn. Brain Res. 2003, 17, 406–418. [Google Scholar] [CrossRef] [Green Version]
- Graham, R.; Labar, K.S. Face Processing and Social Attention. Neuropsychologia 2013, 50, 553–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itier, R.J.; Batty, M. Neural Bases of Eye and Gaze Processing: The Core of Social Cognition. Neurosci. Biobehav. Rev. 2009, 33, 843–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, N.; Driver, J.; Dolan, R.J. Seen Gaze-Direction Modulates Fusiform Activity and Its Coupling with Other Brain Areas during Face Processing. Neuroimage 2001, 13, 1102–1112. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, R.; Sugiura, M.; Kato, T.; Nakamura, A.; Hatano, K.; Ito, K.; Fukuda, H.; Kojima, S.; Nakamura, K. The Human Amygdala Plays an Important Role in Gaze Monitoring: A PET Study. Brain 1999, 122, 779–783. [Google Scholar] [CrossRef]
- Wicker, B.; Perrett, D.I.; Baron-Cohen, S.; Decety, J. Being the Target of Another’s Emotion: A PET Study. Neuropsychologia 2003, 41, 139–146. [Google Scholar] [CrossRef]
- Senju, A.; Johnson, M.H. The Eye Contact Effect: Mechanisms and Development. Trends Cogn. Sci. 2009, 13, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Pageler, N.M.; Menon, V.; Merin, N.M.; Eliez, S.; Brown, W.E.; Reiss, A.L. Effect of Head Orientation on Gaze Processing in Fusiform Gyrus and Superior Temporal Sulcus. Neuroimage 2003, 20, 318–329. [Google Scholar] [CrossRef]
- Sato, W.; Yoshikawa, S.; Kochiyama, T.; Matsumura, M. The Amygdala Processes the Emotional Significance of Facial Expressions: An FMRI Investigation Using the Interaction between Expression and Face Direction. Neuroimage 2004, 22, 1006–1013. [Google Scholar] [CrossRef]
- Pfeiffer, U.J.; Vogeley, K.; Schilbach, L. From Gaze Cueing to Dual Eye-Tracking: Novel Approaches to Investigate the Neural Correlates of Gaze in Social Interaction. Neurosci. Biobehav. Rev. 2013, 37, 2516–2528. [Google Scholar] [CrossRef]
- Haxby, J.V.; Hoffman, E.A.E.; Gobbini, M.M.I. The Distributed Human Neural System for Face Perception. Trends Cogn. Sci. 2000, 4, 223–233. [Google Scholar] [CrossRef]
- Nummenmaa, L.; Passamonti, L.; Rowe, J.; Engell, A.D.; Calder, A.J. Connectivity Analysis Reveals a Cortical Network for Eye Gaze Perception. Cereb. Cortex 2010, 20, 1780–1787. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Byrne, P.; Yan, X.; Henriques, D.Y.P.; Crawford, J.D. Allocentric versus Egocentric Representation of Remembered Reach Targets in Human Cortex. J. Neurosci. 2014, 34, 12515–12526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanwisher, N.; McDermott, J.; Chun, M.M. The Fusiform Face Area: A Module in Human Extrastriate Cortex Specialized for Face Perception. J. Neurosci. 1997, 17, 4302–4311. [Google Scholar] [CrossRef] [PubMed]
- Calder, A.J.; Beaver, J.D.; Winston, J.S.; Dolan, R.J.; Jenkins, R.; Eger, E.; Henson, R.N.A.A. Separate Coding of Different Gaze Directions in the Superior Temporal Sulcus and Inferior Parietal Lobule. Curr. Biol. 2007, 17, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelphrey, K.A.; Singerman, J.D.; Allison, T.; McCarthy, G. Brain Activation Evoked by Perception of Gaze Shifts: The Influence of Context. Neuropsychologia 2003, 41, 156–170. [Google Scholar] [CrossRef]
- Puce, A.; Allison, T.; Bentin, S.; Gore, J.C.; Mccarthy, G. Temporal Cortex Activation in Humans Viewing Eye and Mouth Movements. J. Neurosci. 1998, 18, 2188–2199. [Google Scholar] [CrossRef] [Green Version]
- Mcgettigan, C.; Jasmin, K.; Eisner, F.; Agnew, Z.K.; Josephs, O.J.; Calder, A.J.; Jessop, R.; Lawson, R.P.; Spielmann, M.; Scott, S.K. Neuropsychologia You Talkin’ to Me? Communicative Talker Gaze Activates Left-Lateralized Superior Temporal Cortex during Perception of Degraded Speech. Neuropsychologia 2017, 100, 51–63. [Google Scholar] [CrossRef]
