
The Role of KLF2 in the Regulation of Atherosclerosis Development and Potential Use of KLF2-Targeted Therapy

 , and
, and 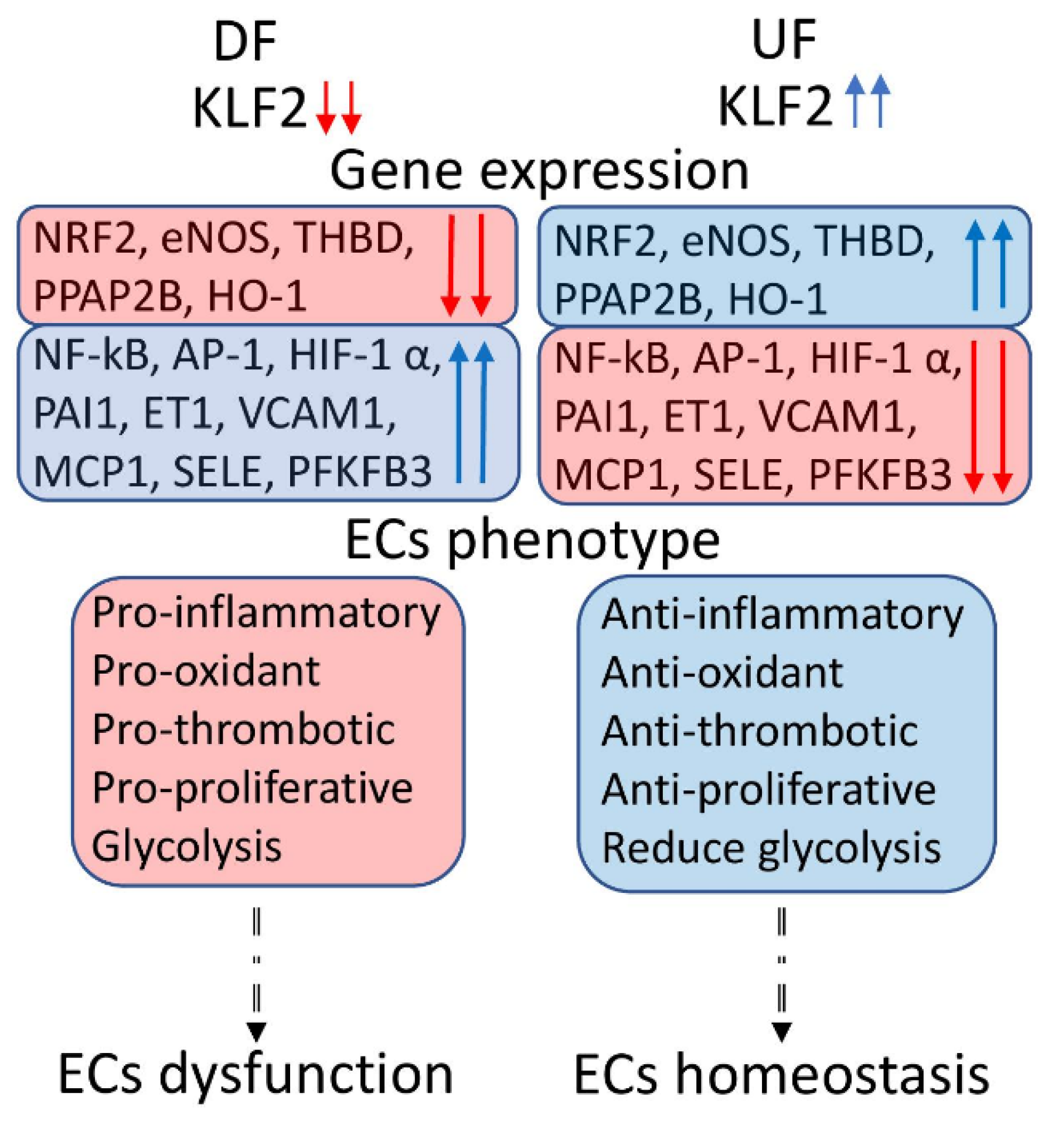{kind=link}
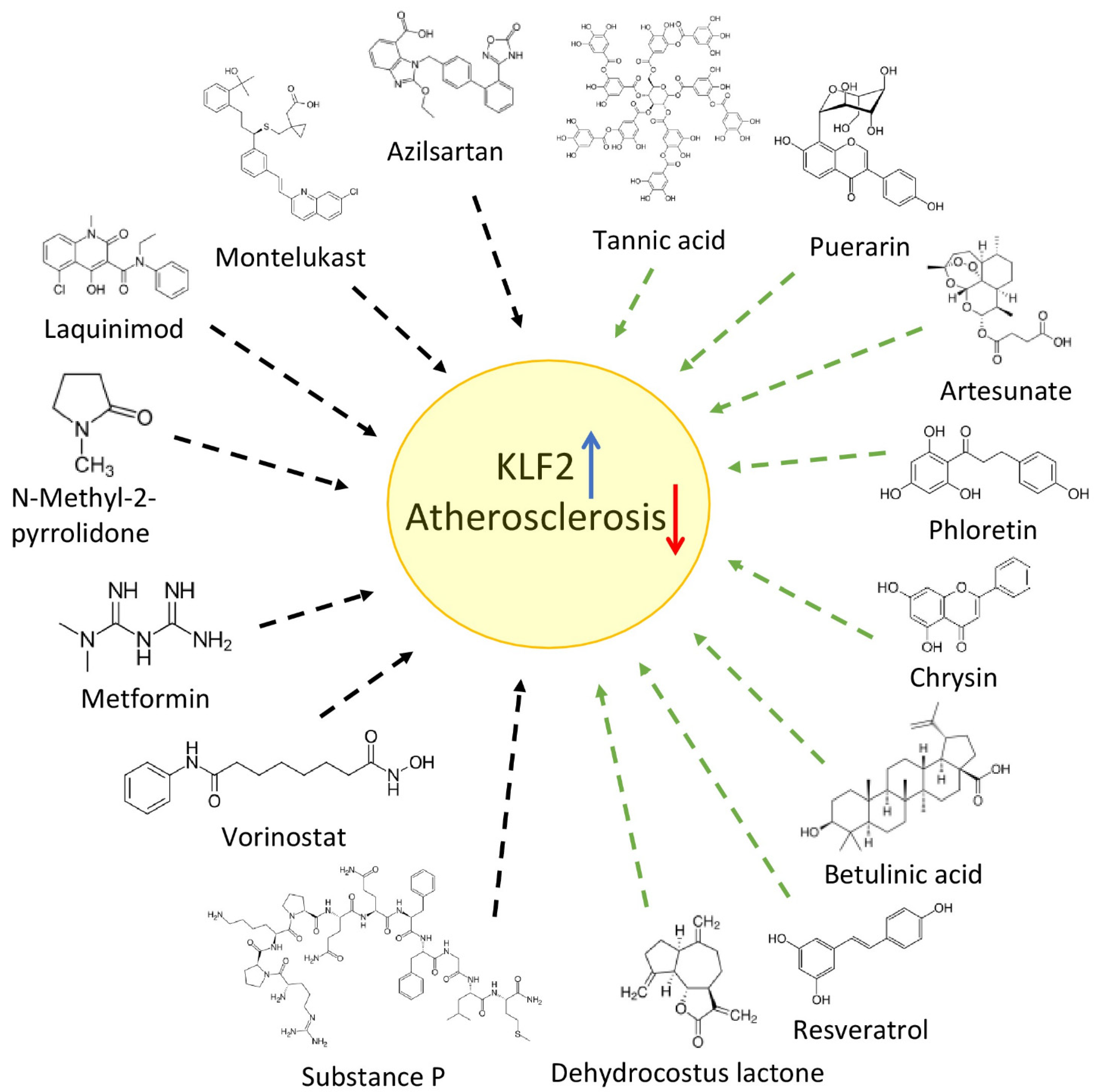{kind=link}
Abstract
:1. Introduction
1.1. Atherosclerosis and Shear Stress
1.2. Kruppel like Factor 2
2. KLF2 as the Main Regulator of the NO Production
2.1. KLF2-Based eNOS/NO Regulation
2.2. Genes Regulating KLF2
3. Repurposing Drugs to Treat Atherosclerosis (Acting on KLF2)
3.1. Anti-Diabetic Drugs
The Role KLF2 in Autophagy Activation
4. Plant Compounds
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CVDs | cardiovascular diseases |
| VSMCs | vascular smooth muscle cells |
| LDL | low-density lipoproteins |
| DF | disturbed flow |
| UF | unidirectional laminar flow |
| MSTFs | mechanosensitive transcription factors |
| THBD | thrombomodulin |
| PAI1 | plasminogen activator inhibitor-1 |
| SELE | E-selectin |
| ET1 | endothelin 1 |
| MCP-1 | monocyte chemoattractant protein |
| VCAM1 | vascular cell adhesion molecule |
| PPAP2B | phosphatidic acid phosphatase type 2B |
| PFKFB3 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase-3 |
| NO | nitric oxide |
| ECs | endothelial cells |
| eNOS | endothelial nitric oxide synthase |
| TGFB1 | transforming growth factor beta 1 |
| TNFα | tumor necrosis factor α |
| MAZ | MYC-associated zinc finger protein |
| AP-1 | jun proto-oncogene |
| ELF1 | E74-like ETS transcription factor 1 |
| SP1 | specificity protein 1 |
| LOX-1 | lectin-like oxidized low-density lipoprotein receptor-1 |
| AP-1 | activator protein-1 |
| HN | humanin |
| HRD1 | 3-hydroxy-3-methylglutaryl reductase degradation |
| NRF2 | nuclear factor, erythroid 2 like 2 |
| HO-1 | heme oxygenase 1 |
| AMPK | protein kinase, AMP-activated, alpha 2 catalytic subunit |
| HIF-1α | hypoxia-inducible factor 1 subunit alpha |
| NF-kB | nuclear factor kappa B subunit 1 |
| ox-LDL | oxidation of low-density lipoproteins |
| GPR81 | G coupled-protein receptor 81 |
| HCA1 | hydroxycarboxylic acid receptor 1 |
| HMGB1 | high mobility group box 1 |
| IRF2BP2 | interferon regulatory factor 2-binding protein 2 |
| AGE | advanced glycation end product |
| FOXP1 | forkhead box P1 |
| Nlrp3 | NLR family pyrin domain containing 3 |
| BDNF | brain-derived neurotrophic factor |
| HK1 | hexokinase 1 |
| HDAC4 | histone deacetylases 4 |
| MEF2 | myocyte enhancer factor 2C |
| CysLT1 | cysteinyl leukotriene receptor 1 |
| NMP | N-methyl-2-pyrrolidone |
| SAHA | suberanilohydroxamic acid |
| GLP-1 | glucagon-like peptide 1 |
| GPR120 | free fatty acid receptor 4 |
| AAA | abdominal aortic aneurysms |
| ICAM-1 | intercellular adhesion molecule-1 |
| ESL | ethanol extract of soy leaf |
| MMP-9 | matrix metallopeptidase 9 |
| LPL | lipoprotein lipase |
| VSMC | vascular smooth muscle cells |
| BA | betulinic acid |
| TRPC1 | capacitative calcium channel protein Trp1 |
| CaMKIIα | calcium/calmodulin-dependent protein kinase II Alpha |
| CaMKKβ | calcium/calmodulin-dependent protein kinase kinase 2 |
References
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 Update: A Report From the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef] [PubMed]
- Mallick, R.; Duttaroy, A.K. Modulation of endothelium function by fatty acids. Mol. Cell. Biochem. 2021, 477, 15–38. [Google Scholar] [CrossRef] [PubMed]
