
Cutaneous and Developmental Effects of CARD14 Overexpression in Zebrafish
Abstract
:
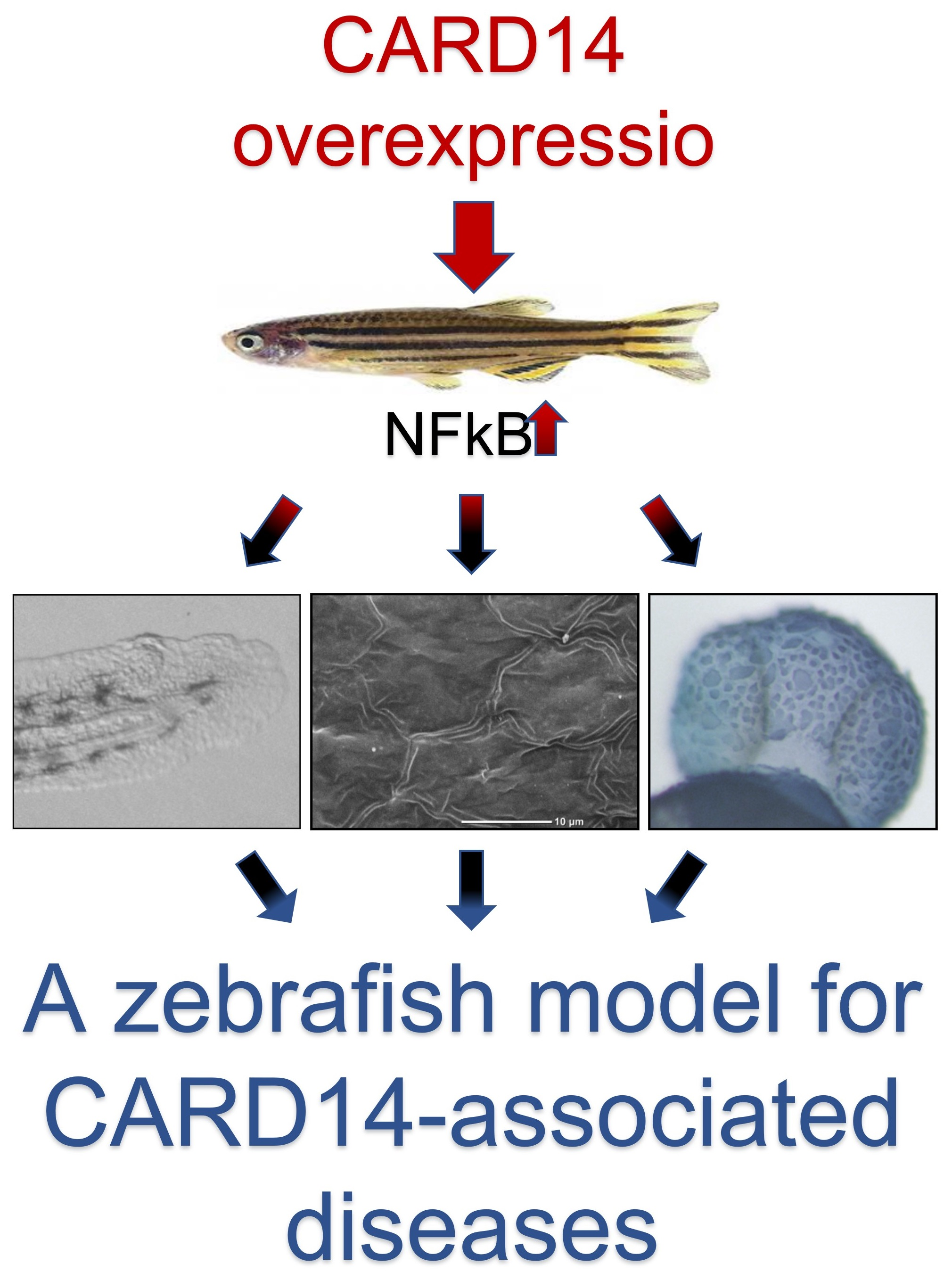{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. DNA and RNA Injection to Zebrafish Embryos
2.2. Whole Mount In Situ Hybridization (WISH)
2.3. Scanning Electron Microscopy (SEM)
2.4. Histology
2.5. RNA Purification
2.6. Real Time PCR
3. Results
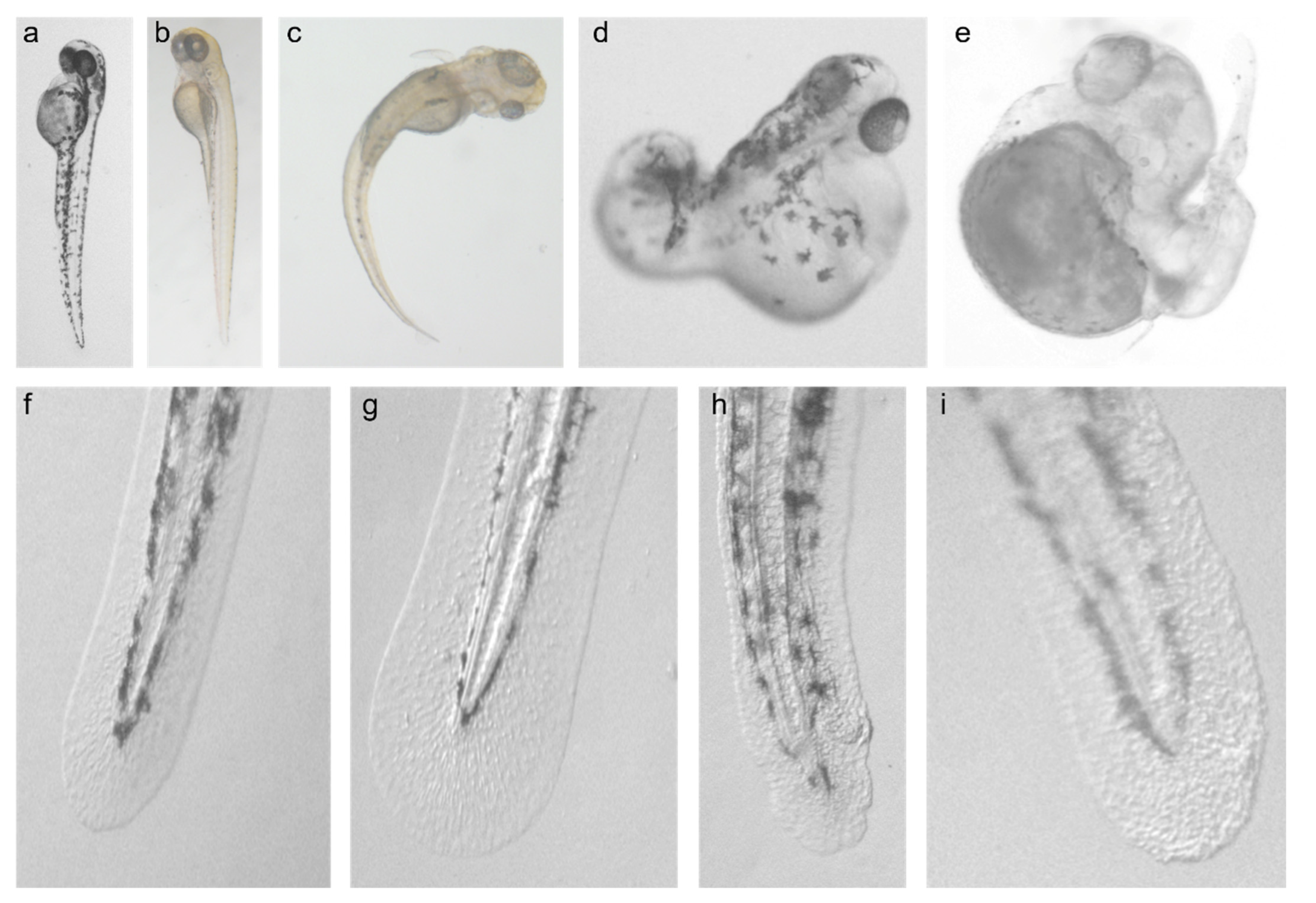
3.1. Effect of CARD14 Overexpression on Zebrafish Larvae Development
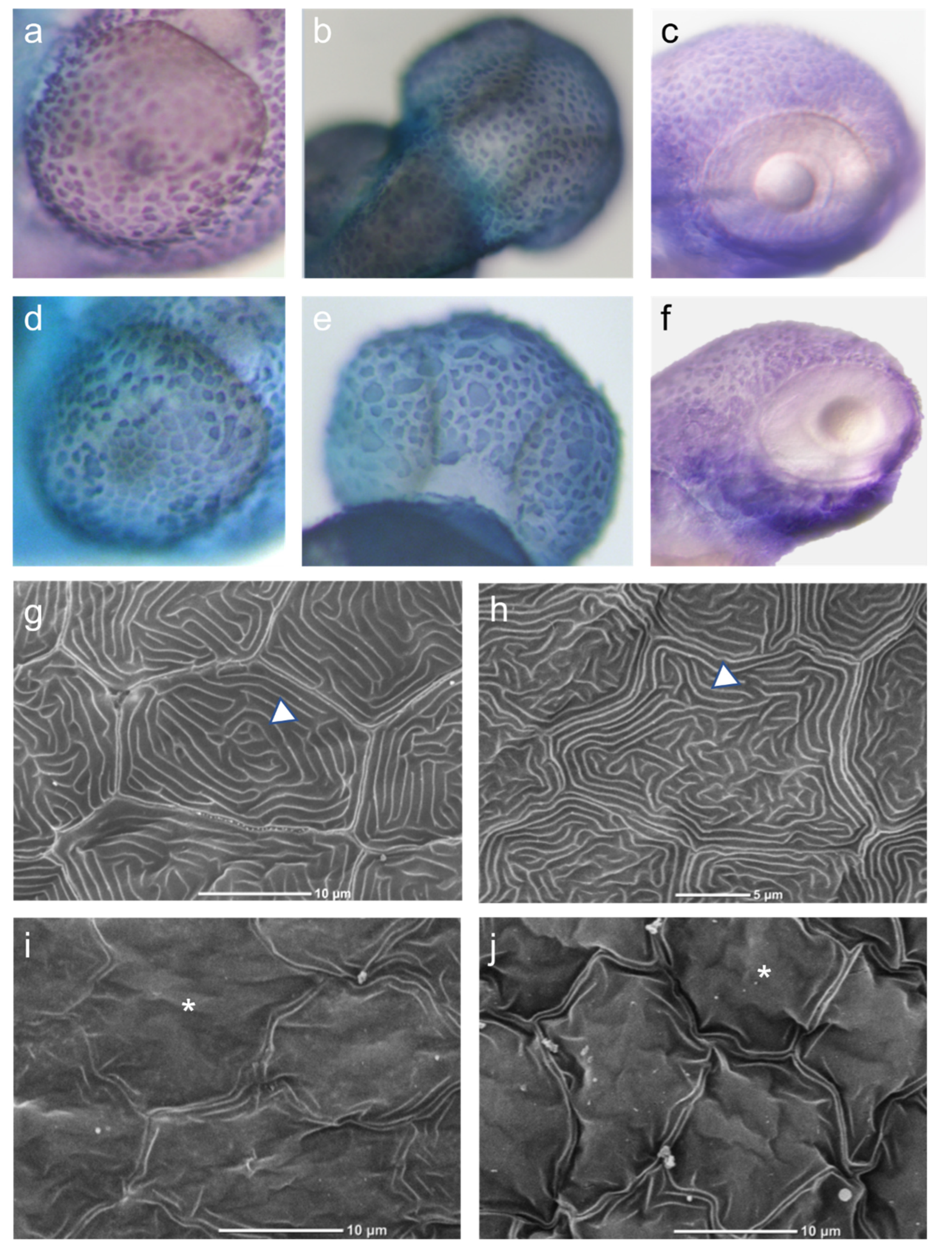
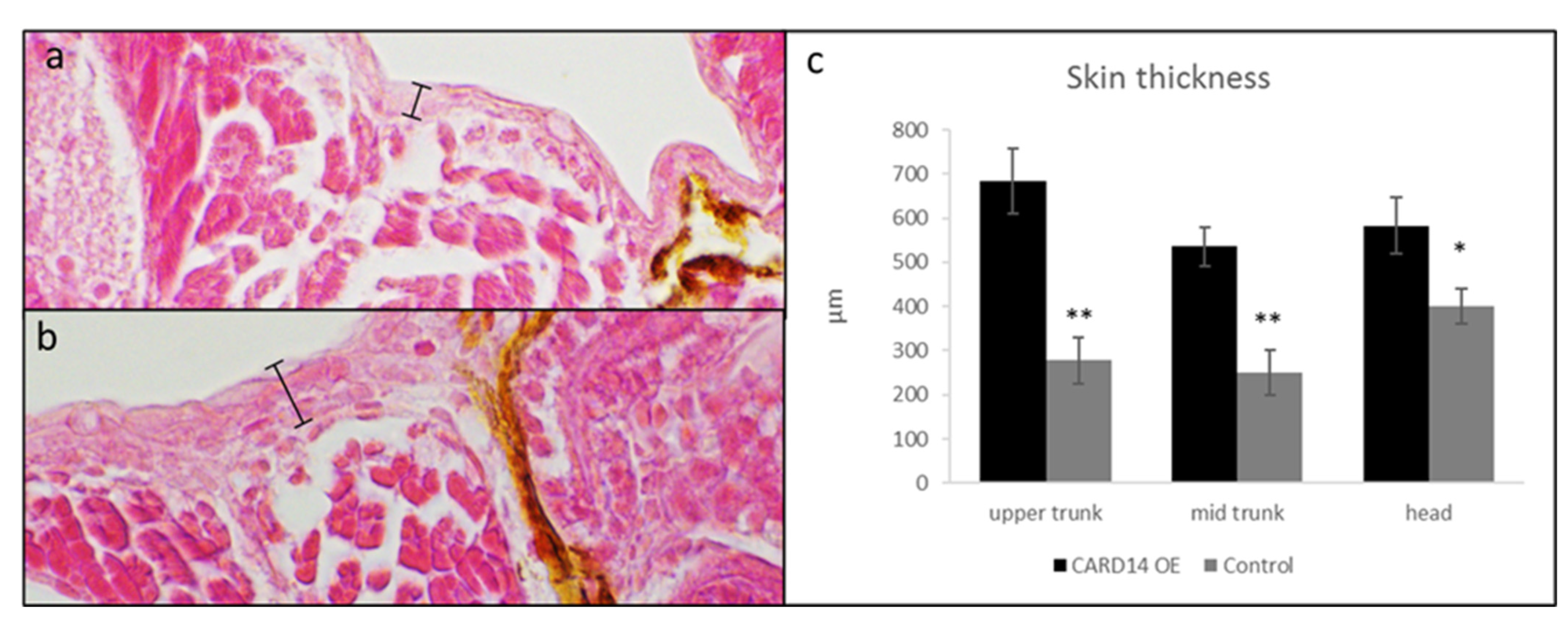
3.2. Effect of CARD14 Overexpression on Zebrafish Larvae Cutaneous Morphology
3.3. Effect of CARD14 Overexpression on Keratinocyte Morphology
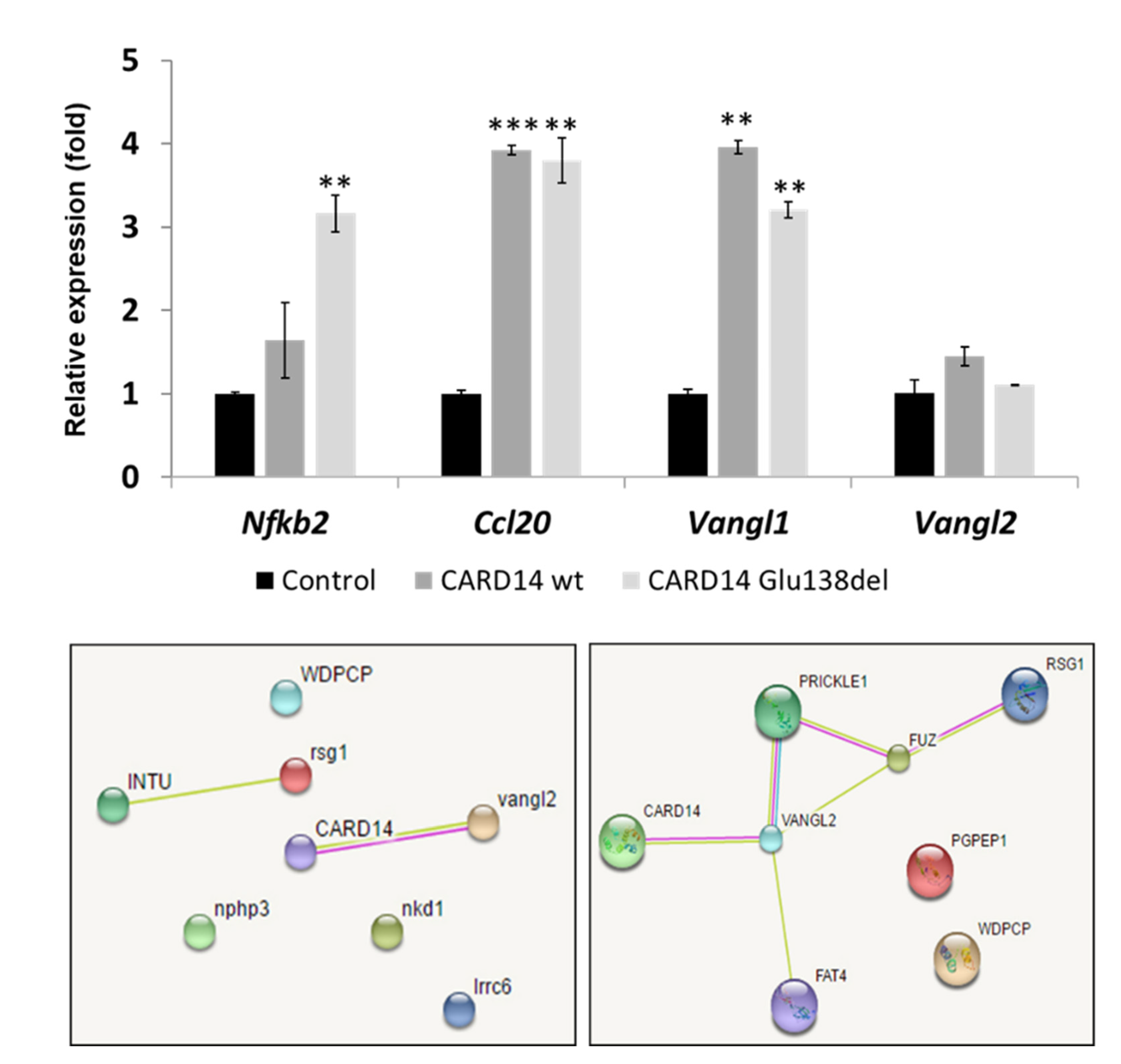
3.4. Over Expression of CARD14 Affects Nfκb and Planar Cell Polarity Signaling Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, C.V.; Farah, R.S.; Maguiness, S.M.; Miller, D.D. Follicular Psoriasis: Differentiation from Pityriasis Rubra Pilaris-An Illustrative Case and Review of the Literature. Pediatr. Dermatol. 2017, 34, e65–e68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoi, L.C.; Spain, S.L.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Knight, J.; Tejasvi, T.; Kang, H.M.; Allen, M.H.; Lambert, S.; et al. Enhanced meta-analysis and replication studies identify five new psoriasis susceptibility loci. Nat. Commun. 2015, 6, 7001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderhooft, S.L.; Francis, J.S.; Holbrook, K.A.; Dale, B.A.; Fleckman, P. Familial pityriasis rubra pilaris. Arch. Dermatol. 