
Gastrointestinal Microbiome and Neurologic Injury


Abstract
:1. Introduction
2. Neurologic Control of the Gut Microbiome
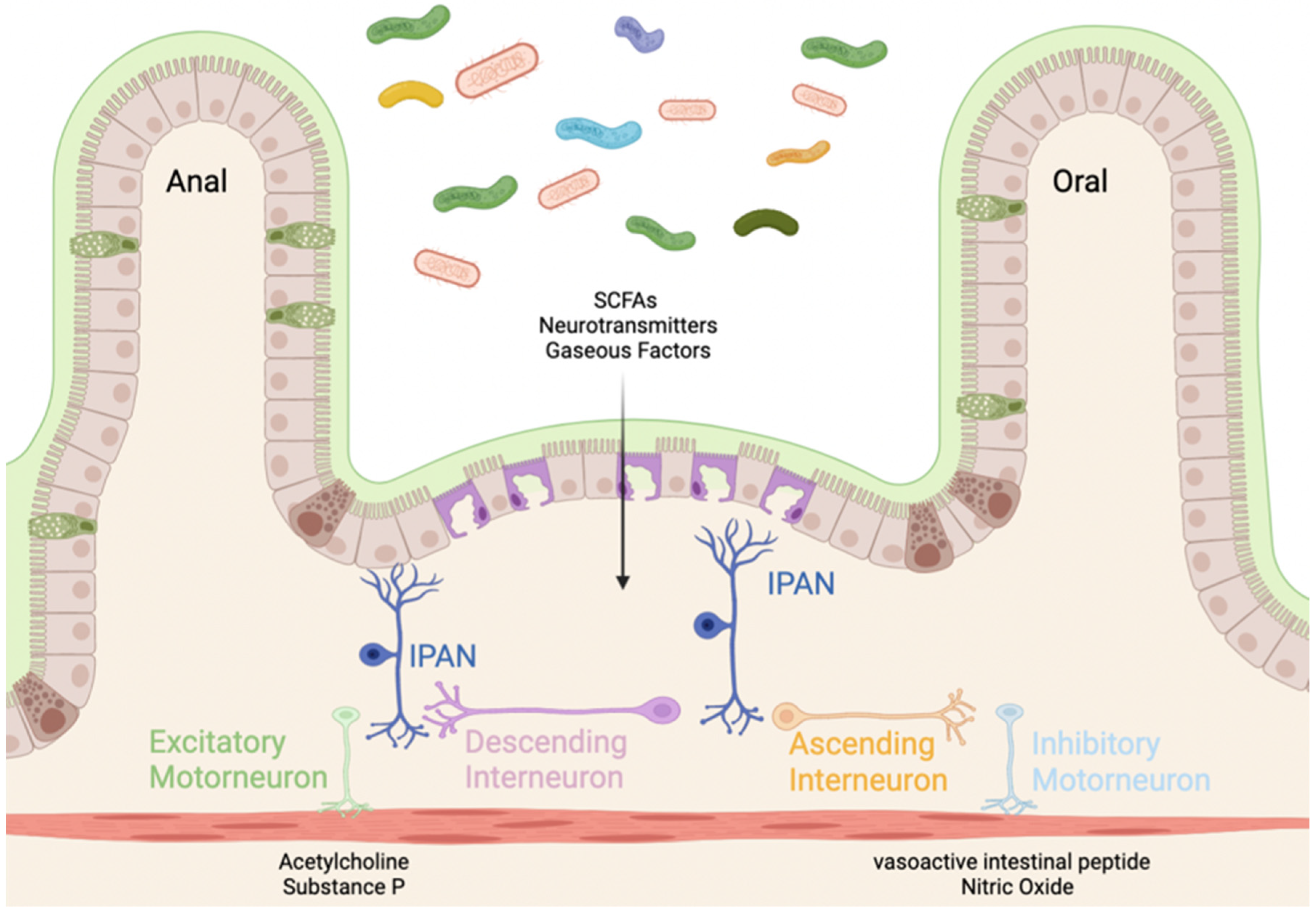
2.1. Intrinsic (Enteric) Nervous System
2.2. Microbiome Effect on Enteric Nervous System
2.3. Extrinsic Innervation of Gut
3. Mechanisms of Microbiome Disruption in Neurologic Disease
3.1. Ischemic Stroke
3.2. Spinal Cord Injury
3.3. Traumatic Brain Injury
3.4. Hemorrhagic Cerebrovascular Lesions
4. Interventions
4.1. Probiotics/Prebiotics
4.2. Fecal Microbiota Transplantation
4.3. Oral Short-Chain Fatty Acids (SCFAs)
4.4. Other Novel Interventions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The Enteric Nervous System and Gastrointestinal Innervation: Integrated Local and Central Control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Lyte, M., Cryan, J.F., Eds.; Springer: New York, NY, USA, 2014; pp. 39–71. ISBN 978-1-4939-0897-4. [Google Scholar]
- Browning, K.N.; Travagli, R.A. Central Nervous System Control of Gastrointestinal Motility and Secretion and Modulation of Gastrointestinal Functions. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2014; pp. 1339–1368. ISBN 978-0-470-65071-4. [Google Scholar]
- Chang, H.Y.; Mashimo, H.; Goyal, R.K., IV. Current Concepts of Vagal Efferent Projections to the Gut. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 284, G357–G366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, W.B. Oesophageal Peristalsis after Bilateral Vagotomy. Am. J. Physiol.-Leg. Content 1907, 19, 436–444. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System and Neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R.; Blackshaw, L.A.; Brookes, S.J.H.; Grundy, D. Neuroanatomy of Extrinsic Afferents Supplying the Gastrointestinal Tract. Neurogastroenterol. Motil. 2004, 16, 28–33. [Google Scholar] [CrossRef]
- Le Roux, C.W.; Neary, N.M.; Halsey, T.J.; Small, C.J.; Martinez-Isla, A.M.; Ghatei, M.A.; Theodorou, N.A.; Bloom, S.R. Ghrelin Does Not Stimulate Food Intake in Patients with Surgical Procedures Involving Vagotomy. J. Clin. Endocrinol. Metab. 2005, 90, 4521–4524. [Google Scholar] [CrossRef] [Green Version]
- Jänig, W. 3 Integration of Gut Function by Sympathetic Reflexes. Baillière’s Clin. Gastroenterol. 1988, 2, 45–62. [Google Scholar] [CrossRef]
- Garry, R.C. The Nervous Control of the Caudal Region of the Large Bowel in the Cat. J. Physiol. 1933, 77, 422–431. [Google Scholar] [CrossRef] [Green Version]
- King, B.F.; Townsend-Nicholson, A. Involvement of P2Y 1 and P2Y 11 Purinoceptors in Parasympathetic Inhibition of Colonic Smooth Muscle. J. Pharm. Exp. 2008, 324, 1055–1063. [Google Scholar] [CrossRef] [Green Version]
- Kyloh, M.; Nicholas, S.; Zagorodnyuk, V.P.; Brookes, S.J.; Spencer, N.J. Identification of the Visceral Pain Pathway Activated by Noxious Colorectal Distension in Mice. Front. Neurosci. 2011, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Valentino, R.J.; Page, M.E.; Luppi, P.-H.; Zhu, Y.; Van Bockstaele, E.; Aston-Jones, G. Evidence for Widespread Afferents to Barrington’s Nucleus, a Brainstem Region Rich in Corticotropin-Releasing Hormone Neurons. Neuroscience 1994, 62, 125–143. [Google Scholar] [CrossRef]
- Ding, Y.-Q.; Zheng, H.-X.; Gong, L.-W.; Lu, Y.; Zhao, H.; Qin, B.-Z. Direct Projections from the Lumbosacral Spinal Cord to Barrington’s Nucleus in the Rat: A Special Reference to Micturition Reflex. J. Comp. Neurol. 1997, 389, 149–160. [Google Scholar] [CrossRef]
- De Groat, W.C.; Krier, J. The Sacral Parasympathetic Reflex Pathway Regulating Colonic Motility and Defaecation in the Cat. J. Physiol. 1978, 276, 481–500. [Google Scholar] [CrossRef] [PubMed]