- Kasikara, C.; Doran, A.C.; Cai, B.; Tabas, I. The role of non-resolving inflammation in atherosclerosis. J. Clin. Investig. 2018, 128, 2713–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myasoedova, V.A.; Di Minno, A.; Songia, P.; Massaiu, I.; Alfieri, V.; Valerio, V.; Moschetta, D.; Andreini, D.; Alamanni, F.; Pepi, M.; et al. Sex-specific differences in age-related aortic valve calcium load: A systematic review and meta-analysis. Ageing Res. Rev. 2020, 61, 101077. [Google Scholar] [CrossRef] [PubMed]
- Gliozzi, M.; Scicchitano, M.; Bosco, F.; Musolino, V.; Carresi, C.; Scarano, F.; Maiuolo, J.; Nucera, S.; Maretta, A.; Paone, S.; et al. Modulation of Nitric Oxide Synthases by Oxidized LDLs: Role in Vascular Inflammation and Atherosclerosis Development. Int. J. Mol. Sci. 2019, 20, 3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunderland, K.; Jiang, J.; Zhao, F. Disturbed flow’s impact on cellular changes indicative of vascular aneurysm initiation, expansion, and rupture: A pathological and methodological review. J. Cell. Physiol. 2021. [Google Scholar] [CrossRef]
- Mo, F.-E. Shear-Regulated Extracellular Microenvironments and Endothelial Cell Surface Integrin Receptors Intertwine in Atherosclerosis. Front. Cell Dev. Biol. 2021, 9, 640781. [Google Scholar] [CrossRef]
- Urschel, K.; Tauchi, M.; Achenbach, S.; Dietel, B. Investigation of Wall Shear Stress in Cardiovascular Research and in Clinical Practice—From Bench to Bedside. Int. J. Mol. Sci. 2021, 22, 5635. [Google Scholar] [CrossRef]
- Cameron, J.N.; Mehta, O.H.; Michail, M.; Chan, J.; Nicholls, S.J.; Bennett, M.R.; Brown, A.J. Exploring the relationship between biomechanical stresses and coronary atherosclerosis. Atherosclerosis 2020, 302, 43–51. [Google Scholar] [CrossRef]
- Roux, E.; Bougaran, P.; Dufourcq, P.; Couffinhal, T. Fluid Shear Stress Sensing by the Endothelial Layer. Front. Physiol. 2020, 11, 861. [Google Scholar] [CrossRef]
- Theodorou, K.; Boon, R.A. Endothelial Cell Metabolism in Atherosclerosis. Front. Cell Dev. Biol. 2018, 6, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eelen, G.; De Zeeuw, P.; Treps, L.; Harjes, U.; Wong, B.; Carmeliet, P. Endothelial Cell Metabolism. Physiol. Rev. 2018, 98, 3–58. [Google Scholar] [CrossRef] [PubMed]
- Sweet, D.R.; Fan, L.; Hsieh, P.N.; Jain, M.K. Krüppel-Like Factors in Vascular Inflammation: Mechanistic Insights and Therapeutic Potential. Front. Cardiovasc. Med. 2018, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, R.D.; Kumar, S.; Jo, H. The role of endothelial mechanosensitive genes in atherosclerosis and omics approaches. Arch. Biochem. Biophys. 2015, 591, 111–131. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.N.; Fan, L.; Sweet, D.R.; Jain, M.K. The Krüppel-Like Factors and Control of Energy Homeostasis. Endocr. Rev. 2018, 40, 137–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, H.; Chen, S.; He, J.; Chen, W.; Ding, Y.; Huang, J. P300/CBP-associated factor (PCAF) attenuated M1 macrophage inflammatory responses possibly through KLF2 and KLF4. Immunol. Cell Biol. 2021, 99, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Ulfhammer, E.; Larsson, P.; Magnusson, M.; Karlsson, L.; Bergh, N.; Jern, S. Dependence of Proximal GC Boxes and Binding Transcription Factors in the Regulation of Basal and Valproic Acid-Induced Expression of t-PA. Int. J. Vasc. Med. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Veseli, B.E.; Perrotta, P.; De Meyer, G.R.A.; Roth, L.; Van der Donckt, C.; Martinet, W.; De Meyer, G.R.Y. Animal models of atherosclerosis. Eur. J. Pharmacol. 2017, 816, 3–13. [Google Scholar] [CrossRef]
