

Influence of Protein Carbonylation on Human Adipose Tissue Dysfunction in Obesity and Insulin Resistance
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Sample Collection
2.2. Protein Extraction
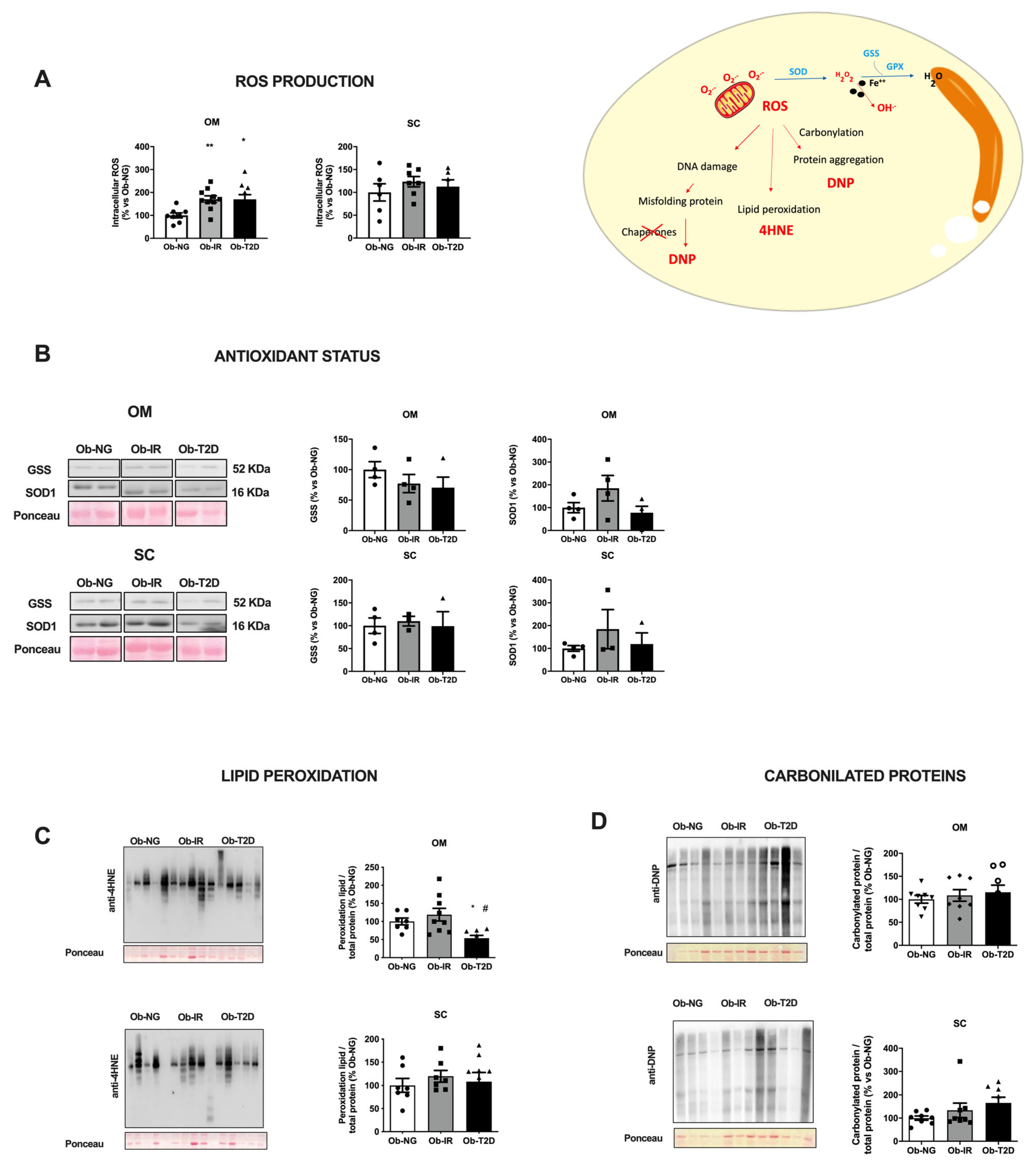
2.3. Intracellular ROS Production
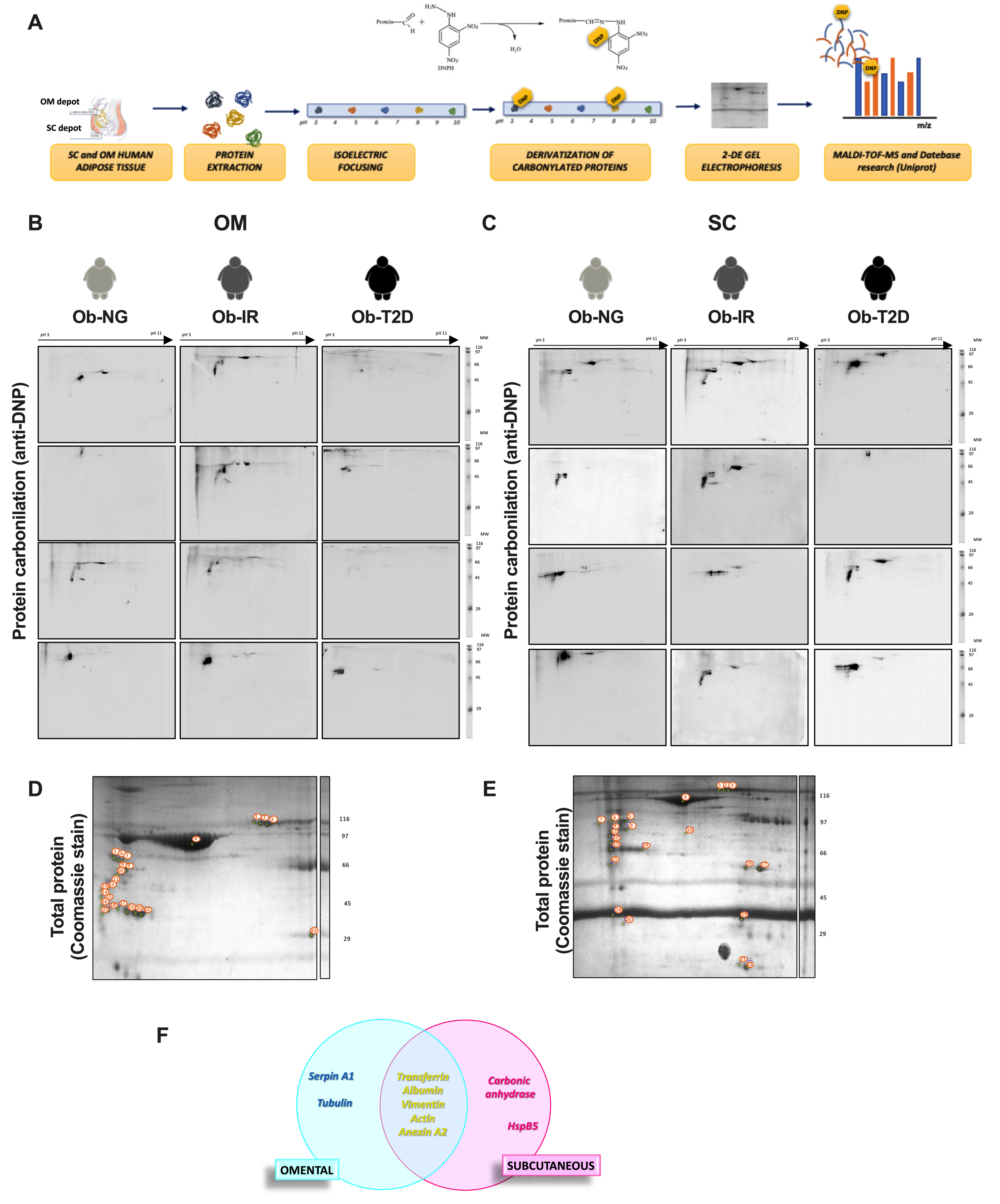
2.4. Derivatization of Carbonylated Proteins
2.5. 4-Hydroxynenal (4-HNE) Detection
2.6. Quantitative Immunoblotting
2.7. 2D-PAGE and MALDI-TOF-MS Analysis
2.8. Iron Content and Histology Staining
2.9. Culture and Treatment of 3T3-L1 Cell Line
2.10. Cell Transfection
2.11. Confocal Studies of Transferrin Receptor (TfR) Translocation in 3T3-L1 Adipocytes
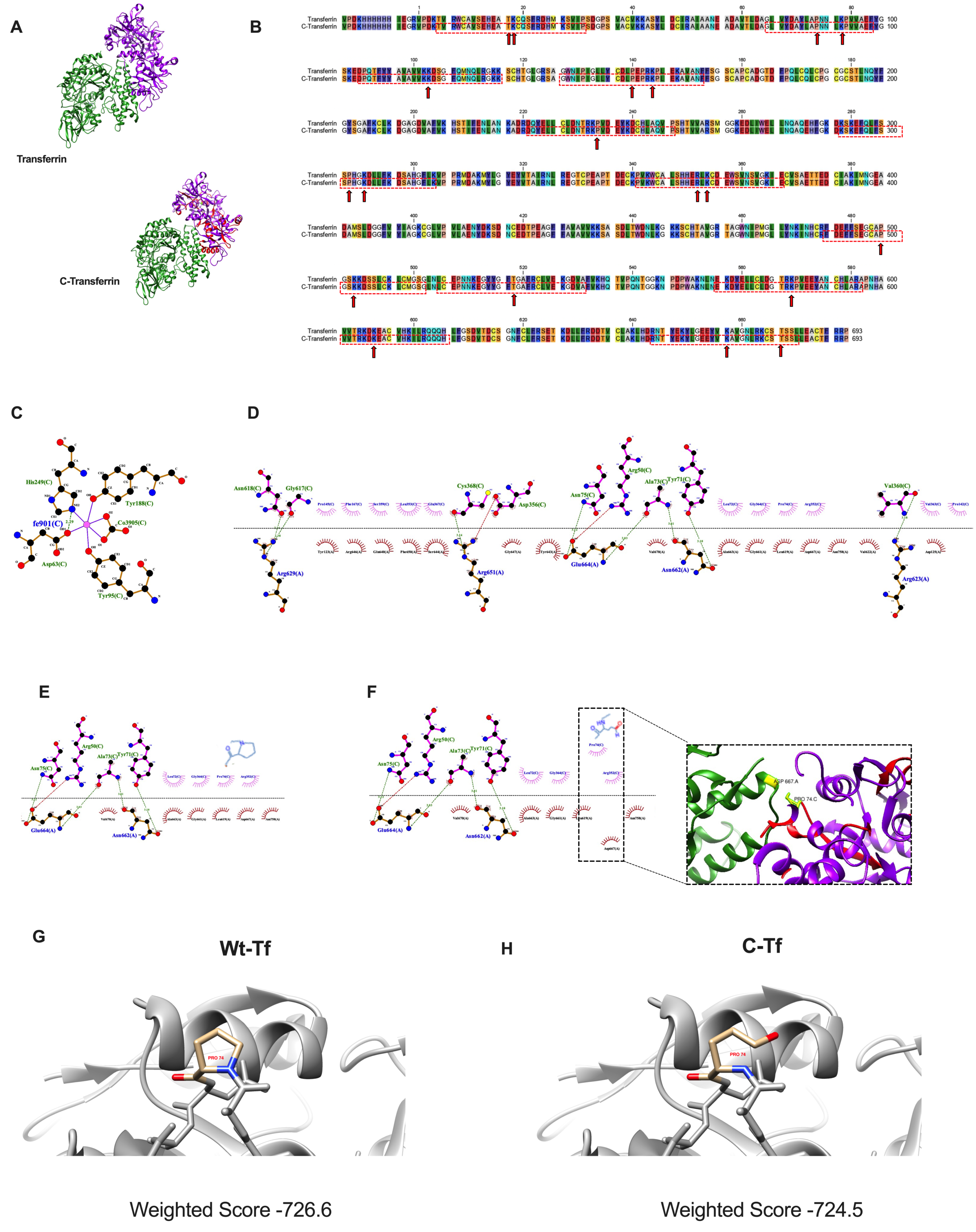
2.12. Molecular Modeling of Transferrin
2.13. Statistical Analysis
3. Results
3.1. Subject’s Characteristics
3.2. Redox Status of Adipose Tissue in Obesity-Associated Insulin Resistance
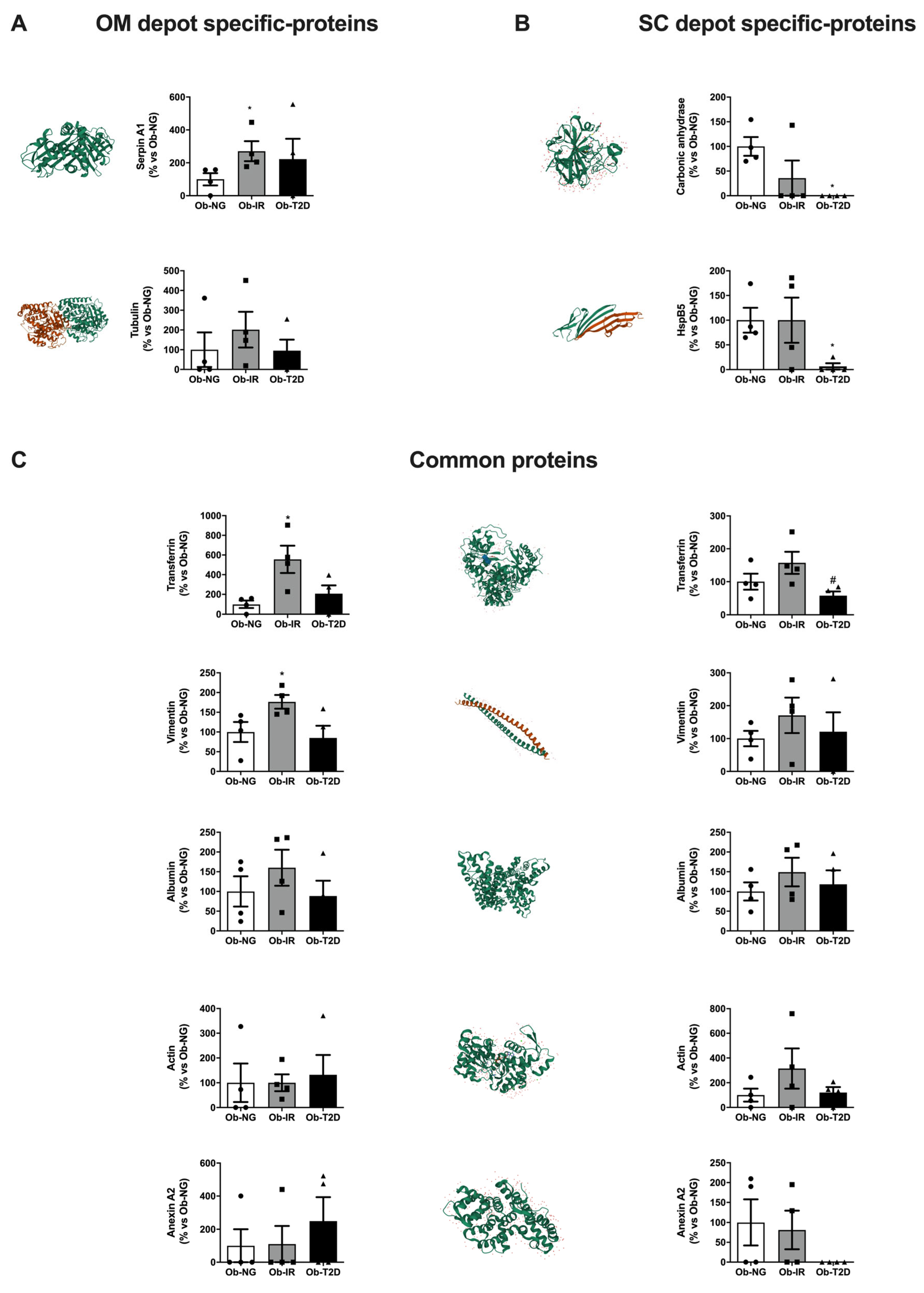
3.3. Adipose Tissue Carbonylome in Obesity-Associated Insulin Resistance
3.4. The Effect of Hyperglycaemia/Hyperinsulinemia on Adipose Tissue Carbonylome
3.5. Impact of Iron Homeostasis on Adipose Tissue in Obesity-Associated Insulin Resistance
3.6. Effect of Transferrin Protein Carbonylation on Molecular Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curtis, J.M.; Hahn, W.S.; Burril, J.S.; Bernlohr, D.A. Protein Carbonylation and Metabolic Control Systems. Trends Endocrinol. Metab. 2012, 23, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2020, 10, 1638. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.; Davies, M.J.; Grune, T. Determination of protein carbonyls in plasma, cell extracts, tissue homogenates, isolated proteins: Focus on sample preparation and derivatization conditions. Redox Biol. 2015, 5, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogowska-Wrzesinska, A.; Wojdyla, K.; Nedić, O.; Baron, C.P.; Griffiths, H.R. Analysis of protein carbonylation—Pitfalls and promise in commonly used methods. Free Radic. Res. 2014, 48, 1145–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tola, A.J.; Jaballi, A.; Missihoun, T.D. Protein carbonylation: Emerging roles in plant redox biology and future prospects. Plants 2021, 10, 1451. [Google Scholar] [CrossRef] [PubMed]
- Hauck, A.K.; Huang, Y.; Hertzel, A.V.; Bernlohr, D.A. Adipose oxidative stress and protein carbonylation. J. Biol. Chem. 2019, 294, 1083–1088. [Google Scholar] [CrossRef] [Green Version]
- Jaganjac, M.; Almuraikhy, S.; Al-Khelaifi, F.; Al-Jaber, M.; Bashah, M.; Mazloum, N.A.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Kafienah, W.; et al. Combined metformin and insulin treatment reverses metabolically impaired omental adipogenesis and accumulation of 4-hydroxynonenal in obese diabetic patients. Redox Biol. 2017, 12, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Sabbagh, F.; Muhamad, I.I.; Niazmand, R.; Dikshit, P.K.; Kim, B.S. Recent progress in polymeric non-invasive insulin delivery. Int. J. Biol. Macromol. 2022, 203, 222–243. [Google Scholar] [CrossRef] [PubMed]
- Grimsrud, P.A.; Picklo, M.J.; Griffin, T.J.; Bernlohr, D.A. Carbonylation of adipose proteins in obesity and insulin resistance: Identification of adipocyte fatty acid-binding protein as a cellular target of 4-hydroxynonenal. Mol. Cell. Proteomics. 2007, 6, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Frohnert, B.I.; Bernlohr, D.A. Protein carbonylation, mitochondrial dysfunction, and insulin resistance. Adv. Nutr. 2013, 4, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Ruskovska, T.; Bernlohr, D.A. Numerous questions remain, including how gradients of ROS inside the cell impact signaling cascades and gene regulation. J. Proteom. 2013, 23, 1–7. [Google Scholar] [CrossRef]