- Carlin, J.D.; Calder, A.J.; Kriegeskorte, N.; Nili, H.; Rowe, J.B. A Head View-Invariant Representation of Gaze Direction in Anterior Superior Temporal Sulcus. Curr. Biol. 2011, 21, 1817–1821. [Google Scholar] [CrossRef] [Green Version]
- Carlin, J.D.; Rowe, J.B.; Kriegeskorte, N.; Thompson, R.; Calder, A.J. Direction-Sensitive Codes for Observed Head Turns in Human Superior Temporal Sulcus. Cereb. Cortex 2012, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Lockhofen, D.E.L.; Gruppe, H.; Ruprecht, C.; Gallhofer, B.; Sammer, G. Hemodynamic Response Pattern of Spatial Cueing Is Different for Social and Symbolic Cues. Front. Hum. Neurosci. 2014, 8, 912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingstone, A.; Tipper, C.; Ristic, J.; Ngan, E. The Eyes Have It!: An FMRI Investigation. Brain Cogn. 2004, 55, 269–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Materna, S.; Dicke, P.W.; Thier, P. Dissociable Roles of the Superior Temporal Sulcus and the Intraparietal Sulcus in Joint Attention: A Functional Magnetic Resonance Imaging Study. J. Cogn. Neurosci. 2008, 20, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Sato, W.; Kochiyama, T.; Uono, S.; Yoshikawa, S. Amygdala Integrates Emotional Expression and Gaze Direction in Response to Dynamic Facial Expressions. Neuroimage 2010, 50, 1658–1665. [Google Scholar] [CrossRef] [Green Version]
- Callejas, A.; Shulman, G.L.; Corbetta, M. Dorsal and Ventral Attention Systems Underlie Social and Symbolic Cueing. J. Cogn. Neurosci. 2014, 26, 63–80. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.J.; Mooshagian, E.; Kaplan, J.T.; Zaidel, E.; Iacoboni, M. The Neural Correlates of Social Attention: Automatic Orienting to Social and Nonsocial Cues. Psychol. Res. 2009, 73, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Engell, A.D.; Nummenmaa, L.; Oosterhof, N.N.; Henson, R.N.; Haxby, J.V.; Calder, A.J. Differential Activation of Frontoparietal Attention Networks by Social and Symbolic Spatial Cues. Soc. Cogn. Affect. Neurosci. 2010, 5, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Hietanen, J.K.; Nummenmaa, L.; Nyman, M.J.; Parkkola, R.; Hämäläinen, H. Automatic Attention Orienting by Social and Symbolic Cues Activates Different Neural Networks: An FMRI Study. Neuroimage 2006, 33, 406–413. [Google Scholar] [CrossRef]
- Joseph, R.M.; Fricker, Z.; Keehn, B. Activation of Frontoparietal Attention Networks by Non-Predictive Gaze and Arrow Cues. Soc. Cogn. Affect. Neurosci. 2015, 10, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Uono, S.; Sato, W.; Kochiyama, T. Commonalities and Differences in the Spatiotemporal Neural Dynamics Associated with Automatic Attentional Shifts Induced by Gaze and Arrows. Neurosci. Res. 2014, 87, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, C.; Uono, S.; Yoshimura, S.; Toichi, M. Human Cortical Activity Evoked by Contextual Processing in Attentional Orienting. Sci. Rep. 2017, 7, 2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelphrey, K.A.; Morris, J.P.; McCarthy, G. Grasping the Intentions of Others: The Perceived Intentionality of an Action Influences Activity in the Superior Temporal Sulcus during Social Perception. J. Cogn. Neurosci. 2004, 16, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, K.; Ramezanpour, H.; Dicke, P.W.; Thier, P. Following Eye Gaze Activates a Patch in the Posterior Temporal Cortex That Is Not Part of the Human “Face Patch” System. eNeuro 2017, 4, ENEURO.0317-16.2017. [Google Scholar] [CrossRef] [Green Version]