- De Groat, W.C.; Nadelhaft, I.; Milne, R.J.; Booth, A.M.; Morgan, C.; Thor, K. Organization of the Sacral Parasympathetic Reflex Pathways to the Urinary Bladder and Large Intestine. J. Auton. Nerv. Syst. 1981, 3, 135–160. [Google Scholar] [CrossRef]
- Pavlov, I.P. The Work of the Digestive Glands; C. Griffin & Company Limited: London, UK, 1910. [Google Scholar]
- Cannon, W.B.; Newton, H.F.; Bright, E.M.; Menkin, V.; Moore, R.M. Some Aspects of the Physiology of Animals Surviving Complete Exclusion of Sympathetic Nerve Impulses. Am. J. Physiol.-Leg. Content 1929, 89, 84–107. [Google Scholar] [CrossRef]
- Bingham, J.R.; Ingelfinger, F.J.; Smithwick, R.H. The Effects of Sympathectomy on Abdominal Pain in Man. Gastroenterology 1950, 15, 18–33. [Google Scholar] [CrossRef]
- Kang, C.M.; Lee, H.Y.; Yang, H.J.; Jang, H.J.; Chun Gil, Y.; Kim, K.S.; Choi, J.S.; Lee, W.J.; Kim, B.R. Bilateral Thoracoscopic Splanchnicectomy with Sympathectomy for Managing Abdominal Pain in Cancer Patients. Am. J. Surg. 2007, 194, 23–29. [Google Scholar] [CrossRef]
- Rivera, L.R.; Poole, D.P.; Thacker, M.; Furness, J.B. The Involvement of Nitric Oxide Synthase Neurons in Enteric Neuropathies: NOS Neurons in Enteric Neuropathology. Neurogastroenterol. Motil. 2011, 23, 980–988. [Google Scholar] [CrossRef]
- Schubert, M.L.; Peura, D.A. Control of Gastric Acid Secretion in Health and Disease. Gastroenterology 2008, 134, 1842–1860. [Google Scholar] [CrossRef] [Green Version]
- Toumi, F.; Neunlist, M.; Cassagnau, E.; Parois, S.; Laboisse, C.L.; Galmiche, J.-P.; Jarry, A. Human Submucosal Neurones Regulate Intestinal Epithelial Cell Proliferation: Evidence from a Novel Co-Culture Model. Neurogastroenterol. Motil. 2003, 15, 239–242. [Google Scholar] [CrossRef]
- Bayliss, W.M.; Starling, E.H. The Movements and Innervation of the Small Intestine. J. Physiol. 1899, 24, 99–143. [Google Scholar] [CrossRef]
- Gwynne, R.M.; Thomas, E.A.; Goh, S.M.; Sjövall, H.; Bornstein, J.C. Segmentation Induced by Intraluminal Fatty Acid in Isolated Guinea-Pig Duodenum and Jejunum: Nutrient-Induced Segmentation in vitro. J. Physiol. 2004, 556, 557–569. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, T.Y.; Markus, H.; Diamant, N.E. Neural Control of the Intestinal Migrating Myoelectric Complex. A Pharmacological Analysis. Can. J. Physiol. Pharmacol. 1982, 60, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Sarna, S.; Stoddard, C.; Belbeck, L.; McWade, D. Intrinsic Nervous Control of Migrating Myoelectric Complexes. Am. J. Physiol.-Gastrointest. Liver Physiol. 1981, 241, G16–G23. [Google Scholar] [CrossRef] [PubMed]
- Ehrlein, H.J. Retroperistaltism and Duodenogastric Reflux in Dogs. Scand. J. Gastroenterol. Suppl. 1981, 67, 29–32. [Google Scholar]
- Vallance, B.A.; Blennerhassett, P.A.; Collins, S.M. Increased Intestinal Muscle Contractility and Worm Expulsion in Nematode-Infected Mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 1997, 272, G321–G327. [Google Scholar] [CrossRef]
- Lundgren, O. Enteric Nerves and Diarrhoea: Enteric Nerves, Secretion and Diarrhoea. Pharmacol. Toxicol. 2002, 90, 109–120. [Google Scholar] [CrossRef]
- Sjovall, H.; Abrahamsson, H.; Westlander, G.; Gillberg, R.; Redfors, S.; Jodal, M.; Lundgren, O. Intestinal Fluid and Electrolyte Transport in Man during Reduced Circulating Blood Volume. Gut 1986, 27, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Shirazi-Beechey, S.P.; Moran, A.W.; Batchelor, D.J.; Daly, K.; Al-Rammahi, M. Glucose Sensing and Signalling; Regulation of Intestinal Glucose Transport. Proc. Nutr. Soc. 2011, 70, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Gwynne, R.M.; Ellis, M.; Sjövall, H.; Bornstein, J.C. Cholera Toxin Induces Sustained Hyperexcitability in Submucosal Secretomotor Neurons in Guinea Pig Jejunum. Gastroenterology 2009, 136, 299–308.e4. [Google Scholar] [CrossRef]