1995, 131, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Fuchs-Telem, D.; Sarig, O.; van Steensel, M.A.; Isakov, O.; Israeli, S.; Nousbeck, J.; Richard, K.; Winnepenninckx, V.; Vernooij, M.; Shomron, N.; et al. Familial pityriasis rubra pilaris is caused by mutations in CARD14. Am. J. Hum. Genet. 2012, 91, 163–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Duan, S.; Helms, C.A.; Nair, R.P.; Duffin, K.C.; Stuart, P.E.; Goldgar, D.; Hayashi, G.; et al. Rare and common variants in CARD14, encoding an epidermal regulator of NF-kappaB, in psoriasis. Am. J. Hum. Genet. 2012, 90, 796–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helms, C.; Cao, L.; Krueger, J.G.; Wijsman, E.M.; Chamian, F.; Gordon, D.; Heffernan, M.; Daw, J.A.; Robarge, J.; Ott, J.; et al. A putative RUNX1 binding site variant between SLC9A3R1 and NAT9 is associated with susceptibility to psoriasis. Nat. Genet. 2003, 35, 349–356. [Google Scholar] [CrossRef]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Pierson, K.C.; Yang, C.F.; Joyce, C.E.; Ryan, C.; Duan, S.; Helms, C.A.; Liu, Y.; et al. PSORS2 is due to mutations in CARD14. Am. J. Hum. Genet. 2012, 90, 784–795. [Google Scholar] [CrossRef] [Green Version]
- Israel, L.; Mellett, M. Clinical and Genetic Heterogeneity of CARD14 Mutations in Psoriatic Skin Disease. Front. Immunol. 2018, 9, 2239. [Google Scholar] [CrossRef] [Green Version]
- Mellett, M. Regulation and dysregulation of CARD14 signalling and its physiological consequences in inflammatory skin disease. Cell. Immunol. 2020, 354, 104147. [Google Scholar] [CrossRef]
- DeVore, S.B.; Stevens, M.L.; He, H.; Biagini, J.M.; Kroner, J.W.; Martin, L.J.; Khurana Hershey, G.K. Novel role for caspase recruitment domain family member 14 and its genetic variant rs11652075 in skin filaggrin homeostasis. J. Allergy Clin. Immunol. 2022, 149, 708–717. [Google Scholar] [CrossRef]