- Sato, W.; Kochiyama, T.; Uono, S.; Yoshikawa, S. Commonalities in the Neural Mechanisms Underlying Automatic Attentional Shifts by Gaze, Gestures, and Symbols. Neuroimage 2009, 45, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Ristic, J.; Friesen, C.K.; Kingstone, A. Are Eyes Special? It Depends on How You Look at It. Psychon. Bull. Rev. 2002, 9, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Tipples, J. Eye Gaze Is Not Unique: Automatic Orienting in Response to Uninformative Arrows. Psychon. Bull. Rev. 2002, 9, 314–318. [Google Scholar] [CrossRef] [Green Version]
- Tipper, C.M.; Handy, T.C.; Giesbrecht, B.; Kingstone, A. Brain Responses to Biological Relevance. J. Cogn. Neurosci. 2008, 20, 879–891. [Google Scholar] [CrossRef]
- N’Diaye, K.; Sander, D.; Vuilleumier, P. Self-Relevance Processing in the Human Amygdala: Gaze Direction, Facial Expression, and Emotion Intensity. Emotion 2009, 9, 798–806. [Google Scholar] [CrossRef]
- Krämer, K.; Bente, G.; Kuzmanovic, B.; Barisic, I.; Pfeiffer, U.J.; Georgescu, A.L.; Vogeley, K. Neural Correlates of Emotion Perception Depending on Culture and Gaze Direction. Cult. Brain 2014, 2, 27–51. [Google Scholar] [CrossRef] [Green Version]
- Engell, A.D.; Haxby, J.V. Facial Expression and Gaze-Direction in Human Superior Temporal Sulcus. Neuropsychologia 2007, 45, 3234–3241. [Google Scholar] [CrossRef]
- Ewbank, M.P.; Fox, E.; Calder, A.J. The Interaction between Gaze and Facial Expression in the Amygdala and Extended Amygdala Is Modulated by Anxiety. Front. Hum. Neurosci. 2010, 4, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straube, T.; Langohr, B.; Schmidt, S.; Mentzel, H.J.; Miltner, W.H.R. Increased Amygdala Activation to Averted versus Direct Gaze in Humans Is Independent of Valence of Facial Expression. Neuroimage 2010, 49, 2680–2686. [Google Scholar] [CrossRef] [PubMed]
- Ziaei, M.; Ebner, N.C.; Burianová, H. Functional Brain Networks Involved in Gaze and Emotional Processing. Eur. J. Neurosci. 2017, 45, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Schobert, A.-K.; Corradi-Dell’Acqua, C.; Frühholz, S.; van der Zwaag, W.; Vuilleumier, P. Functional Organization of Face Processing in the Human Superior Temporal Sulcus: A 7T High-Resolution FMRI Study. Soc. Cogn. Affect. Neurosci. 2018, 13, 102–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kätsyri, J.; de Gelder, B.; de Borst, A.W. Amygdala Responds to Direct Gaze in Real but Not in Computer-Generated Faces. Neuroimage 2020, 204, 116216. [Google Scholar] [CrossRef]
- Allison, T.; Puce, A.; McCarthy, G. Social Perception from Visual Cues: Role of the STS Region. Trends Cogn. Sci. 2000, 4, 267–278. [Google Scholar] [CrossRef]
- Campbell, R.; Heywood, C.A.; Cowey, A.; Regard, M.; Landis, T. Sensitivity to Eye Gaze in Prosopagnosic Patients and Monkeys with Superior Temporal Sulcus Ablation. Neuropsychologia 1990, 28, 1123–1142. [Google Scholar] [CrossRef]
- Boll, S.; Gamer, M.; Kalisch, R.; Büchel, C. Processing of Facial Expressions and Their Significance for the Observer in Subregions of the Human Amygdala. Neuroimage 2011, 56, 299–306. [Google Scholar] [CrossRef]
- Balogh, L.; Tanaka, M.; Török, N.; Taguchi, S. Crosstalk between Existential Phenomenological Psychotherapy and Neurological Sciences in Mood and Anxiety Disorders. Biomedicines 2021, 9, 340. [Google Scholar] [CrossRef]
- Tanaka, M.; Török, N.; Fanni, T.; Szab, Á. Co-Players in Chronic Pain: Neuroinflammation and the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 897. [Google Scholar] [CrossRef]