- Sayegh, A.I.; Covasa, M.; Ritter, R.C. Intestinal Infusions of Oleate and Glucose Activate Distinct Enteric Neurons in the Rat. Auton. Neurosci. 2004, 115, 54–63. [Google Scholar] [CrossRef]
- Ferens, D.M.; Habgood, M.D.; Saunders, N.R.; Tan, Y.H.; Brown, D.J.; Brock, J.A.; Furness, J.B. Stimulation of Defecation in Spinal Cord-Injured Rats by a Centrally Acting Ghrelin Receptor Agonist. Spinal Cord 2011, 49, 1036–1041. [Google Scholar] [CrossRef] [Green Version]
- Brody, H. The Gut Microbiome. Nature 2020, 577, S5. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, M.; Moretti, R.; Campisciano, G.; da Silveira, A.B.M.; Monda, V.M.; Comar, M.; Di Bella, S.; Antonello, R.M.; Luzzati, R.; Crocè, L.S. You Talking to Me? Says the Enteric Nervous System (ENS) to the Microbe. How Intestinal Microbes Interact with the ENS. J. Clin. Med. 2020, 9, 3705. [Google Scholar] [CrossRef] [PubMed]
- Radocchia, G.; Neroni, B.; Marazzato, M.; Capuzzo, E.; Zuccari, S.; Pantanella, F.; Zenzeri, L.; Evangelisti, M.; Vassallo, F.; Parisi, P.; et al. Chronic Intestinal Pseudo-Obstruction: Is There a Connection with Gut Microbiota? Microorganisms 2021, 9, 2549. [Google Scholar] [CrossRef] [PubMed]
- Worthington, J.J.; Reimann, F.; Gribble, F.M. Enteroendocrine Cells-Sensory Sentinels of the Intestinal Environment and Orchestrators of Mucosal Immunity. Mucosal Immunol. 2018, 11, 3–20. [Google Scholar] [CrossRef]
- Sigalet, D.L.; Wallace, L.E.; Holst, J.J.; Martin, G.R.; Kaji, T.; Tanaka, H.; Sharkey, K.A. Enteric Neural Pathways Mediate the Anti-Inflammatory Actions of Glucagon-like Peptide 2. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 293, G211–G221. [Google Scholar] [CrossRef]
- Fried, S.; Wemelle, E.; Cani, P.D.; Knauf, C. Interactions between the Microbiota and Enteric Nervous System during Gut-Brain Disorders. Neuropharmacology 2021, 197, 108721. [Google Scholar] [CrossRef]
- Furness, J.B.; Jones, C.; Nurgali, K.; Clerc, N. Intrinsic Primary Afferent Neurons and Nerve Circuits within the Intestine. Prog. Neurobiol. 2004, 72, 143–164. [Google Scholar] [CrossRef]
- Fung, C.; Vanden Berghe, P. Functional Circuits and Signal Processing in the Enteric Nervous System. Cell. Mol. Life Sci. 2020, 77, 4505–4522. [Google Scholar] [CrossRef]
- Natale, G.; Ryskalin, L.; Morucci, G.; Lazzeri, G.; Frati, A.; Fornai, F. The Baseline Structure of the Enteric Nervous System and Its Role in Parkinson’s Disease. Life 2021, 11, 732. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. Types of Neurons in the Enteric Nervous System. J. Auton. Nerv. Syst. 2000, 81, 87–96. [Google Scholar] [CrossRef]
- Brun, P.; Giron, M.C.; Qesari, M.; Porzionato, A.; Caputi, V.; Zoppellaro, C.; Banzato, S.; Grillo, A.R.; Spagnol, L.; De Caro, R.; et al. Toll-like Receptor 2 Regulates Intestinal Inflammation by Controlling Integrity of the Enteric Nervous System. Gastroenterology 2013, 145, 1323–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-Chain Fatty Acids Activate GPR41 and GPR43 on Intestinal Epithelial Cells to Promote Inflammatory Responses in Mice. Gastroenterology 2013, 145, 396–406.e10. [Google Scholar] [CrossRef]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut Microbiota: The Neglected Endocrine Organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [Green Version]
- Brierley, S.M.; Linden, D.R. Neuroplasticity and Dysfunction after Gastrointestinal Inflammation. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 611–627. [Google Scholar] [CrossRef]