- Murase, Y.; Takeichi, T.; Akiyama, M. Aberrant CARD14 function might cause defective barrier formation. J. Allergy Clin. Immunol. 2019, 143, 1656–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peled, A.; Sarig, O.; Sun, G.; Samuelov, L.; Ma, C.A.; Zhang, Y.; Dimaggio, T.; Nelson, C.G.; Stone, K.D.; Freeman, A.F.; et al. Loss-of-function mutations in caspase recruitment domain-containing protein 14 (CARD14) are associated with a severe variant of atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, I.; Vito, P.; Stilo, R. The three CARMA sisters: So different, so similar: A portrait of the three CARMA proteins and their involvement in human disorders. J. Cell. Physiol. 2014, 229, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Lin, X. Regulation of NF-kappaB by the CARD proteins. Immunol. Rev. 2012, 246, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Afonina, I.S.; Van Nuffel, E.; Baudelet, G.; Driege, Y.; Kreike, M.; Staal, J.; Beyaert, R. The paracaspase MALT1 mediates CARD14-induced signaling in keratinocytes. EMBO Rep. 2016, 17, 914–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.X.; Chen, W.Q.; Nie, P. Structure and expression pattern of teleost caspase recruitment domain (CARD) containing proteins that are potentially involved in NF-kappaB signalling. Dev. Comp. Immunol. 2010, 34, 1–13. [Google Scholar] [CrossRef]