- Tanaka, M.; Fanni, T.; Poly, H.; Szab, Á.; Yvette, M. Immune Influencers in Action: Metabolites and Enzymes of the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Keil, M.F.; Briassoulis, G.; Stratakis, C.A.; Wu, T.J. Protein Kinase A and Anxiety-Related Behaviors: A Mini-Review. Front. Endocrinol. 2016, 7, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, D.; Grafman, J.; Zalla, T. The Human Amygdala: An Evolved System for Relevance Detection. Rev. Neurosci. 2003, 14, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Talipski, L.A.; Bell, E.; Goodhew, S.C.; Dawel, A.; Edwards, M. Examining the Effects of Social Anxiety and Other Individual Differences on Gaze-Directed Attentional Shifts. Q. J. Exp. Psychol. 2021, 74, 771–785. [Google Scholar] [CrossRef]
- Hoffman, K.L.; Gothard, K.M.; Schmid, M.C.; Logothetis, N.K. Facial-Expression and Gaze-Selective Responses in the Monkey Amygdala. Curr. Biol. 2007, 17, 766–772. [Google Scholar] [CrossRef] [Green Version]
- Cushing, C.A.; Im, H.Y.; Adams, R.B.; Ward, N.; Albohn, D.N.; Steiner, T.G.; Kveraga, K. Neurodynamics and Connectivity during Facial Fear Perception: The Role of Threat Exposure and Signal Congruity. Sci. Rep. 2018, 8, 2776. [Google Scholar] [CrossRef] [Green Version]
- Lachat, F.; Farroni, T.; George, N. Watch out! Magnetoencephalographic Evidence for Early Modulation of Attention Orienting by Fearful Gaze Cueing. PLoS ONE 2012, 7, e50499. [Google Scholar] [CrossRef]
- Pourtois, G.; Thut, G.; Grave de Peralta, R.; Michel, C.; Vuilleumier, P. Two Electrophysiological Stages of Spatial Orienting towards Fearful Faces: Early Temporo-Parietal Activation Preceding Gain Control in Extrastriate Visual Cortex. Neuroimage 2005, 26, 149–163. [Google Scholar] [CrossRef]
- Righart, R.; de Gelder, B. Context Influences Early Perceptual Analysis of Faces-an Electrophysiological Study. Cereb. Cortex 2006, 16, 1249–1257. [Google Scholar] [CrossRef]
- Williams, L.M.; Palmer, D.; Liddell, B.J.; Song, L.; Gordon, E. The “when” and “Where” of Perceiving Signals of Threat versus Non-Threat. Neuroimage 2006, 31, 458–467. [Google Scholar] [CrossRef]
- Borhani, K.; Borgomaneri, S.; Làdavas, E.; Bertini, C. The Effect of Alexithymia on Early Visual Processing of Emotional Body Postures. Biol. Psychol. 2016, 115, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dumas, T.; Dubal, S.; Attal, Y.; Chupin, M.; Jouvent, R.; Morel, S.; George, N. MEG Evidence for Dynamic Amygdala Modulations by Gaze and Facial Emotions. PLoS ONE 2013, 8, e74145. [Google Scholar] [CrossRef]
- Conty, L.; Dezecache, G.; Hugueville, L.; Grèzes, J. Early Binding of Gaze, Gesture, and Emotion: Neural Time Course and Correlates. J. Neurosci. 2012, 32, 4531–4539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuilleumier, P.; Armony, J.L.; Driver, J.; Dolan, R.J. Distinct Spatial Frequency Sensitivities for Processing Faces and Emotional Expressions. Nat. Neurosci. 2003, 6, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.B.; Franklin, R.G.; Kveraga, K.; Ambady, N.; Kleck, R.E.; Whalen, P.J.; Hadjikhani, N.; Nelson, A.J. Amygdala Responses to Averted vs Direct Gaze Fear Vary as a Function of Presentation Speed. Soc. Cogn. Affect. Neurosci. 2012, 7, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Im, H.Y.; Adams, R.B.; Cushing, C.A.; Boshyan, J.; Ward, N.; Kveraga, K. Sex-Related Differences in Behavioral and Amygdalar Responses to Compound Facial Threat Cues. Hum. Brain Mapp. 2018, 39, 2725–2741. [Google Scholar] [CrossRef] [Green Version]
- Lahnakoski, J.M.; Glerean, E.; Salmi, J.; Jääskeläinen, I.P.; Sams, M.; Hari, R.; Nummenmaa, L. Naturalistic FMRI Mapping Reveals Superior Temporal Sulcus as the Hub for the Distributed Brain Network for Social Perception. Front. Hum. Neurosci. 2012, 6, 233. [Google Scholar] [CrossRef] [Green Version]