- Ojeda, J.; Ávila, A.; Vidal, P.M. Gut Microbiota Interaction with the Central Nervous System throughout Life. J. Clin. Med. 2021, 10, 1299. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.; Bercik, P. The Interplay between the Intestinal Microbiota and the Brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the Microbiota, Immune and Nervous Systems in Health and Disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef]
- Ma, E.L.; Smith, A.D.; Desai, N.; Cheung, L.; Hanscom, M.; Stoica, B.A.; Loane, D.J.; Shea-Donohue, T.; Faden, A.I. Bidirectional Brain-Gut Interactions and Chronic Pathological Changes after Traumatic Brain Injury in Mice. Brain Behav. Immun. 2017, 66, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shaanan, T.L.; Azulay-Debby, H.; Dubovik, T.; Starosvetsky, E.; Korin, B.; Schiller, M.; Green, N.L.; Admon, Y.; Hakim, F.; Shen-Orr, S.S.; et al. Activation of the Reward System Boosts Innate and Adaptive Immunity. Nat. Med. 2016, 22, 940–944. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Guan, N.L.; Kundu, P.; et al. The Gut Microbiota Influences Blood-Brain Barrier Permeability in Mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, K.M.; Kang, N.; Bienenstock, J.; Foster, J.A. Reduced Anxiety-like Behavior and Central Neurochemical Change in Germ-Free Mice. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2011, 23, 255-e119. [Google Scholar] [CrossRef] [PubMed]
- Ahern, G.P. 5-HT and the Immune System. Curr. Opin. Pharmacol. 2011, 11, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Barragan, A.; Weidner, J.M.; Jin, Z.; Korpi, E.R.; Birnir, B. GABAergic Signalling in the Immune System. Acta Physiol. 2015, 213, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Islas Weinstein, L.; Revuelta, A.; Pando, R.H. Catecholamines and Acetylcholine Are Key Regulators of the Interaction between Microbes and the Immune System. Ann. N. Y. Acad. Sci. 2015, 1351, 39–51. [Google Scholar] [CrossRef]
- Erny, D.; Angelis, A.L.H.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; DeLuca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan Catabolites from Microbiota Engage Aryl Hydrocarbon Receptor and Balance Mucosal Reactivity via Interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics Analysis Reveals Large Effects of Gut Microflora on Mammalian Blood Metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [Green Version]
- Rothhammer, V.; Mascanfroni, I.D.; Bunse, L.; Takenaka, M.C.; Kenison, J.E.; Mayo, L.; Chao, C.C.; Patel, B.; Yan, R.; Blain, M.; et al. Type I Interferons and Microbial Metabolites of Tryptophan Modulate Astrocyte Activity and Central Nervous System Inflammation via the Aryl Hydrocarbon Receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Gao, X.; Peng, Y.; Wu, Q.; Zhu, J.; Tan, C.; Xia, G.; You, C.; Xu, R.; Pan, S.; et al. Higher Risk of Stroke Is Correlated with Increased Opportunistic Pathogen Load and Reduced Levels of Butyrate-Producing Bacteria in the Gut. Front. Cell. Infect. Microbiol. 2019, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.; Wu, Q.; Wang, H.; Gao, X.; Xu, R.; Cui, Z.; Zhu, J.; Zeng, X.; Zhou, H.; He, Y.; et al. Dysbiosis of Gut Microbiota and Short-Chain Fatty Acids in Acute Ischemic Stroke and the Subsequent Risk for Poor Functional Outcomes. JPEN J. Parenter. Enter. Nutr. 2021, 45, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Coman, V.; Vodnar, D.C. Gut Microbiota and Old Age: Modulating Factors and Interventions for Healthy Longevity. Exp. Gerontol. 2020, 141, 111095. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; D’Aigle, J.; Atadja, L.; Quaicoe, V.; Honarpisheh, P.; Ganesh, B.P.; Hassan, A.; Graf, J.; Petrosino, J.; Putluri, N.; et al. Gut Microbiota-Derived Short-Chain Fatty Acids Promote Poststroke Recovery in Aged Mice. Circ. Res. 2020, 127, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Benakis, C.; Brea, D.; Caballero, S.; Faraco, G.; Moore, J.; Murphy, M.; Sita, G.; Racchumi, G.; Ling, L.; Pamer, E.G.; et al. Commensal Microbiota Affects Ischemic Stroke Outcome by Regulating Intestinal Γδ T Cells. Nat. Med. 2016, 22, 516–523. [Google Scholar] [CrossRef]