- Li, Q.; Jin Chung, H.; Ross, N.; Keller, M.; Rews, J.; Kingman, J.; Sarig, O.; Fuchs-Telem, D.; Sprecher, E.; Uitto, J. Analysis of CARD14 Polymorphisms in Pityriasis Rubra Pilaris: Activation of NF-kappaB. J. Investig. Dermatol. 2015, 135, 1905–1908. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Wang, T.; Li, S.J.; Fan, Y.M.; Shi, G.; Zhu, K.J. CARD14 gene polymorphism c.C2458T (p.Arg820Trp) is associated with clinical features of psoriasis vulgaris in a Chinese cohort. J. Dermatol. 2016, 43, 294–297. [Google Scholar] [CrossRef]
- Ammar, M.; Jordan, C.T.; Cao, L.; Lim, E.; Bouchlaka Souissi, C.; Jrad, A.; Omrane, I.; Kouidhi, S.; Zaraa, I.; Anbunathan, H.; et al. CARD14 alterations in Tunisian patients with psoriasis and further characterization in European cohorts. Br. J. Dermatol. 2016, 174, 330–337. [Google Scholar] [CrossRef]
- Mossner, R.; Frambach, Y.; Wilsmann-Theis, D.; Lohr, S.; Jacobi, A.; Weyergraf, A.; Muller, M.; Philipp, S.; Renner, R.; Traupe, H.; et al. Palmoplantar Pustular Psoriasis Is Associated with Missense Variants in CARD14, but Not with Loss-of-Function Mutations in IL36RN in European Patients. J. Investig. Dermatol. 2015, 135, 2538–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berki, D.M.; Liu, L.; Choon, S.E.; Burden, A.D.; Griffiths, C.E.; Navarini, A.A.; Tan, E.S.; Irvine, A.D.; Ranki, A.; Ogo, T.; et al. Activating CARD14 Mutations Are Associated with Generalized Pustular Psoriasis but Rarely Account for Familial Recurrence in Psoriasis Vulgaris. J. Investig. Dermatol. 2015, 135, 2964–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, K.; Muto, M.; Akiyama, M. CARD14 c.526G>C (p.Asp176His) is a significant risk factor for generalized pustular psoriasis with psoriasis vulgaris in the Japanese cohort. J. Investig. Dermatol. 