- Adolphs, R. The Social Brain: Neural Basis of Social Knowledge. Annu. Rev. Psychol. 2009, 60, 693–716. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.B.; Rule, N.O.; Franklin, R.G.; Wang, E.; Stevenson, M.T.; Yoshikawa, S.; Nomura, M.; Sato, W.; Kveraga, K.; Ambady, N. Cross-Cultural Reading the Mind in the Eyes: An FMRI Investigation. J. Cogn. Neurosci. 2010, 22, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Sauer, A.; Mothes-Lasch, M.; Miltner, W.H.R.; Straube, T. Effects of Gaze Direction, Head Orientation and Valence of Facial Expression on Amygdala Activity. Soc. Cogn. Affect. Neurosci. 2014, 9, 1246–1252. [Google Scholar] [CrossRef] [Green Version]
- Gamer, M.; Büchel, C. Amygdala Activation Predicts Gaze toward Fearful Eyes. J. Neurosci. 2009, 29, 9123–9126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsuda, T.; Masaki, S. Subliminal Gaze Cues Increase Preference Levels for Items in the Gaze Direction. Cogn. Emot. 2018, 32, 1146–1151. [Google Scholar] [CrossRef] [PubMed]
- Sato, W.; Okada, T.; Toichi, M. Attentional Shift by Gaze Is Triggered without Awareness. Exp. Brain Res. 2007, 183, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, S.; Miyatani, M. The Effects of Facial Expressions on Gaze-Triggered Attention without Awareness. Jpn. J. Cogn. Psychol. 2010, 8, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Zhang, S.; Geng, H. Gaze-Induced Joint Attention Persists under High Perceptual Load and Does Not Depend on Awareness. Vis. Res. 2011, 51, 2048–2056. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Zhang, S.; Tao, R.; Geng, H. The Power of Subliminal and Supraliminal Eye Contact on Social Decision Making: An Individual-Difference Perspective. Conscious. Cogn. 2016, 40, 131–140. [Google Scholar] [CrossRef]
- Schütz, C.; Güldenpenning, I.; Koester, D.; Schack, T. Social Cues Can Impact Complex Behavior Unconsciously. Sci. Rep. 2020, 10, 21017. [Google Scholar] [CrossRef]
- Sato, W.; Kochiyama, T.; Uono, S.; Toichi, M. Neural Mechanisms Underlying Conscious and Unconscious Attentional Shifts Triggered by Eye Gaze. Neuroimage 2016, 124, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Bailey, P.E.; Slessor, G.; Rendell, P.G.; Bennetts, R.J.; Campbell, A.; Ruffman, T. Age Differences in Conscious versus Subconscious Social Perception: The Influence of Face Age and Valence on Gaze Following. Psychol. Aging 2014, 29, 491–502. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Serio, G.; Battaglia, S. Please, Don’t Do It! Fifteen Years of Progress of Non-Invasive Brain Stimulation in Action Inhibition. Cortex 2020, 132, 404–422. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Lu, M.K.; Antal, A.; Classen, J.; Nitsche, M.; Ziemann, U.; Ridding, M.; Hamada, M.; Ugawa, Y.; Jaberzadeh, S.; et al. Plasticity Induced by Non-Invasive Transcranial Brain Stimulation: A Position Paper. Clin. Neurophysiol. 2017, 128, 2318–2329. [Google Scholar] [CrossRef] [PubMed]
- Vosskuhl, J.; Strüber, D.; Herrmann, C.S. Non-Invasive Brain Stimulation: A Paradigm Shift in Understanding Brain Oscillations. Front. Hum. Neurosci. 2018, 12, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miniussi, C.; Harris, J.A.; Ruzzoli, M. Modelling Non-Invasive Brain Stimulation in Cognitive Neuroscience. Neurosci. Biobehav. Rev. 2013, 37, 1702–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xuan, R.; Chen, Q.; Zhao, Q.; Shi, Z.; Du, J.; Zhu, C.; Yu, F.; Ji, G.; Wang, K. High-definition transcranial direct current stimulation modulates eye gaze on emotional faces in college students with alexithymia: An eye-tracking study. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 116, 110521. [Google Scholar] [CrossRef]