- Boden, G.; Homko, C.; Barrero, C.A.; Stein, T.P.; Chen, X.; Cheung, P.; Fecchio, C.; Koller, S.; Merali, S. Excessive caloric intake acutely causes oxidative stress, GLUT4 carbonylation, and insulin resistance in healthy men. Sci. Transl. Med. 2015, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Ruiz, A.; Guzmán-Ruiz, R.; Moreno, N.R.; García-Rios, A.; Delgado-Casado, N.; Membrives, A.; Túnez, I.; El Bekay, R.; Fernández-Real, J.M.; Tovar, S.; et al. Proteasome Dysfunction Associated to Oxidative Stress and Proteotoxicity in Adipocytes Compromises Insulin Sensitivity in Human Obesity. Antioxid. Redox Signal. 2015, 23, 597–612. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.J.; Gauthier, M.-S.; Hess, D.T.; Apovian, C.M.; Cacicedo, J.M.; Gokce, N.; Farb, M.; Valentine, R.J.; Ruderman, N.B. Insulin sensitive and resistant obesity in humans: AMPK activity, oxidative stress, and depot-specific changes in gene expression in adipose tissue. J. Lipid Res. 2012, 53, 792–801. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Ruiz, R.; Tercero-Alcázar, C.; Rabanal-Ruiz, Y.; Díaz-Ruiz, A.; El Bekay, R.; Rangel-Zuñiga, O.A.; Navarro-Ruiz, M.C.; Molero, L.; Membrives, A.; Ruiz-Rabelo, J.F.; et al. Adipose tissue depot-specific intracellular and extracellular cues contributing to insulin resistance in obese individuals. FASEB J. 2020, 34, 7520–7539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente-Cantarero, F.; Gil-Campos, M.; Benitez-Sillero, J.; Muñoz-Villanueva, M.; Túnez, I.; Pérez-Navero, J. Prepubertal children with suitable fitness and physical activity present reduced risk of oxidative stress. Free Radic. Biol. Med. 2012, 53, 415–420. [Google Scholar] [CrossRef]