- Wu, C.; Huang, R.-T.; Kuo, C.-H.; Kumar, S.; Kim, C.W.; Lin, Y.-C.; Chen, Y.-J.; Birukova, A.; Birukov, K.G.; Dulin, N.O.; et al. Mechanosensitive PPAP2B Regulates Endothelial Responses to Atherorelevant Hemodynamic Forces. Circ. Res. 2015, 117, e41–e53. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.D.; Huang, R.-T.; Wu, D.; Shentu, T.-P.; Harrison, D.L.; Whalen, M.B.; Stolze, L.K.; Di Rienzo, A.; Moskowitz, I.P.; Civelek, M.; et al. Genetic variant at coronary artery disease and ischemic stroke locus 1p32.2 regulates endothelial responses to hemodynamics. Proc. Natl. Acad. Sci. USA 2018, 115, E11349–E11358. [Google Scholar] [CrossRef] [Green Version]
- Doddaballapur, A.; Michalik, K.M.; Manavski, Y.; Lucas, T.; Houtkooper, R.H.; You, X.; Chen, W.; Zeiher, A.M.; Potente, M.; Dimmeler, S.; et al. Laminar Shear Stress Inhibits Endothelial Cell Metabolism via KLF2-Mediated Repression of PFKFB3. Arter. Thromb. Vasc. Biol. 2015, 35, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhoutte, P.M. Nitric Oxide: From Good to Bad. Ann. Vasc. Dis. 2018, 11, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimokawa, H.; Godo, S. Nitric oxide and endothelium-dependent hyperpolarization mediated by hydrogen peroxide in health and disease. Basic Clin. Pharmacol. Toxicol. 2020, 127, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.; Sessa, W.C. Endothelial NOS: Perspective and recent developments. J. Cereb. Blood Flow Metab. 2019, 176, 189–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daiber, A.; Xia, N.; Steven, S.; Oelze, M.; Hanf, A.; Kröller-Schön, S.; Münzel, T.; Li, H. New Therapeutic Implications of Endothelial Nitric Oxide Synthase (eNOS) Function/Dysfunction in Cardiovascular Disease. Int. J. Mol. Sci. 2019, 20, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kattoor, A.J.; Goel, A.; Mehta, J.L. LOX-1: Regulation, Signaling and Its Role in Atherosclerosis. Antioxidants 2019, 8, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Chung, J.; Kim, K.H.; An, S.H.; Kim, M.; Park, J.; Kwon, K. Fluid shear stress regulates the expression of Lectin-like oxidized low density lipoprotein receptor-1 via KLF2-AP-1 pathway depending on its intensity and pattern in endothelial cells. Atherosclerosis 2018, 270, 76–88. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Nikiforov, N.G.; Starodubova, A.V.; Popkova, T.V.; Orekhov, A.N. The Role of Mitochondria-Derived Peptides in Cardiovascular Diseases and Their Potential as Therapeutic Targets. Int. J. Mol. Sci. 2021, 22, 8770. [Google Scholar] [CrossRef]
- Merry, T.L.; Chan, A.; Woodhead, J.S.T.; Reynolds, J.C.; Kumagai, H.; Kim, S.-J.; Lee, C. Mitochondrial-derived peptides in energy metabolism. Am. J. Physiol. Metab. 2020, 319, E659–E666. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Z.; He, Y.; Zhang, H.; Tian, L.; Zheng, C.; Shang, T.; Zhu, Q.; Li, D.; He, Y. Humanin prevents high glucose-induced monocyte adhesion to endothelial cells by targeting KLF2. Mol. Immunol. 2018, 101, 245–250. [Google Scholar] [CrossRef]