- Winek, K.; Engel, O.; Koduah, P.; Heimesaat, M.M.; Fischer, A.; Bereswill, S.; Dames, C.; Kershaw, O.; Gruber, A.D.; Curato, C.; et al. Depletion of Cultivatable Gut Microbiota by Broad-Spectrum Antibiotic Pretreatment Worsens Outcome After Murine Stroke. Stroke 2016, 47, 1354–1363. [Google Scholar] [CrossRef]
- Gungor, B.; Adiguzel, E.; Gursel, I.; Yilmaz, B.; Gursel, M. Intestinal Microbiota in Patients with Spinal Cord Injury. PLoS ONE 2016, 11, e0145878. [Google Scholar] [CrossRef] [Green Version]
- Bazzocchi, G.; Turroni, S.; Bulzamini, M.C.; D’Amico, F.; Bava, A.; Castiglioni, M.; Cagnetta, V.; Losavio, E.; Cazzaniga, M.; Terenghi, L.; et al. Changes in Gut Microbiota in the Acute Phase after Spinal Cord Injury Correlate with Severity of the Lesion. Sci. Rep. 2021, 11, 12743. [Google Scholar] [CrossRef]
- Schmidt, E.K.A.; Torres-Espin, A.; Raposo, P.J.F.; Madsen, K.L.; Kigerl, K.A.; Popovich, P.G.; Fenrich, K.K.; Fouad, K. Fecal Transplant Prevents Gut Dysbiosis and Anxiety-like Behaviour after Spinal Cord Injury in Rats. PLoS ONE 2020, 15, e0226128. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Hall, J.C.E.; Wang, L.; Mo, X.; Yu, Z.; Popovich, P.G. Gut Dysbiosis Impairs Recovery after Spinal Cord Injury. J. Exp. Med. 2016, 213, 2603–2620. [Google Scholar] [CrossRef] [PubMed]
- Hanscom, M.; Loane, D.J.; Shea-Donohue, T. Brain-Gut Axis Dysfunction in the Pathogenesis of Traumatic Brain Injury. J. Clin. Investig. 2021, 131, e143777. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Tang, W.; Zhou, C.; Sun, X.; Wei, Z.; Zhong, J.; Huang, Z. Fecal Microbiota Transplantation Is a Promising Method to Restore Gut Microbiota Dysbiosis and Relieve Neurological Deficits after Traumatic Brain Injury. Oxidative Med. Cell. Longev. 2021, 2021, 5816837. [Google Scholar] [CrossRef] [PubMed]
- Opeyemi, O.M.; Rogers, M.B.; Firek, B.A.; Janesko-Feldman, K.; Vagni, V.; Mullett, S.J.; Wendell, S.G.; Nelson, B.P.; New, L.A.; Mariño, E.; et al. Sustained Dysbiosis and Decreased Fecal Short-Chain Fatty Acids after Traumatic Brain Injury and Impact on Neurologic Outcome. J. Neurotrauma 2021, 38, 2610–2621. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.W.; Rogers, M.B.; Gao, Y.; Vincent, G.; Firek, B.A.; Janesko-Feldman, K.; Vagni, V.; Kochanek, P.M.; Ozolek, J.A.; Mollen, K.P.; et al. Depletion of Gut Microbiota Is Associated with Improved Neurologic Outcome Following Traumatic Brain Injury. Brain Res. 2020, 1747, 147056. [Google Scholar] [CrossRef]
- Angoa-Pérez, M.; Zagorac, B.; Anneken, J.H.; Briggs, D.I.; Winters, A.D.; Greenberg, J.M.; Ahmad, M.; Theis, K.R.; Kuhn, D.M. Repetitive, Mild Traumatic Brain Injury Results in a Progressive White Matter Pathology, Cognitive Deterioration, and a Transient Gut Microbiota Dysbiosis. Sci. Rep. 2020, 10, 8949. [Google Scholar] [CrossRef]
- Tang, A.T.; Choi, J.P.; Kotzin, J.J.; Yang, Y.; Hong, C.C.; Hobson, N.; Girard, R.; Zeineddine, H.A.; Lightle, R.; Moore, T.; et al. Endothelial TLR4 and the Microbiome Drive Cerebral Cavernous Malformations. Nature 2017, 545, 305–310. [Google Scholar] [CrossRef]
- Mouchtouris, N.; Chalouhi, N.; Chitale, A.; Starke, R.M.; Tjoumakaris, S.I.; Rosenwasser, R.H.; Jabbour, P.M. Management of Cerebral Cavernous Malformations: From Diagnosis to Treatment. Sci. World J. 2015, 2015, 808314. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zhou, G.; Shao, B.; Zhou, H.; Xu, C.; Yan, F.; Wang, L.; Chen, G.; Li, J.; Fu, X. Gut Microbiota Dysbiosis Induced by Intracerebral Hemorrhage Aggravates Neuroinflammation in Mice. Front. Microbiol. 2021, 12, 876. [Google Scholar] [CrossRef]