- Care, D.; Suppl, S.S. Classification and diagnosis of diabetes: Standards of medical care in diabetes-2021. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef]
- Peinado, J.R.; Quiros, P.M.; Pulido, M.R.; Mariño, G.; Chantar, M.L.M.; Vázquez-Martínez, R.; Freije, J.M.; López-Otín, C.; Malagón, M.M. Proteomic profiling of adipose tissue from Zmpste24-/- mice, a model of lipodystrophy and premature aging, reveals major changes in mitochondrial function and vimentin processing. Mol. Cell. Proteom. 2011, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Ruiz, M.D.C.; López-Alcalá, J.; Díaz-Ruiz, A.; Del Moral, S.D.; Tercero-Alcázar, C.; Nieto-Calonge, A.; López-Miranda, J.; Tinahones, F.J.; Malagón, M.M.; Guzmán-Ruiz, R. Understanding the adipose tissue acetylome in obesity and insulin resistance. Transl. Res. 2022, 246, 15–32. [Google Scholar] [CrossRef]
- Sánchez-Ceinos, J.; Rangel-Zuñiga, O.A.; Clemente-Postigo, M.; Podadera-Herreros, A.; Camargo, A.; Alcalá-Diaz, J.F.; Guzmán-Ruiz, R.; López-Miranda, J.; Malagón, M.M. miR-223-3p as a potential biomarker and player for adipose tissue dysfunction preceding type 2 diabetes onset. Mol. Ther. Nucleic Acids 2021, 23, 1035–1052. [Google Scholar] [CrossRef] [PubMed]
- Almabouada, F.; Diaz-Ruiz, A.; Rabanal-Ruiz, Y.; Peinado, J.R.; Vazquez-Martinez, R.; Malagon, M.M. Adiponectin receptors form homomers and heteromers exhibiting distinct ligand binding and intracellular signaling properties. J. Biol. Chem. 2013, 288, 3112–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, W.; Xu, Z.; Liu, Y.; Chang, H.; Yu, P.; Zhang, X.; Yang, Y.; Liu, K.; Chang, Y. Iron regulatory protein 2 deficiency may correlate with insulin resistance. Biochem. Biophys. Res. Commun. 2019, 510, 191–197. [Google Scholar] [CrossRef]
