
How the Tumor Micromilieu Modulates the Recruitment and Activation of Colorectal Cancer-Infiltrating Lymphocytes
{kind=link}
{kind=link}
Abstract
:1. Introduction
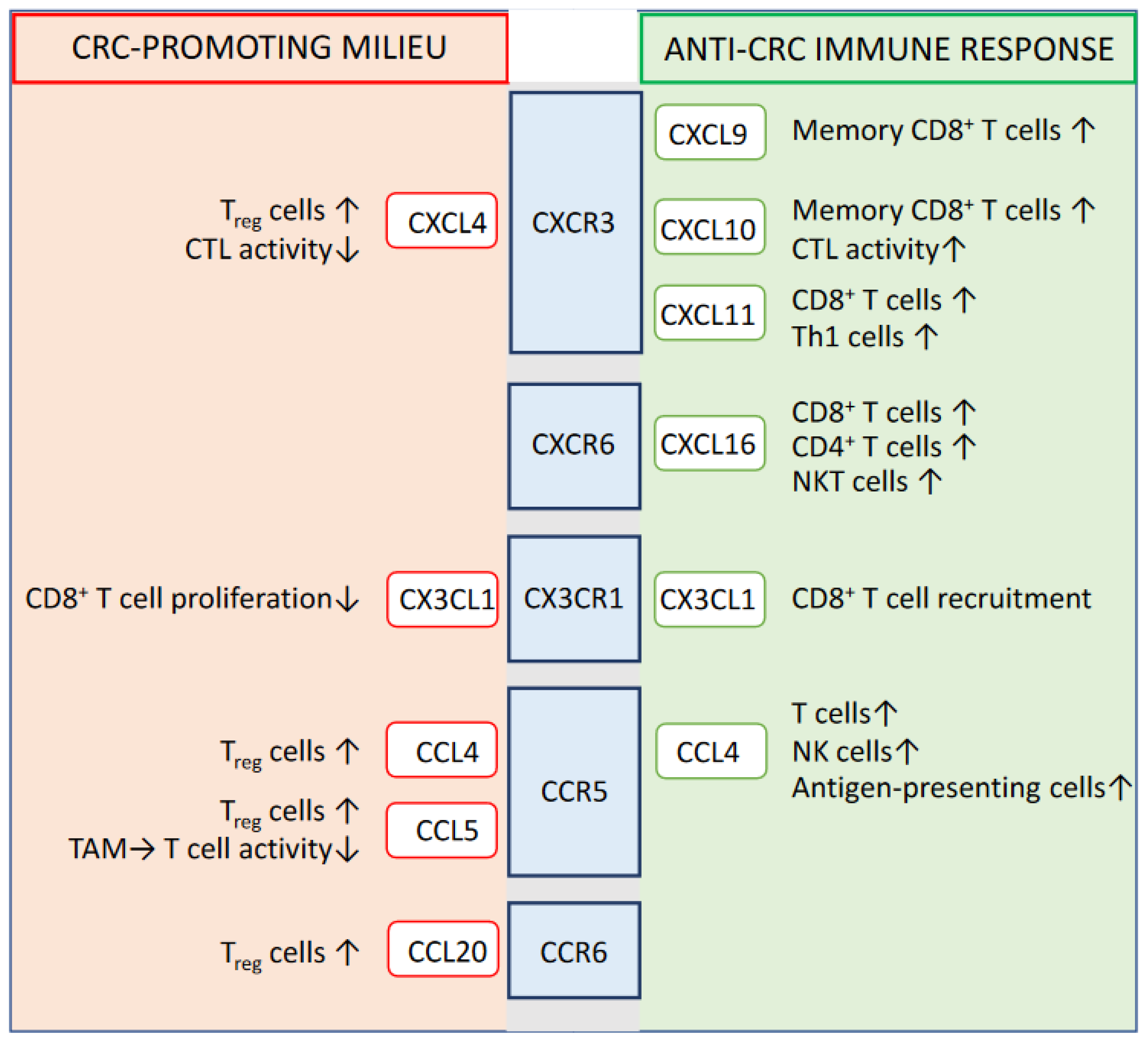
2. Prognostic-Relevant Chemokine Signaling Impacting the Immune System-Mediated Control of CRC
2.1. CXCR3
2.2. CXCR6
2.3. CX3CR1
2.4. CCR5
2.5. CCR6
3. Cytokines Influencing the Efficacy of the Antitumor Immune Response
3.1. IFNγ
3.2. IL-17A and IL-17F
3.3. IL-10
3.4. IL-6
3.5. IL-12 Family of Cytokines
3.6. TGFβ
4. Conclusions and Targets beyond Chemokines and Cytokines
Author Contributions
Funding
Conflicts of Interest
References
- Hossain, S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, D.J.; John, A.; Lim, Y.C.; Kibria, K.M.K.; Mohiuddin, A.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, M.; Greten, F.R. The inflammatory pathogenesis of colorectal cancer. Nat. Rev. Immunol. 2021, 21, 653–667. [Google Scholar] [CrossRef]
- Yue, Y.; Zhang, Q.; Sun, Z. CX3CR1 Acts as a Protective Biomarker in the Tumor Microenvironment of Colorectal Cancer. Front. Immunol. 2022, 12, 758040. [Google Scholar] [CrossRef]
- Pagès, F.; Berger, A.; Camus, M.; Sanchez-Cabo, F.; Costes, A.; Molidor, R.; Mlecnik, B.; Kirilovsky, A.; Nilsson, M.; Damotte, D.; et al. Effector memory T cells, early metastasis, and survival in colorectal cancer. N. Engl. J. Med. 2005, 353, 2654–2666. [Google Scholar] [CrossRef]
- Boquet, I.; Kassambara, A.; Lui, A.; Tanner, A.; Latil, M.; Lovera, Y.; Arnoux, F.; Hermitte, F.; Galon, J.; Catteau, A. Comparison of Immune Response Assessment in Colon Cancer by Immunoscore (Automated Digital Pathology) and Pathologist Visual Scoring. Cancers 2022, 14, 1170. [Google Scholar] [CrossRef]
- Argilés, G.; Tabernero, J.; Labianca, R.; Hochhauser, D.; Salazar, R.; Iveson, T.; Laurent-Puig, P.; Quirke, P.; Yoshino, T.; Taieb, J.; et al. Localised colon cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2020, 31, 1291–1305. [Google Scholar] [CrossRef]