- Li, Q.; Xuan, W.; Jia, Z.; Li, H.; Li, M.; Liang, X.; Su, D. HRD1 prevents atherosclerosis-mediated endothelial cell apoptosis by promoting LOX-1 degradation. Cell Cycle 2020, 19, 1466–1477. [Google Scholar] [CrossRef] [PubMed]
- Njock, M.-S.; Cheng, H.S.; Dang, L.T.; Nazari-Jahantigh, M.; Lau, A.C.; Boudreau, E.; Roufaiel, M.; Cybulsky, M.I.; Schober, A.; Fish, J.E. Endothelial cells suppress monocyte activation through secretion of extracellular vesicles containing antiinflammatory microRNAs. Blood 2015, 125, 3202–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Wu, C.; Xiao, J.; Li, D.; Sun, Z.; Li, M. Endothelial extracellular vesicles modulate the macrophage phenotype: Potential implications in atherosclerosis. Scand. J. Immunol. 2018, 87, e12648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.; Sun, Y.; Gao, Y.; Guan, X.; Yan, H.; Cui, X.; Zhang, X.; Li, X.; Li, H.; Cheng, M. Function of Krüppel-like factor 2 in the shear stress-induced cell differentiation of endothelial progenitor cells to endothelial cells. Mol. Med. Rep. 2019, 19, 1739–1746. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Geng, P.; Zhu, J.; Li, J.; Zhang, L.; Chen, W.; Zhang, D.; Lu, Y.; Xu, X. KLF2 regulates eNOS uncoupling via Nrf2/HO-1 in endothelial cells under hypoxia and reoxygenation. Chem. Interact. 2019, 305, 105–111. [Google Scholar] [CrossRef]
- Chen, G.-H.; Li, X.-L.; Deng, Y.-Q.; Zhou, F.-M.; Zou, W.-Q.; Jiang, W.-X.; Shangguan, S.-Q.; Lu, Z.-N. The Molecular Mechanism of EPO Regulates the Angiogenesis after Cerebral Ischemia through AMPK-KLF2 Signaling Pathway. Crit. Rev. Eukaryot. Gene Expr. 2019, 29, 105–112. [Google Scholar] [CrossRef]
- Akhtar, S.; Hartmann, P.; Karshovska, E.; Rinderknecht, F.-A.; Subramanian, P.; Gremse, F.; Grommes, J.; Jacobs, M.; Kiessling, F.; Weber, C.; et al. Endothelial Hypoxia-Inducible Factor-1α Promotes Atherosclerosis and Monocyte Recruitment by Upregulating MicroRNA-19a. Hypertension 2015, 66, 1220–1226. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [Green Version]
- Comprehensive Physiology, 1st ed.; Terjung, R. (Ed.) Wiley: Hoboken, NJ, USA, 2011; ISBN 978-0-470-65071-4. [Google Scholar]
- Lin, S.; Li, X.; Zhang, J.; Zhang, Y. Omentin-1: Protective impact on ischemic stroke via ameliorating atherosclerosis. Clin. Chim. Acta 2021, 517, 31–40. [Google Scholar] [CrossRef]
- Askin, L.; Duman, H.; Ozyıldız, A.; Tanriverdi, O.; Turkmen, S. Association between Omentin-1 and Coronary Artery Disease: Pathogenesis and Clinical Research. Curr. Cardiol. Rev. 2020, 16, 198–201. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, M.; Wang, Z.; Li, X.; Zhu, Y.; Li, Y. Omentin-1 ameliorates the attachment of the leukocyte THP-1 cells to HUVECs by targeting the transcriptional factor KLF2. Biochem. Biophys. Res. Commun. 2018, 498, 152–156. [Google Scholar] [CrossRef]
- Sun, Z.; Han, Y.; Song, S.; Chen, T.; Han, Y.; Liu, Y. Activation of GPR81 by lactate inhibits oscillatory shear stress-induced endothelial inflammation by activating the expression of KLF2. IUBMB Life 2019, 71, 2010–2019. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Shen, Q. IRF2BP2 prevents ox-LDL-induced inflammation and EMT in endothelial cells via regulation of KLF2. Exp. Ther. Med. 2021, 21, 481. [Google Scholar] [CrossRef] [PubMed]