- Fernández-Real, J.M.; Blasco, G.; Puig, J.; Moreno, M.; Xifra, G.; Sánchez-Gonzalez, J.; Alustiza, J.M.; Pedraza, S.; Ricart, W.; Moreno-Navarrete, J.M. Adipose tissue R2∗ signal is increased in subjects with obesity: A preliminary MRI study. Obesity 2016, 24, 352–358. [Google Scholar] [CrossRef] [Green Version]
- McClain, D.; Sharma, N.K.; Jain, S.; Harrison, A.; Salaye, L.N.; E Comeau, M.; Langefeld, C.D.; Lorenzo, F.R.; Das, S.K. Adipose tissue transferrin and insulin resistance. J. Clin. Endocrinol. Metab. 2018, 103, 4197–4208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guglielmi, V.; D’Adamo, M.; Bellia, A.; Ciotto, R.T.; Federici, M.; Lauro, D.; Sbraccia, P. Iron status in obesity: An independent association with metabolic parameters and effect of weight loss. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 541–547. [Google Scholar] [CrossRef]
- Fernández-Real, J.M.; Manco, M. Effects of iron overload on chronic metabolic diseases. Lancet Diabetes Endocrinol. 2014, 2, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, A.; Korac, A.; Srdic-Galic, B.; Buzadzic, B.; Otasevic, V.; Stancic, A.; Vucetic, M.; Markelic, M.; Velickovic, K.; Golic, I.; et al. Differences in the redox status of human visceral and subcutaneous adipose tissues—Relationships to obesity and metabolic risk. Metabolism 2014, 63, 661–671. [Google Scholar] [CrossRef]
- Ando, Y.; Kuroda, A.; Kusama, K.; Matsutani, T.; Matsuda, A.; Tamura, K. Impact of serine protease inhibitor alpha1-antitrypsin on expression of endoplasmic reticulum stress-induced proinflammatory factors in adipocytes. Biochem. Biophys. Rep. 2021, 26, 100967. [Google Scholar] [CrossRef]