2014, 134, 1755–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, P.; Zhang, Q.; Chen, M.; Fu, X.; Wang, C.; Wang, Z.; Yu, G.; Yu, Y.; Li, X.; Sun, Y.; et al. Variant analysis of CARD14 in a Chinese Han population with psoriasis vulgaris and generalized pustular psoriasis. J. Investig. Dermatol. 2014, 134, 2994–2996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeichi, T.; Sugiura, K.; Nomura, T.; Sakamoto, T.; Ogawa, Y.; Oiso, N.; Futei, Y.; Fujisaki, A.; Koizumi, A.; Aoyama, Y.; et al. Pityriasis Rubra Pilaris Type V as an Autoinflammatory Disease by CARD14 Mutations. JAMA Dermatol. 2017, 153, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Dainichi, T.; Fujisawa, A.; Nakano, H.; Sawamura, D.; Kabashima, K. CARD14 Glu138 mutation in familial pityriasis rubra pilaris does not warrant differentiation from familial psoriasis. J. Dermatol. 2016, 43, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.B.; Chen, P.L.; Chen, Y.T.; Tsai, T.F. Genetic analysis of CARD14 in non-familial pityriasis rubra pilaris: A case series. Acta Derm.-Venereol. 2014, 94, 587–588. [Google Scholar] [CrossRef] [Green Version]
- Eytan, O.; Qiaoli, L.; Nousbeck, J.; van Steensel, M.A.; Burger, B.; Hohl, D.; Taieb, A.; Prey, S.; Bachmann, D.; Avitan-Hersh, E.; et al. Increased epidermal expression and absence of mutations in CARD14 in a series of patients with sporadic pityriasis rubra pilaris. Br. J. Dermatol. 2014, 170, 1196–1198. [Google Scholar] [CrossRef]
- Eytan, O.; Sarig, O.; Sprecher, E.; van Steensel, M.A. Clinical response to ustekinumab in familial pityriasis rubra pilaris caused by a novel mutation in CARD14. Br. J. Dermatol. 2014, 171, 420–422. [Google Scholar] [CrossRef]