- Zhai, Q.; Sun, T.; Sun, C.; Yan, L.; Wang, X.; Wang, Y.; Sun, J.; Zhao, Y. High Plasma Levels of Trimethylamine N-Oxide Are Associated with Poor Outcome in Intracerebral Hemorrhage Patients. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2021, 42, 1009–1016. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, J.; Wang, J.; Yang, Y.; Huang, J.; Gong, H.; Cui, H.; Chen, D. Successful Treatment with Fecal Microbiota Transplantation in Patients with Multiple Organ Dysfunction Syndrome and Diarrhea Following Severe Sepsis. Crit. Care 2016, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivancos, J.; Gilo, F.; Frutos, R.; Maestre, J.; García-Pastor, A.; Quintana, F.; Roda, J.M.; Ximénez-Carrillo, A.; Díez Tejedor, E.; Fuentes, B.; et al. Clinical Management Guidelines for Subarachnoid Haemorrhage. Diagnosis and Treatment. Neurología 2014, 29, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, R.L.; Schweizer, T.A. Spontaneous Subarachnoid Haemorrhage. Lancet 2017, 389, 655–666. [Google Scholar] [CrossRef]
- Shikata, F.; Shimada, K.; Sato, H.; Ikedo, T.; Kuwabara, A.; Furukawa, H.; Korai, M.; Kotoda, M.; Yokosuka, K.; Makino, H.; et al. Potential Influences of Gut Microbiota on the Formation of Intracranial Aneurysm. Hypertension 2019, 73, 491–496. [Google Scholar] [CrossRef]
- Vendrik, K.E.W.; Ooijevaar, R.E.; de Jong, P.R.C.; Laman, J.D.; van Oosten, B.W.; van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal Microbiota Transplantation in Neurological Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef] [Green Version]
- Yuan, B.; Lu, X.-J.; Wu, Q. Gut Microbiota and Acute Central Nervous System Injury: A New Target for Therapeutic Intervention. Front. Immunol. 2021, 12, 800796. [Google Scholar] [CrossRef]
- Barrio, C.; Arias-Sánchez, S.; Martín-Monzón, I. The Gut Microbiota-Brain Axis, Psychobiotics and Its Influence on Brain and Behaviour: A Systematic Review. Psychoneuroendocrinology 2021, 137, 105640. [Google Scholar] [CrossRef]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of Gut-Brain Axis, Gut Microbial Composition, and Probiotic Intervention in Alzheimer’s Disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef]
- Pluta, R.; Januszewski, S.; Czuczwar, S.J. The Role of Gut Microbiota in an Ischemic Stroke. Int. J. Mol. Sci. 2021, 22, 915. [Google Scholar] [CrossRef]
- Baldi, S.; Mundula, T.; Nannini, G.; Amedei, A. Microbiota Shaping–The Effects of Probiotics, Prebiotics, and Fecal Microbiota Transplant on Cognitive Functions: A Systematic Review. World J. Gastroenterol. 2021, 27, 6715–6732. [Google Scholar] [CrossRef]
- Setbo, E.; Campbell, K.; O’Cuiv, P.; Hubbard, R. Utility of Probiotics for Maintenance or Improvement of Health Status in Older People–A Scoping Review. J. Nutr. Health Aging 2019, 23, 364–372. [Google Scholar] [CrossRef] [PubMed]
- George, A.K.; Behera, J.; Homme, R.P.; Tyagi, N.; Tyagi, S.C.; Singh, M. Rebuilding Microbiome for Mitigating Traumatic Brain Injury: Importance of Restructuring the Gut-Microbiome-Brain Axis. Mol. Neurobiol. 2021, 58, 3614–3627. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.W.; Pandya, J.D.; Shear, D.A. Gut Microbiota as a Therapeutic Target to Ameliorate the Biochemical, Neuroanatomical, and Behavioral Effects of Traumatic Brain Injuries. Front. Neurol. 2019, 10, 875. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, F.; Ling, Z.; Yu, X.; Chen, W.; Li, H.; Jin, J.; Pang, M.; Zhang, H.; Yu, J.; et al. Clostridium Butyricum Attenuates Cerebral Ischemia/Reperfusion Injury in Diabetic Mice via Modulation of Gut Microbiota. Brain Res. 2016, 1642, 180–188. [Google Scholar] [CrossRef]