- Shen, W.J.; Patel, S.; Eriksson, J.E.; Kraemer, F.B. Vimentin is a functional partner of hormone sensitive lipase and facilitates lipolysis. J. Proteome Res. 2010, 9, 1786–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, Y.; Hosaka, T.; Iwata, T.; Le, C.T.; Jambaldorj, B.; Teshigawara, K.; Harada, N.; Sakaue, H.; Sakai, T.; Yoshimoto, K.; et al. Vimentin binds IRAP and is involved in GLUT4 vesicle trafficking. Biochem. Biophys. Res. Commun. 2011, 405, 96–101. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, I.; Cho, W.; Oh, G.T.; Park, Y.M. Vimentin deficiency prevents high-fat diet-induced obesity and insulin resistance in mice. Diabetes Metab. J. 2021, 45, 97–108. [Google Scholar] [CrossRef]
- Molina-Ayala, M.A.; Rodríguez-Amador, V.; Suárez-Sánchez, R.; León-Solís, L.; Gómez-Zamudio, J.; Mendoza-Zubieta, V.; Cruz, M.; Suárez-Sánchez, F. Expression of obesity- and type-2 diabetes-associated genes in omental adipose tissue of individuals with obesity. Gene 2022, 815. [Google Scholar] [CrossRef]
- Lampson, M.A.; Schmoranzer, J.; Zeigerer, A.; Simon, S.M.; McGraw, T.E. Insulin-regulated release from the endosomal recycling compartment is regulated by budding of specialized vesicles. Mol. Biol. Cell. 2001, 12, 3489–3501. [Google Scholar] [CrossRef] [Green Version]
- Holman, G.D.; Sandoval, I.V. Moving the insulin-regulated glucose transporter GLUT4 into and out of storage. Trends Cell. Biol. 2001, 11, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Ramm, G.; Slot, J.W.; James, D.E.; Stoorvogel, W. Insulin recruits GLUT4 from specialized VAMP2-carrying vesicles as well as from the dynamic endosomal/trans-golgi network in rat adipocytes. Mol. Biol. Cell. 2000, 11, 4079–4091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin Lawrence, C.; Ray, S.; Babyonyshev, M.; Galluser, R.; Borhani, D.W.; SCH. Borhani and SCH. Lawrence Tfr Structure.Pdf. Available online: https://www.jstor.org/stable/2899387 (accessed on 1 June 2022).