- Van Nuffel, E.; Schmitt, A.; Afonina, I.S.; Schulze-Osthoff, K.; Beyaert, R.; Hailfinger, S. CARD14-Mediated Activation of Paracaspase MALT1 in Keratinocytes: Implications for Psoriasis. J. Investig. Dermatol. 2016, 137, 569–575. [Google Scholar] [CrossRef]
- Wu, K.C.; Reynolds, N.J. CARD14 mutations may predict response to antitumour necrosis factor-alpha therapy in psoriasis: A potential further step towards personalized medicine. Br. J. Dermatol. 2016, 175, 17–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, A.E.; Driever, W.; Kimelman, D. Psoriasis regulates epidermal development in zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2008, 237, 1153–1164. [Google Scholar] [CrossRef]
- Lam, P.Y.; Mangos, S.; Green, J.M.; Reiser, J.; Huttenlocher, A. In vivo imaging and characterization of actin microridges. PLoS ONE 2015, 10, e0115639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperry, D.G.; Wassersug, R.J. A proposed function for microridges on epithelial cells. Anat. Rec. 1976, 185, 253–257. [Google Scholar] [CrossRef]
- Raman, R.; Damle, I.; Rote, R.; Banerjee, S.; Dingare, C.; Sonawane, M. aPKC regulates apical localization of Lgl to restrict elongation of microridges in developing zebrafish epidermis. Nat. Commun. 2016, 7, 11643. [Google Scholar] [CrossRef] [Green Version]
- Antic, D.; Stubbs, J.L.; Suyama, K.; Kintner, C.; Scott, M.P.; Axelrod, J.D. Planar cell polarity enables posterior localization of nodal cilia and left-right axis determination during mouse and Xenopus embryogenesis. PLoS ONE 2010, 5, e8999. [Google Scholar] [CrossRef]
- Torban, E.; Kor, C.; Gros, P. Van Gogh-like2 (Strabismus) and its role in planar cell polarity and convergent extension in vertebrates. Trends Genet. TIG 2004, 20, 570–577. [Google Scholar] [CrossRef]