- Angolano, C.; Kaczmarek, E.; Essayagh, S.; Daniel, S.; Choi, L.Y.; Tung, B.; Sauvage, G.; Lee, A.; Kipper, F.C.; Arvelo, M.B.; et al. A20/TNFAIP3 Increases ENOS Expression in an ERK5/KLF2-Dependent Manner to Support Endothelial Cell Health in the Face of Inflammation. Front. Cardiovasc. Med. 2021, 8, 651230. [Google Scholar] [CrossRef] [PubMed]
- Saum, K.; Campos, B.; Celdran-Bonafonte, D.; Nayak, L.; Sangwung, P.; Thakar, C.; Roy-Chaudhury, P.; OwensIII, A.P. Uremic Advanced Glycation End Products and Protein-Bound Solutes Induce Endothelial Dysfunction Through Suppression of Krüppel-Like Factor 2. J. Am. Heart Assoc. 2018, 7, e007566. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, T.; Liu, J.; Chen, X.; Zhang, L.; Pi, J.; Sun, H.; Li, L.; Bauer, R.; Wang, H.; Yu, Z.; et al. Endothelial Foxp1 Suppresses Atherosclerosis via Modulation of Nlrp3 Inflammasome Activation. Circ. Res. 2019, 125, 590–605. [Google Scholar] [CrossRef]
- Wolf, A.J.; Reyes, C.N.; Liang, W.; Becker, C.; Shimada, K.; Wheeler, M.L.; Cho, H.C.; Popescu, N.; Coggeshall, K.M.; Arditi, M.; et al. Hexokinase Is an Innate Immune Receptor for the Detection of Bacterial Peptidoglycan. Cell 2016, 166, 624–636. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Zhu, Y.; Wang, X.-D.; Luo, E.-F.; Li, Y.-P.; Wang, B.-L.; Chen, Y.-F. BDNF corrects NLRP3 inflammasome-induced pyroptosis and glucose metabolism reprogramming through KLF2/HK1 pathway in vascular endothelial cells. Cell. Signal. 2020, 78, 109843. [Google Scholar] [CrossRef]
- Rane, M.J.; Zhao, Y.; Cai, L. Krϋppel-like factors (KLFs) in renal physiology and disease. eBioMedicine 2019, 40, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y. Potential roles of Kruppel-like factors in mediating adverse vascular effects of nanomaterials: A review. J. Appl. Toxicol. 2021, 42, 4–16. [Google Scholar] [CrossRef]
- Xu, Z.; Yoshida, T.; Wu, L.; Maiti, D.; Cebotaru, L.; Duh, E.J. Transcription Factor MEF2C Suppresses Endothelial Cell Inflammation via Regulation of NF-κB and KLF2. J. Cell. Physiol. 2015, 230, 1310–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, M.; Hansen, S.; Leefmann, J.; Guan, K. Repurposing Antidiabetic Drugs for Cardiovascular Disease. Front. Physiol. 2020, 11, 568632. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, F.; Grant, P.J.; Aboyans, V.; Bailey, C.J.; Ceriello, A.; Delgado, V.; Federici, M.; Filippatos, G.; Grobbee, D.E.; Hansen, T.B.; et al. 2019 ESC Guidelines on diabetes, pre-diabetes, and cardiovascular diseases developed in collaboration with the EASD. Eur. Heart J. 2020, 41, 255–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kashef, D.; Abdelrahman, R.S. Montelukast ameliorates Concanavalin A-induced autoimmune hepatitis in mice via inhibiting TNF-α/JNK signaling pathway. Toxicol. Appl. Pharmacol. 2020, 393, 114931. [Google Scholar] [CrossRef] [PubMed]
- Di, X.; Tang, X.; Di, X. Montelukast inhibits oxidized low-density lipoproteins (ox-LDL) induced vascular endothelial attachment: An implication for the treatment of atherosclerosis. Biochem. Biophys. Res. Commun. 2017, 486, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Mistrova, E.; Kruzliak, P.; Chottova Dvorakova, M.C. Role of substance P in the cardiovascular system. Neuropeptides 2016, 58, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Chen, K.; Zhu, Y.; Wang, H.; Chen, Z.; Yong, X.; Yin, H.; Chen, J.; Lai, K.; Liu, Y. The neurokinin-1 receptor antagonist aprepitant ameliorates oxidized LDL-induced endothelial dysfunction via KLF2. Mol. Immunol. 2018, 106, 29–35. [Google Scholar] [CrossRef]