- Cheng, Y.; Zak, O.; Aisen, P.; Harrison, S.C.; Walz, T.; York, N. Structure of the Human Transferrin Receptor-Transferrin Complex Albert Einstein College of Medicine. Cell 2004, 116, 565–576. Available online: http://www.ncbi.nlm.nih.gov/pubmed/14980223 (accessed on 1 June 2022). [CrossRef] [PubMed] [Green Version]
- Chen, J.; Sawyer, N.; Regan, L. Protein-protein interactions: General trends in the relationship between binding affinity and interfacial buried surface area. Protein Sci. 2013, 22, 510–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Zhou, Q.; He, J.; Jiang, Z.; Peng, C.; Tong, R.; Shi, J. Recent advances in the development of protein–protein interactions modulators: Mechanisms and clinical trials. Signal Transduct. Target. Ther. 2020, 5. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ob-NG (n = 20) | Ob-IR (n = 37) | Ob-T2D (n = 16) | ANOVA | |
|---|---|---|---|---|
| Sex (men/women) | 8/12 | 11/26 | 8/8 | |
| Age (years) | 34.50 ± 2.26 | 42.14 ± 1.68 a | 44.69 ± 2.02 aa | <0.01 |
| Lipid-lowering therapy (n, %) | 2, 10 | 7, 19 | 4, 25 | |
| Antidiabetic therapy (n, %) | 2, 10 | 2, 5.4 | 8, 50 | |
| Antihypertensive therapy (n, %) | 3, 15 | 18, 48.6 | 7, 43.8 | |
| Weight (kg) | 143.90 ± 6.5 | 148.60 ± 6.14 | 156.4 ± 6.26 | ns |
| BMI (kg/m2) | 52.38 ± 2.37 | 53.25 ± 1.80 | 54.53 ± 1.49 | ns |
| Waist circumference (cm) | 136.6 ± 4.06 | 145.3 ± 3.85 | 149.0 ± 3.53 | ns |
| Glucose (mg/dL) | 90.95 ± 2.00 | 107.9 ± 1.66 aaa | 171.4 ± 9.92 aaa,bbb | <0.0001 |
| HbAc1 (mmol/mol) | 5.64 ± 0.12 | 6.24 ± 0.22 aa | 9.17 ± 0.5 aaa,bbb | <0.0001 |
| HOMA-IR | 3.31 ± 0.37 | 5.49 ± 0.44 aa | 8.44 ± 1.42 aaa | <0.001 |
| Insulin (mU/L) | 14.72 ± 1.5 | 21.06 ± 1.73 | 19.88 ± 2.56 | ns |
| Triglycerides (mg/dL) | 121.1 ± 12.42 | 117 ± 7.91 | 195 ± 30.71 aa,bb | <0.001 |
| Cholesterol (mg/dL) | 182 ± 10.68 | 201.1 ± 5.96 | 195.8 ± 9.5 | ns |
| HDL (mg/dL) | 40.47 ± 3.05 | 41.86 ± 1.5 | 35.5 ± 1.66 | ns |
| LDL (mg/dL) | 121.2 ± 8.47 | 136 ± 4.9 | 118.6 ± 7.99 | ns |
| Hemoglobin (g/dL) | 13.74 ± 0.38 | 13.31 ± 0.28 | 14.08 ± 0.47 | ns |
| Hematocrit (%) | 41.52 ± 0.94 | 39.47 ± 1.25 | 42.61 ± 1.11 | ns |
| Iron (ug/dL) | 82.42 ± 6.28 | 68.35 ± 5.33 | 69.13 ± 7.74 | ns |
| Ferritin (ng/mL) | 83.59 ± 18.04 | 98.54 ± 15.09 | 149.9 ± 27.56 | ns |
| Transferrin (mg/dL) | 254 ± 12.39 | 279.6 ± 8.04 | 276.1 ± 10.07 | ns |
| PC (pM/L) | 8.22 ± 0.88 | 8.54 ± 1.01 | 8.49 ± 1.01 | ns |
| LPO (μM/L) | 5.75 ± 0.76 | 5.74 ± 1.00 | 5.47 ± 0.64 | ns |
| NOx (μM/L) | 29.5 ± 1.18 | 28.8 ± 1.67 | 42.8 ± 2.84 | ns |
| GSH (mmol/mg Hb) | 1.5 × 10−5 ± 1.9 × 10−6 | 1.6 × 10−5 ± 5.9 × 10−6 | 1.2E-5 ± 1.0 × 10−6 | ns |
| GT (mmol/mg Hb) | 1.8 × 10−5 ± 2.2 × 10−6 | 2.1 × 10−5 ± 9.1 × 10−6 | 1.4E-5 ± 1.1 × 10−6 | ns |
| GPx (UI/mg prot) | 2.44 ± 0.06 | 2.33 ± 0.03 | 2.28 ± 0.02 | ns |
| Spot Num. a | Protein Name | Symbol | Accession Number b | MW (KDa)/pI | %Cover. c | Pep. d | Score |
|---|---|---|---|---|---|---|---|
| Omental adipose tissue (OM depot) | |||||||
| 1 | Transferrin | TRFE_HUMAN | P02787 | 79.3/6.81 | 24 | 19 | 222 |
| 2 | Transferrin | TRFE_HUMAN | P02787 | 79.3/6.81 | 35 | 32 | 613 |
| 3 | Transferrin | TRFE_HUMAN | P02787 | 79.3/6.81 | 39 | 35 | 695 |
| 4 | Serum albumin | ALBU_HUMAN | P02768 | 71.3/5.92 | 54 | 38 | 760 |
| 5 | Alpha-1-antitrypsin | A1AT_HUMAN | P01009 | 46.9/5.37 | 43 | 19 | 297 |
| 6 | Alpha-1-antitrypsin | A1AT_HUMAN | P01009 | 46.9/5.37 | 25 | 14 | 106 |
| 7 | Alpha-1-antitrypsin | A1AT_HUMAN | P01009 | 46.9/5.37 | 43 | 19 | 252 |
| 8 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 61 | 32 | 512 |