- Li, H.; Sun, J.; Du, J.; Wang, F.; Fang, R.; Yu, C.; Xiong, J.; Chen, W.; Lu, Z.; Liu, J. Clostridium Butyricum Exerts a Neuroprotective Effect in a Mouse Model of Traumatic Brain Injury via the Gut-Brain Axis. Neurogastroenterol. Motil. 2018, 30, e13260. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, T.; Fu, J.; Fu, S.; Hu, C.; Sun, B.; Fan, X.; Zhu, J. Lactobacillus Acidophilus Exerts Neuroprotective Effects in Mice with Traumatic Brain Injury. J. Nutr. 2019, 149, 1543–1552. [Google Scholar] [CrossRef]
- Akhoundzadeh, K.; Vakili, A.; Shadnoush, M.; Sadeghzadeh, J. Effects of the Oral Ingestion of Probiotics on Brain Damage in a Transient Model of Focal Cerebral Ischemia in Mice. Iran. J. Med. Sci. 2018, 43, 32–40. [Google Scholar]
- Brenner, L.A.; Stearns-Yoder, K.A.; Hoffberg, A.S.; Penzenik, M.E.; Starosta, A.J.; Hernández, T.D.; Hadidi, D.A.; Lowry, C.A. Growing Literature but Limited Evidence: A Systematic Review Regarding Prebiotic and Probiotic Interventions for Those with Traumatic Brain Injury and/or Posttraumatic Stress Disorder. Brain Behav. Immun. 2017, 65, 57–67. [Google Scholar] [CrossRef]
- Falcão de Arruda, I.S.; de Aguilar-Nascimento, J.E. Benefits of Early Enteral Nutrition with Glutamine and Probiotics in Brain Injury Patients. Clin. Sci. 2004, 106, 287–292. [Google Scholar] [CrossRef]
- Tan, M.; Zhu, J.-C.; Du, J.; Zhang, L.-M.; Yin, H.-H. Effects of Probiotics on Serum Levels of Th1/Th2 Cytokine and Clinical Outcomes in Severe Traumatic Brain-Injured Patients: A Prospective Randomized Pilot Study. Crit. Care 2011, 15, R290. [Google Scholar] [CrossRef] [Green Version]
- Wan, G.; Wang, L.; Zhang, G.; Zhang, J.; Lu, Y.; Li, J.; Yi, X. Effects of Probiotics Combined with Early Enteral Nutrition on Endothelin-1 and C-Reactive Protein Levels and Prognosis in Patients with Severe Traumatic Brain Injury. J. Int. Med. Res. 2020, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Painter, T.J.; Rickerds, J.; Alban, R.F. Immune Enhancing Nutrition in Traumatic Brain Injury–A Preliminary Study. Int. J. Surg. 2015, 21, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Snydman, D.R. The Safety of Probiotics. Clin. Infect. Dis. 2008, 46 (Suppl. 2), S104–S111. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Zhang, X.; Zheng, Y.; Yu, B.; Pan, D.; Jiang, X.; Yan, C.; Yu, Q.; Lu, X. Distinctive Gut Microbiota Alteration Is Associated with Poststroke Functional Recovery: Results from a Prospective Cohort Study. Neural Plast. 2021, 2021, 1469339. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.-J.; Yu, Q.; Li, Y.; Feng, D.; Li, Y.-M. Dysbiosis of Gut Microbiota in Patients with Post-Stroke Cognitive Impairment. Sichuan Da Xue Xue Bao Yi Xue Ban 2021, 52, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Shen, Z.; He, W. Identification of Gut Microbiome Signatures in Patients With Post-Stroke Cognitive Impairment and Affective Disorder. Front. Aging Neurosci. 2021, 13, 706765. [Google Scholar] [CrossRef]
- Singh, V.; Roth, S.; Llovera, G.; Sadler, R.; Garzetti, D.; Stecher, B.; Dichgans, M.; Liesz, A. Microbiota Dysbiosis Controls the Neuroinflammatory Response after Stroke. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 7428–7440. [Google Scholar] [CrossRef]
- Jing, Y.; Yu, Y.; Bai, F.; Wang, L.; Yang, D.; Zhang, C.; Qin, C.; Yang, M.; Zhang, D.; Zhu, Y.; et al. Effect of Fecal Microbiota Transplantation on Neurological Restoration in a Spinal Cord Injury Mouse Model: Involvement of Brain-Gut Axis. Microbiome 2021, 9, 59. [Google Scholar] [CrossRef]