- Park, M.; Moon, R.T. The planar cell-polarity gene stbm regulates cell behaviour and cell fate in vertebrate embryos. Nat. Cell Biol. 2002, 4, 20–25. [Google Scholar] [CrossRef]
- Torban, E.; Patenaude, A.M.; Leclerc, S.; Rakowiecki, S.; Gauthier, S.; Elfinger, G.; Epstein, D.J.; Gros, P. Genetic interaction between members of the Vangl family causes neural tube defects in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 3449–3454. [Google Scholar] [CrossRef] [Green Version]
- Montcouquiol, M.; Rachel, R.A.; Lanford, P.J.; Copeland, N.G.; Jenkins, N.A.; Kelley, M.W. Identification of Vangl2 and Scrb1 as planar polarity genes in mammals. Nature 2003, 423, 173–177. [Google Scholar] [CrossRef]
- Lei, Y.P.; Zhang, T.; Li, H.; Wu, B.L.; Jin, L.; Wang, H.Y. VANGL2 mutations in human cranial neural-tube defects. N. Engl. J. Med. 2010, 362, 2232–2235. [Google Scholar] [CrossRef] [PubMed]
- Kibar, Z.; Torban, E.; McDearmid, J.R.; Reynolds, A.; Berghout, J.; Mathieu, M.; Kirillova, I.; De Marco, P.; Merello, E.; Hayes, J.M.; et al. Mutations in VANGL1 associated with neural-tube defects. N. Engl. J. Med. 2007, 356, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Devenport, D.; Fuchs, E. Planar polarization in embryonic epidermis orchestrates global asymmetric morphogenesis of hair follicles. Nat. Cell Biol. 2008, 10, 1257–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, R.; Natsume, Y.; Noda, T. MAGI-3 is involved in the regulation of the JNK signaling pathway as a scaffold protein for frizzled and Ltap. Oncogene 2004, 23, 6023–6030. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, A.; McDearmid, J.R.; Lachance, S.; De Marco, P.; Merello, E.; Capra, V.; Gros, P.; Drapeau, P.; Kibar, Z. VANGL1 rare variants associated with neural tube defects affect convergent extension in zebrafish. Mech. Dev. 2010, 127, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Ostensson, M.; Monten, C.; Bacelis, J.; Gudjonsdottir, A.H.; Adamovic, S.; Ek, J.; Ascher, H.; Pollak, E.; Arnell, H.; Browaldh, L.; et al. A possible mechanism behind autoimmune disorders discovered by genome-wide linkage and association analysis in celiac disease. PLoS ONE 2013, 8, e70174. [Google Scholar] [CrossRef]
- De Luca, M.; Pellegrini, G.; Zambruno, G.; Marchisio, P.C. Role of integrins in cell adhesion and polarity in normal keratinocytes and human skin pathologies. J. Dermatol. 1994, 21, 821–828. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baniel, A.; Ziv, L.; Ben-Moshe, Z.; Sarig, O.; Mohamad, J.; Peled, A.; Rechavi, G.; Gothilf, Y.; Sprecher, E. Cutaneous and Developmental Effects of CARD14 Overexpression in Zebrafish. Biomedicines 2022, 10, 3192. https://doi.org/10.3390/biomedicines10123192
Baniel A, Ziv L, Ben-Moshe Z, Sarig O, Mohamad J, Peled A, Rechavi G, Gothilf Y, Sprecher E. Cutaneous and Developmental Effects of CARD14 Overexpression in Zebrafish. Biomedicines. 2022; 10(12):3192. https://doi.org/10.3390/biomedicines10123192
Chicago/Turabian StyleBaniel, Avital, Limor Ziv, Zohar Ben-Moshe, Ofer Sarig, Janan Mohamad, Alon Peled, Gideon Rechavi, Yoav Gothilf, and Eli Sprecher. 2022. "Cutaneous and Developmental Effects of CARD14 Overexpression in Zebrafish" Biomedicines 10, no. 12: 3192. https://doi.org/10.3390/biomedicines10123192