| 9 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 54 | 30 | 548 |
| 10 | Tubulin beta-4B chain | TBB4B_HUMAN | P68371 | 50.3/4.79 | 27 | 14 | 157 |
| 11 | Alpha-1-antitrypsin | A1AT_HUMAN | P01009 | 46.9/5.37 | 40 | 18 | 368 |
| 12 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 51 | 25 | 454 |
| 13 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 59 | 34 | 548 |
| 14 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 31 | 23 | 378 |
| 15 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 42 | 26 | 403 |
| 16 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 31 | 19 | 241 |
| 17 | Not detected | ||||||
| 18 | Not detected | ||||||
| 19 | Actin, cytoplasmic 1 | ACTB_HUMAN | P60709 | 42.1/5.29 | 27 | 18 | 352 |
| 20 | Actin, cytoplasmic 1 | ACTB_HUMAN | P60709 | 42.1/5.29 | 57 | 21 | 488 |
| 21 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 47 | 30 | 452 |
| 22 | Actin, cytoplasmic 1 | ACTB_HUMAN | P60709 | 42.1/5.29 | 53 | 26 | 487 |
| 23 | Annexin A2 | ANXA2_HUMAN | P07355 | 38.8/7.57 | 52 | 22 | 430 |
| Subcutaneous adipose tissue (SC depot) | |||||||
| 1 | Transferrin | TRFE_HUMAN | P02787 | 79.3/6.81 | 21 | 15 | 198 |
| 2 | Transferrin | TRFE_HUMAN | P02787 | 79.3/6.81 | 27 | 21 | 259 |
| 3 | Not detected | ||||||
| 4 | Serum albumin | ALBU_HUMAN | P02768 | 71.3/5.92 | 58 | 40 | 686 |
| 5 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 56 | 30 | 510 |
| 6 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 28 | 14 | 147 |
| 7 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 17 | 9 | 43 |
| 8 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 42 | 22 | 432 |
| 9 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 23 | 11 | 123 |
| 10 | Not detected | ||||||
| 11 | Not detected | ||||||
| 12 | Not detected | ||||||
| 13 | Vimentin | VIME_HUMAN | P08670 | 53.7/5.05 | 29 | 15 | 188 |
| 14 | Actin, cytoplasmic 2 | ACTG_HUMAN | P60709 | 42.1/5.29 | 46 | 15 | 379 |
| 15 | Not detected | ||||||
| 16 | Not detected | ||||||
| 17 | Annexin A2 | ANXA2_HUMAN | P07355 | 38.8/7.57 | 50 | 24 | 553 |
| 18 | Actin, cytoplasmic 2 | ACTG_HUMAN | P60709 | 42.1/5.29 | 30 | 9 | 159 |
| 19 | Carbonic anhydrase 1 | CAH1_HUMAN | P00915 | 28.9/6.59 | 57 | 15 | 401 |
| 20 | Not detected | ||||||
| 21 | Alpha-crystallin B chain | CRYAB_HUMAN | P02511 | 20.1/6.76 | 48 | 8 | 59 |
| 22 | Alpha-crystallin B chain | CRYAB_HUMAN | P02511 | 20.1/6.76 | 57 | 14 | 237 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Ruiz, M.C.; Soler-Vázquez, M.C.; Díaz-Ruiz, A.; Peinado, J.R.; Nieto Calonge, A.; Sánchez-Ceinos, J.; Tercero-Alcázar, C.; López-Alcalá, J.; Rangel-Zuñiga, O.A.; Membrives, A.; et al. Influence of Protein Carbonylation on Human Adipose Tissue Dysfunction in Obesity and Insulin Resistance. Biomedicines 2022, 10, 3032. https://doi.org/10.3390/biomedicines10123032
Navarro-Ruiz MC, Soler-Vázquez MC, Díaz-Ruiz A, Peinado JR, Nieto Calonge A, Sánchez-Ceinos J, Tercero-Alcázar C, López-Alcalá J, Rangel-Zuñiga OA, Membrives A, et al. Influence of Protein Carbonylation on Human Adipose Tissue Dysfunction in Obesity and Insulin Resistance. Biomedicines. 2022; 10(12):3032. https://doi.org/10.3390/biomedicines10123032
Chicago/Turabian StyleNavarro-Ruiz, M. Carmen, M. Carmen Soler-Vázquez, Alberto Díaz-Ruiz, Juan R. Peinado, Andrea Nieto Calonge, Julia Sánchez-Ceinos, Carmen Tercero-Alcázar, Jaime López-Alcalá, Oriol A. Rangel-Zuñiga, Antonio Membrives, and et al. 2022. "Influence of Protein Carbonylation on Human Adipose Tissue Dysfunction in Obesity and Insulin Resistance" Biomedicines 10, no. 12: 3032. https://doi.org/10.3390/biomedicines10123032