- Liu, H.; Mao, P.; Wang, J.; Wang, T.; Xie, C.-H. Azilsartan, an angiotensin II type 1 receptor blocker, attenuates tert-butyl hydroperoxide-induced endothelial cell injury through inhibition of mitochondrial dysfunction and anti-inflammatory activity. Neurochem. Int. 2016, 94, 48–56. [Google Scholar] [CrossRef]
- Hardin, M.D.; Jacobs, T.F. Azilsartan. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Li, W.; Wang, C.; Zhang, D.; Zeng, K.; Xiao, S.; Chen, F.; Luo, J. Azilsartan ameliorates ox-LDL-induced endothelial dysfunction via promoting the expression of KLF2. Aging 2021, 13, 12996–13005. [Google Scholar] [CrossRef]
- Annus, A.; Vecsei, L. Kynurenine System and Multiple Sclerosis, Pathomechanism and Drug Targets with An Emphasis on Laquinimod. Curr. Drug Targets 2018, 19, 805–814. [Google Scholar] [CrossRef]
- Jiang, T.; Zhang, W.; Wang, Z. Laquinimod Protects Against TNF-α-Induced Attachment of Monocytes to Human Aortic Endothelial Cells (HAECs) by Increasing the Expression of KLF2. Drug Des. Dev. Ther. 2020, 14, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Roche-Molina, M.; Hardwick, B.; Sanchez-Ramos, C.; Sanz-Rosa, D.; Gewert, D.; Cruz, F.M.; Gonzalez-Guerra, A.; Andrés, V.; Palma, J.A.; Ibanez, B.; et al. The pharmaceutical solvent N-methyl-2-pyrollidone (NMP) attenuates inflammation through Krüppel-like factor 2 activation to reduce atherogenesis. Sci. Rep. 2020, 10, 11636. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Mahendran, A.; Yao, Y.; Ngo, L.; Venta-Perez, G.; Choy, M.L.; Kim, N.; Ham, W.S.; Breslow, R.; Marks, P.A. Development of a histone deacetylase 6 inhibitor and its biological effects. Proc. Natl. Acad. Sci. USA 2013, 110, 15704–15709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Xu, S.; Liu, P.; Koroleva, M.; Zhang, S.; Si, S.; Jin, Z.G. Suberanilohydroxamic Acid as a Pharmacological Kruppel-Like Factor 2 Activator That Represses Vascular Inflammation and Atherosclerosis. J. Am. Hear. Assoc. 2017, 6, e007134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauck, M.A.; Meier, J.J.; Cavender, M.A.; El Aziz, M.A.; Drucker, D.J. Cardiovascular Actions and Clinical Outcomes With Glucagon-Like Peptide-1 Receptor Agonists and Dipeptidyl Peptidase-4 Inhibitors. Circulation 2017, 136, 849–870. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Li, Y.; Ou, D.; Yang, Q. The GLP-1 receptor agonist liraglutide protects against oxidized LDL-induced endothelial inflammation and dysfunction via KLF2. IUBMB Life 2019, 71, 1347–1354. [Google Scholar] [CrossRef]
- Zhi, D.; Zhang, M.; Lin, J.; Liu, P.; Duan, M. GPR120 Ameliorates Apoptosis and Inhibits the Production of Inflammatory Cytokines in Renal Tubular Epithelial Cells. Inflammation 2020, 44, 493–505. [Google Scholar] [CrossRef]
- Croze, M.L.; Guillaume, A.; Ethier, M.; Fergusson, G.; Tremblay, C.; Campbell, S.A.; Maachi, H.; Ghislain, J.; Poitout, V. Combined Deletion of Free Fatty-Acid Receptors 1 and 4 Minimally Impacts Glucose Homeostasis in Mice. Endocrinology 2021, 162, bqab002. [Google Scholar] [CrossRef]
- Jiang, T.; Jiang, D.; You, D.; Zhang, L.; Liu, L.; Zhao, Q. Agonism of GPR120 prevents ox-LDL-induced attachment of monocytes to endothelial cells. Chem. Interact. 2019, 316, 108916. [Google Scholar] [CrossRef]