- Chen, R.; Xu, Y.; Wu, P.; Zhou, H.; Lasanajak, Y.; Fang, Y.; Tang, L.; Ye, L.; Li, X.; Cai, Z.; et al. Transplantation of Fecal Microbiota Rich in Short Chain Fatty Acids and Butyric Acid Treat Cerebral Ischemic Stroke by Regulating Gut Microbiota. Pharm. Res. 2019, 148, 104403. [Google Scholar] [CrossRef]
- Zhu, W.; Romano, K.A.; Li, L.; Buffa, J.A.; Sangwan, N.; Prakash, P.; Tittle, A.N.; Li, X.S.; Fu, X.; Androjna, C.; et al. Gut Microbes Impact Stroke Severity via the Trimethylamine N-Oxide Pathway. Cell Host Microbe 2021, 29, 1199–1208.e5. [Google Scholar] [CrossRef]
- Delgado Jiménez, R.; Benakis, C. The Gut Ecosystem: A Critical Player in Stroke. Neuromol. Med. 2021, 23, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.; Dandekar, M.P. Crosstalk between Gut Microbiome and Immunology in the Management of Ischemic Brain Injury. J. Neuroimmunol. 2021, 353, 577498. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-M.; Yu, R.; Zhang, L.-P.; Wen, S.-Y.; Wang, S.-J.; Zhang, X.-Y.; Xu, Q.; Kong, L.-D. Dietary Fructose-Induced Gut Dysbiosis Promotes Mouse Hippocampal Neuroinflammation: A Benefit of Short-Chain Fatty Acids. Microbiome 2019, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Sadler, R.; Cramer, J.V.; Heindl, S.; Kostidis, S.; Betz, D.; Zuurbier, K.R.; Northoff, B.H.; Heijink, M.; Goldberg, M.P.; Plautz, E.J.; et al. Short-Chain Fatty Acids Improve Poststroke Recovery via Immunological Mechanisms. J. Neurosci. 2020, 40, 1162–1173. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Ban, M.; Kishi, M.; Ono, T. Safety Evaluation of the Excessive Intake of Ceramide-Containing Acetic Acid Bacteria–A Randomized, Double-Blind, Placebo-Controlled Study Over a 4-Week Period. J. Oleo Sci. 2021, 70, 417–430. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, T.; Wang, Y.; Si, C.; Wang, X.; Wang, R.-T.; Lv, Z. Baicalin Ameliorates Neuropathology in Repeated Cerebral Ischemia-Reperfusion Injury Model Mice by Remodeling the Gut Microbiota. Aging 2020, 12, 3791–3806. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Zhang, D.-X.; Zheng, W.-C.; Hu, J.-S.; Fu, L.; Li, Y.; Xin, Y.; Wang, X.-P. CORM-3 Exerts a Neuroprotective Effect in a Rodent Model of Traumatic Brain Injury via the Bidirectional Gut-Brain Interactions. Exp. Neurol. 2021, 341, 113683. [Google Scholar] [CrossRef]
- Pang, S.Q.; Luo, Z.T.; Wang, C.C.; Hong, X.P.; Zhou, J.; Chen, F.; Ge, L.; Li, X.; Dai, Y.L.; Wu, Y.L.; et al. Effects of Dioscorea Polystachya “yam Gruel” on the Cognitive Function of Diabetic Rats with Focal Cerebral Ischemia-Reperfusion Injury via the Gut-Brain Axis. J. Integr. Neurosci. 2020, 19, 273–283. [Google Scholar] [CrossRef]


{kind=link}
| Intervention | Type of Research | Main Findings | Advantages | Disadvantages |
|---|---|---|---|---|
| Probiotics/ prebiotics | Clinical, preclinical (mice) |
|
|
|
| Fecal microbiota transplant | Preclinical (mice, rats) |
|
|
|
| Oral short-chain fatty acids | Preclinical (mice) |
|
|
|
| Baicalin | Preclinical (mice) |
|
|
|
| CORM-3 | Preclinical (rats) |
|
|
|
| Dioscorea polystachya | Preclinical (rats) |
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panther, E.J.; Dodd, W.; Clark, A.; Lucke-Wold, B. Gastrointestinal Microbiome and Neurologic Injury. Biomedicines 2022, 10, 500. https://doi.org/10.3390/biomedicines10020500
Panther EJ, Dodd W, Clark A, Lucke-Wold B. Gastrointestinal Microbiome and Neurologic Injury. Biomedicines. 2022; 10(2):500. https://doi.org/10.3390/biomedicines10020500
Chicago/Turabian StylePanther, Eric J., William Dodd, Alec Clark, and Brandon Lucke-Wold. 2022. "Gastrointestinal Microbiome and Neurologic Injury" Biomedicines 10, no. 2: 500. https://doi.org/10.3390/biomedicines10020500