- Betti, S.; Zani, G.; Granziol, U.; Guerra, S.; Castiello, U.; Sartori, L.; Shepherd, S.V. Look at Me: Early Gaze Engagement Enhances Corticospinal Excitability During Action Observation. Front. Psychol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Ito, T.; Kamiue, M.; Kihara, T.; Ishimaru, Y.; Kimura, D.; Tsubahara, A. Visual Attention and Motion Visibility Modulate Motor Resonance during Observation of Human Walking in Different Manners. Brain Sci. 2021, 11, 679. [Google Scholar] [CrossRef]
- Akiyama, T.; Kato, M.; Muramatsu, T.; Saito, F.; Nakachi, R.; Kashima, H. A Deficit in Discriminating Gaze Direction in a Case with Right Superior Temporal Gyrus Lesion. Neuropsychologia 2006, 44, 161–170. [Google Scholar] [CrossRef]
- Akiyama, T.; Kato, M.; Muramatsu, T.; Saito, F.; Umeda, S.; Kashima, H. Gaze but Not Arrows: A Dissociative Impairment after Right Superior Temporal Gyrus Damage. Neuropsychologia 2006, 44, 1804–1810. [Google Scholar] [CrossRef]
- Roy, A.; Shepherd, S.V.; Platt, M.L. Reversible Inactivation of PSTS Suppresses Social Gaze Following in the Macaque (Macaca Mulatta). Soc. Cogn. Affect. Neurosci. 2014, 9, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Young, A.W.; Aggleton, J.P.; Hellawell, D.J.; Johnson, M.; Broks, P.; Hanley, J.R. Face Processing Impairments after Amygdalotomy. Brain 1995, 118, 15–24. [Google Scholar] [CrossRef]
- Cristinzio, C.; N’Diaye, K.; Seeck, M.; Vuilleumier, P.; Sander, D. Integration of Gaze Direction and Facial Expression in Patients with Unilateral Amygdala Damage. Brain 2010, 133, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Spezio, M.L.; Huang, P.Y.S.; Castelli, F.; Adolphs, R. Amygdala Damage Impairs Eye Contact during Conversations with Real People. J. Neurosci. 2007, 27, 3994–3997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sliwinska, M.W.; Pitcher, D. TMS Demonstrates That Both Right and Left Superior Temporal Sulci Are Important for Facial Expression Recognition. Neuroimage 2018, 183, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paracampo, R.; Pirruccio, M.; Costa, M.; Borgomaneri, S.; Avenanti, A. Visual, Sensorimotor and Cognitive Routes to Understanding Others’ Enjoyment: An Individual Differences RTMS Approach to Empathic Accuracy. Neuropsychologia 2018, 116, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D. Facial Expression Recognition Takes Longer in the Posterior Superior Temporal Sulcus than in the Occipital Face Area. J. Neurosci. 2014, 34, 9173–9177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourtois, G.; Sander, D.; Andres, M.; Grandjean, D.; Reveret, L.; Olivier, E.; Vuilleumier, P. Dissociable Roles of the Human Somatosensory and Superior Temporal Cortices for Processing Social Face Signals. Eur. J. Neurosci. 2004, 20, 3507–3515. [Google Scholar] [CrossRef] [PubMed]
- Saitovitch, A.; Popa, T.; Lemaitre, H.; Rechtman, E.; Lamy, J.-C.; Grévent, D.; Calmon, R.; Meunier, S.; Brunelle, F.; Samson, Y.; et al. Tuning Eye-Gaze Perception by Transitory STS Inhibition. Cereb. Cortex 2016, 26, 2823–2831. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, H.; Kaufman, O.; Damasio, H.; Damasio, A.R.; Granner, M.; Bakken, H.; Hori, T.; Howard, M.A.; Adolphs, R. Single-Neuron Responses to Emotional Visual Stimuli Recorded in Human Ventral Prefrontal Cortex. Nat. Neurosci. 2001, 4, 15–16. [Google Scholar] [CrossRef]
- Eimer, M.; Holmes, A. An ERP Study on the Time Course of Emotional Face Processing. Neuroreport 2002, 13, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Eimer, M.; Kiss, M.; Holmes, A. Links between Rapid ERP Responses to Fearful Faces and Conscious Awareness. J. Neuropsychol. 2008, 2, 165–181. [Google Scholar] [CrossRef]