- Wu, H.; Feng, K.; Zhang, C.; Zhang, H.; Zhang, J.; Hua, Y.; Dong, Z.; Zhu, Y.; Yang, S.; Ma, C. Metformin attenuates atherosclerosis and plaque vulnerability by upregulating KLF2-mediated autophagy in apoE−/- mice. Biochem. Biophys. Res. Commun. 2021, 557, 334–341. [Google Scholar] [CrossRef]
- Vion, A.-C.; Kheloufi, M.; Hammoutene, A.; Poisson, J.; Lasselin, J.; Devue, C.; Pic, I.; Dupont, N.; Busse, J.; Stark, K.; et al. Autophagy is required for endothelial cell alignment and atheroprotection under physiological blood flow. Proc. Natl. Acad. Sci. USA 2017, 114, E8675–E8684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, M.; Spinosa, M.; Zehner, Z.E.; Upchurch, G.R.; Ailawadi, G. Klf4, Klf2, and Zfp148 activate autophagy-related genes in smooth muscle cells during aortic aneurysm formation. Physiol. Rep. 2019, 7, e14058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Fonseca, M.; Galan, M.; Martinez-Lopez, D.; Cañes, L.; Roldan-Montero, R.; Alonso, J.; Reyero-Postigo, T.; Orriols, M.; Mendez-Barbero, N.; Sirvent, M.; et al. Fisiopatología del aneurisma de aorta abdominal: Biomarcadores y nuevas dianas terapéuticas. Clínica Investig. Arterioscler. 2019, 31, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Laha, D.; Deb, M.; Das, H. KLF2 (kruppel-like factor 2 [lung]) regulates osteoclastogenesis by modulating autophagy. Autophagy 2019, 15, 2063–2075. [Google Scholar] [CrossRef] [PubMed]
- Maity, J.; Deb, M.; Greene, C.; Das, H. KLF2 regulates dental pulp-derived stem cell differentiation through the induction of mitophagy and altering mitochondrial metabolism. Redox Biol. 2020, 36, 101622. [Google Scholar] [CrossRef]
- Sharif, H.; Akash, M.S.H.; Rehman, K.; Irshad, K.; Imran, I. Pathophysiology of atherosclerosis: Association of risk factors and treatment strategies using plant-based bioactive compounds. J. Food Biochem. 2020, 44, e13449. [Google Scholar] [CrossRef]
- Chu, H.; Li, H.; Guan, X.; Yan, H.; Zhang, X.; Cui, X.; Li, X.; Cheng, M. Resveratrol protects late endothelial progenitor cells from TNF-α-induced inflammatory damage by upregulating Kr�ppel-like factor-2. Mol. Med. Rep. 2018, 17, 5708–5715. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-M.; Li, H.; Cho, M.-H.; Baek, S.-H.; Lee, C.-H.; Park, H.-Y.; Jeong, T.-S. Soy-Leaf Extract Exerts Atheroprotective Effects via Modulation of Krüppel-Like Factor 2 and Adhesion Molecules. Int. J. Mol. Sci. 2017, 18, 373. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Liu, P.; Xu, S.; Koroleva, M.; Zhang, S.; Si, S.; Jin, Z.G. Tannic acid as a plant-derived polyphenol exerts vasoprotection via enhancing KLF2 expression in endothelial cells. Sci. Rep. 2017, 7, 6686. [Google Scholar] [CrossRef]
- Zhou, Y.-X.; Zhang, H.; Peng, C. Puerarin: A Review of Pharmacological Effects. Phytother. Res. 2013, 28, 961–975. [Google Scholar] [CrossRef]
- Deng, Y.; Lei, T.; Li, H.; Mo, X.; Wang, Z.; Ou, H. ERK5/KLF2 activation is involved in the reducing effects of puerarin on monocyte adhesion to endothelial cells and atherosclerotic lesion in apolipoprotein E-deficient mice. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2018, 1864, 2590–2599. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, A.; Ruan, M.; Jin, Z.; Lu, J.; Wang, Q.; Lu, C. Dehydrocostus lactone suppresses ox-LDL-induced attachment of monocytes to endothelial cells. Am. J. Transl. Res. 2019, 11, 6159–6169. [Google Scholar] [PubMed]