- Ellena, G.; Battaglia, S.; Làdavas, E. The Spatial Effect of Fearful Faces in the Autonomic Response. Exp. Brain Res. 2020, 238, 2009–2018. [Google Scholar] [CrossRef] [PubMed]
- Öhman, A.; Soares, J.J. “Unconscious Anxiety”: Phobic Responses to Masked Stimuli. J. Abnorm. Psychol. 1994, 103, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Globisch, J.; Hamm, A.O.; Esteves, F.; Öhman, A. Fear Appears Fast: Temporal Course of Startle Reflex Potentiation in Animal Fearful Subjects. Psychophysiology 1999, 36, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Candini, M.; Battaglia, S.; Benassi, M.; Pellegrino, G.; Frassinetti, F. The Physiological Correlates of Interpersonal Space. Sci. Rep. 2021, 11, 2611. [Google Scholar] [CrossRef]
- Öhman, A.; Lundqvist, D.; Esteves, F. The Face in the Crowd Revisited: A Threat Advantage with Schematic Stimuli. J. Pers. Soc. Psychol. 2001, 80, 381–396. [Google Scholar] [CrossRef]
- Holmes, A.; Vuilleumier, P.; Eimer, M. The Processing of Emotional Facial Expression Is Gated by Spatial Attention: Evidence from Event-Related Brain Potentials. Brain Res. Cogn. Brain Res. 2003, 16, 174–184. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Avenanti, A. Early Changes in Corticospinal Excitability When Seeing Fearful Body Expressions. Sci. Rep. 2015, 5, 14122. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Gazzola, V.; Avenanti, A. Seeing Fearful Body Language Rapidly Freezes the Observer’s Motor Cortex. Cortex 2015, 65, 232–245. [Google Scholar] [CrossRef]
- Vicario, C.M.; Rafal, R.D.; Borgomaneri, S.; Paracampo, R.; Kritikos, A.; Avenanti, A. Pictures of Disgusting Foods and Disgusted Facial Expressions Suppress the Tongue Motor Cortex. Soc. Cogn. Affect. Neurosci. 2017, 12, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Borgomaneri, S.; Vitale, F.; Avenanti, A. Behavioral Inhibition System Sensitivity Enhances Motor Cortex Suppression When Watching Fearful Body Expressions. Brain Struct. Funct. 2017, 222, 3267–3282. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Vitale, F.; Avenanti, A. Early Motor Reactivity to Observed Human Body Postures Is Affected by Body Expression, Not Gender. Neuropsychologia 2020, 146, 107541. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Gazzola, V.; Avenanti, A. Temporal Dynamics of Motor Cortex Excitability during Perception of Natural Emotional Scenes. Soc. Cogn. Affect. Neurosci. 2014, 9, 1451–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgomaneri, S.; Vitale, F.; Battaglia, S.; Avenanti, A. Early Right Motor Cortex Response to Happy and Fearful Facial Expressions: A TMS Motor-Evoked Potential Study. Brain Sci. 2021, 11, 1203. [Google Scholar] [CrossRef] [PubMed]
- Nummenmaa, L.; Calder, A.J. Neural Mechanisms of Social Attention. Trends Cogn. Sci. 2009, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Carlin, J.D.; Calder, A.J. The Neural Basis of Eye Gaze Processing. Curr. Opin. Neurobiol. 2013, 23, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]



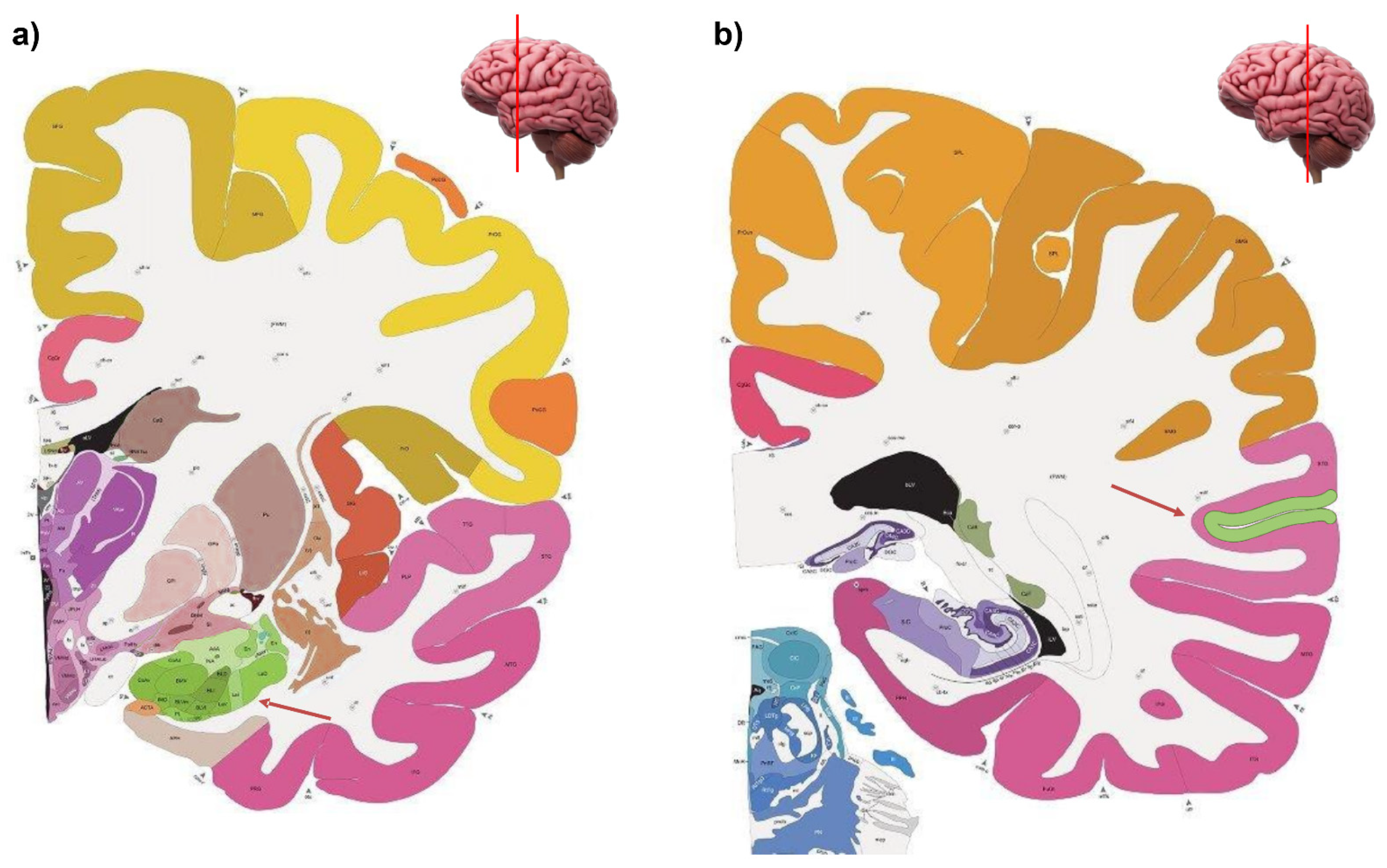{kind=link}
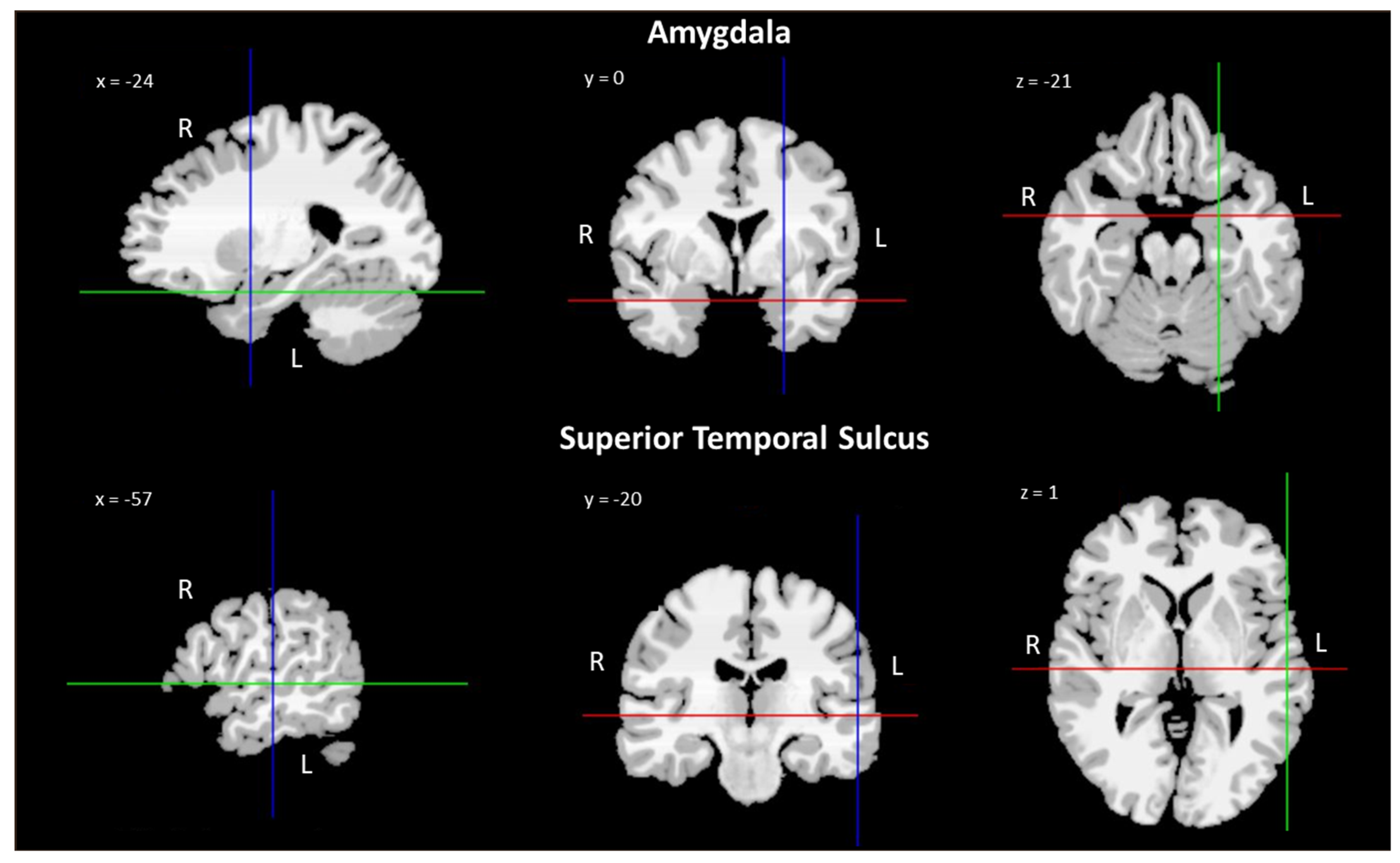{kind=link}
| Study | Emotional Facial Expression | fMRI Contrast | Paradigm | Coordinates (MNI) | Main Findings |
|---|---|---|---|---|---|
| Hooker et al. (2003) | Happy–angry | Angry > happy | Gaze-cueing | Right STS: 42 −57 26 Left STS: −35 −67 31 | STS for angry faces > happy faces |
| Adams et al. (2003) | Fear–angry | Angry averted/fearful direct > angry direct/fearful averted | Passive observation | Left amygdala: −15 0 −18 | Amygdala > angry averted and fearful directed gaze |
| Sato et al. (2004) | Angry–neutral | Expression × face direction | Gender task | Left amygdala: −22 −9 −16 | Amygdala > angry expressions looking toward |
| Engell and Haxby (2007) | Neutral direct–averted gaze anger, disgust, fear and surprise with direct gaze | Emotions > neutral Averted > direct | Match identity task | Right STS (emotion): 52 −48 8 Right STS (gaze): 36 −54 15 | STS emotions > neutral faces STS averted > direct gaze faces |
| Hadjikhani et al. (2008) | Fear | Fearful averted > fearful direct | Passive observation | Right STS (gaze): 48 −54 14 Amygdala (emotion): −12 −11 26 | Amygdala and the STS fearful averted gaze > fearful direct gaze |
| N’Diaye et al. (2009) | Fear–angry (intensity morphing) | Emotions > neutral Averted > direct | Emotional intensity rating | Left amygdala (emotion): −24 +3 −18 Right amygdala (emotion): +24 +6 −15 Left pSTS (gaze): −57 −42 +9 Right pSTS (gaze): +45 −66 +12 | Amygdala fearful averted gaze > directed gaze Amygdala angry directed gaze > averted gaze STS averted gaze > direct gaze |
| Ewbank et al. (2010) | Fear–angry–neutral | Angry direct > neutral direct Angry direct > angry averted Fearful averted > neutral averted | Gender task | Right amygdala (gaze): 22 −6 −12 Right amygdala (emotion × gaze): 24 −4 −12 | Amygdala high-anxious angry direct gaze > angry averted gaze Highly anxious > fearful direct gaze and averted gaze |
| Straube et al. (2010) | Happy–angry–neutral | Averted > direct Valence × gaze | Gender task | Left amygdala (gaze): −27 −9 −14 Right amygdala (gaze): 16 −6 −11 Right STS (valence × gaze): 54 −49 16 | Amygdala averted gaze > direct gaze STS averted emotional > averted neutral |
| Sato et al. (2010) | Happy–angry (static and dynamic) | Dynamic > static Direct > averted | Gender task | Left amygdala: −18 −10 −10 Right STS: 58 −46 8 | Amygdala dynamic happy and angry directed gaze > averted gaze STS dynamic > static |
| Krämer et al. (2014) | Happy–angry | European angry averted > Asian angry averted | Valence rating | Right amygdala: 28 −8 16 | Amygdala > angry averted gaze in ingroup |
| Ziaei et al. (2017) | Happy–angry–neutral | Expression with averted > all other conditions | Emotion recognition task | Left STS: 56 6 2 | Stronger connectivity of the STS and mPFC with bilateral amygdala for angry averted gaze > direct gaze |
| Schobert et al. (2018) | Happy–angry (gaze shift and speech motion) | Emotional (happy and angry) > non-emotional (gaze and speech) Eyes (angry and gaze) > mouth (happy and speech) | Identity detection task | Left pSTS: −50 −48 15 Right pSTS: 50 −47 13 | pSTS gaze-happy and gaze-angry > speech-happy and speech-angry |
| Kätsyri et al. (2020) | Angry–fear–neutral | Directed > averted + angry > fear | Circle detection in catch trials | pSTS: 65 −54 3 | Amygdala direct gaze > averted gaze Amygdala threatening > neutral STS gaze direction × emotional expression |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battaglia, S.; Fabius, J.H.; Moravkova, K.; Fracasso, A.; Borgomaneri, S. The Neurobiological Correlates of Gaze Perception in Healthy Individuals and Neurologic Patients. Biomedicines 2022, 10, 627. https://doi.org/10.3390/biomedicines10030627
Battaglia S, Fabius JH, Moravkova K, Fracasso A, Borgomaneri S. The Neurobiological Correlates of Gaze Perception in Healthy Individuals and Neurologic Patients. Biomedicines. 2022; 10(3):627. https://doi.org/10.3390/biomedicines10030627
Chicago/Turabian StyleBattaglia, Simone, Jasper H. Fabius, Katarina Moravkova, Alessio Fracasso, and Sara Borgomaneri. 2022. "The Neurobiological Correlates of Gaze Perception in Healthy Individuals and Neurologic Patients" Biomedicines 10, no. 3: 627. https://doi.org/10.3390/biomedicines10030627