- Chiu, Y.; Tsai, W.-C.; Wu, C.-H.; Wu, C.-H.; Cheng, C.-C.; Lin, W.-S.; Tsai, T.-N.; Wu, L.-S. Ginkgo biloba Induces Thrombomodulin Expression and Tissue-Type Plasminogen Activator Secretion via the Activation of Krüppel-Like Factor 2 within Endothelial Cells. Am. J. Chin. Med. 2020, 48, 357–372. [Google Scholar] [CrossRef] [PubMed]
- He, P.-P.; Jiang, T.; OuYang, X.-P.; Liang, Y.-Q.; Zou, J.-Q.; Wang, Y.; Shen, Q.-Q.; Liao, L.; Zheng, X.-L. Lipoprotein lipase: Biosynthesis, regulatory factors, and its role in atherosclerosis and other diseases. Clin. Chim. Acta 2018, 480, 126–137. [Google Scholar] [CrossRef] [PubMed]
- He, L.-H.; Gao, J.-H.; Yu, X.-H.; Wen, F.-J.; Luo, J.-J.; Qin, Y.-S.; Chen, M.-X.; Zhang, D.-W.; Wang, Z.-B.; Tang, C.-K. Artesunate inhibits atherosclerosis by upregulating vascular smooth muscle cells-derived LPL expression via the KLF2/NRF2/TCF7L2 pathway. Eur. J. Pharmacol. 2020, 884, 173408. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Feng, H.; Li, Z.-W.; Tang, K.-X.; Gao, H.-Q.; Wang, W.-L.; Cui, X.-P.; Li, X.-L. Low-dose phloretin alleviates diabetic atherosclerosis through endothelial KLF2 restoration. Biosci. Biotechnol. Biochem. 2019, 84, 815–823. [Google Scholar] [CrossRef]
- Grymel, M.; Zawojak, M.; Adamek, J. Triphenylphosphonium Analogues of Betulin and Betulinic Acid with Biological Activity: A Comprehensive Review. J. Nat. Prod. 2019, 82, 1719–1730. [Google Scholar] [CrossRef]
- Lee, G.H.; Park, J.S.; Jin, S.W.; Pham, T.H.; Thai, T.N.; Kim, J.Y.; Kim, C.Y.; Choi, J.H.; Han, E.H.; Jeong, H.G. Betulinic Acid Induces eNOS Expression via the AMPK-Dependent KLF2 Signaling Pathway. J. Agric. Food Chem. 2020, 68, 14523–14530. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Samarghandian, S.; Bafandeh, F. The Cardiovascular Protective Effects of Chrysin: A Narrative Review on Experimental Researches. Cardiovasc. Hematol. Agents Med. Chem. 2019, 17, 17–27. [Google Scholar] [CrossRef]
- Lin, C.-M.; Wang, B.-W.; Pan, C.-M.; Fang, W.-J.; Chua, S.-K.; Cheng, W.-P.; Shyu, K.-G. Chrysin boosts KLF2 expression through suppression of endothelial cell-derived exosomal microRNA-92a in the model of atheroprotection. Eur. J. Nutr. 2021, 60, 4345–4355. [Google Scholar] [CrossRef]
- Fang, Y.; Davies, P.F. Site-Specific MicroRNA-92a Regulation of Krüppel-Like Factors 4 and 2 in Atherosusceptible Endothelium. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 979–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dabravolski, S.A.; Sukhorukov, V.N.; Kalmykov, V.A.; Grechko, A.V.; Shakhpazyan, N.K.; Orekhov, A.N. The Role of KLF2 in the Regulation of Atherosclerosis Development and Potential Use of KLF2-Targeted Therapy. Biomedicines 2022, 10, 254. https://doi.org/10.3390/biomedicines10020254
Dabravolski SA, Sukhorukov VN, Kalmykov VA, Grechko AV, Shakhpazyan NK, Orekhov AN. The Role of KLF2 in the Regulation of Atherosclerosis Development and Potential Use of KLF2-Targeted Therapy. Biomedicines. 2022; 10(2):254. https://doi.org/10.3390/biomedicines10020254
Chicago/Turabian StyleDabravolski, Siarhei A., Vasily N. Sukhorukov, Vladislav A. Kalmykov, Andrey V. Grechko, Nikolay K. Shakhpazyan, and Alexander N. Orekhov. 2022. "The Role of KLF2 in the Regulation of Atherosclerosis Development and Potential Use of KLF2-Targeted Therapy" Biomedicines 10, no. 2: 254. https://doi.org/10.3390/biomedicines10020254