- Liu, F.; Dong, X.; Pang, X.; Xing, Q.; Wang, H.; Zhang, H.; Li, Y.; Yin, Y.; Fant, M.; Ye, Y.; et al. The Specific Immune Response to Tumor Antigen CP1 and Its Correlation With Improved Survival in Colon Cancer Patients. Gastroenterology 2008, 134, 998–1006. [Google Scholar] [CrossRef]
- Pancione, M.; Giordano, G.; Remo, A.; Febbraro, A.; Sabatino, L.; Manfrin, E.; Ceccarelli, M.; Colantuoni, V. Immune Escape Mechanisms in Colorectal Cancer Pathogenesis and Liver Metastasis. J. Immunol. Res. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Fidelle, M.; Yonekura, S.; Picard, M.; Cogdill, A.; Hollebecque, A.; Roberti, M.P.; Zitvogel, L. Resolving the Paradox of Colon Cancer Through the Integration of Genetics, Immunology, and the Microbiota. Front. Immunol. 2020, 11, 600886. [Google Scholar] [CrossRef]
- Breakstone, R. Colon cancer and immunotherapy-can we go beyond microsatellite instability? Transl. Gastroenterol Hepatol. 2021, 6, 12. [Google Scholar] [CrossRef]
- Michael-Robinson, J.M.; Biemer-Hüttmann, A.-E.; Purdie, D.M.; Walsh, M.D.; Simms, L.A.; Biden, K.G.; Young, J.P.; Leggett, B.A.; Jass, J.R.; Radford-Smith, G.L. Tumour infiltrating lymphocytes and apoptosis are independent features in colorectal cancer stratified according to microsatellite instability status. Gut 2001, 48, 360–366. [Google Scholar] [CrossRef] [Green Version]
- DiToro, D.; Basu, R. Emerging Complexity in CD4+T Lineage Programming and Its Implications in Colorectal Cancer. Front. Immunol. 2021, 12, 3362. [Google Scholar] [CrossRef]
- Wu, P.; Wu, D.; Ni, C.; Ye, J.; Chen, W.; Hu, G.; Wang, Z.; Wang, C.; Zhang, Z.; Xia, W.; et al. γδT17 cells promote the accumulation and expansion of myeloid-derived suppressor cells in human colorectal cancer. Immunity. 2014, 40, 785–800. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Nishikawa, H.; Wada, H.; Nagano, Y.; Sugiyama, D.; Atarashi, K.; Maeda, Y.; Hamaguchi, M.; Ohkura, N.; Sato, E.; et al. Two FOXP3+CD4+ T cell subpopulations distinctly control the prognosis of colorectal cancers. Nat. Med. 2016, 22, 679–684. [Google Scholar] [CrossRef]
- Hanke, T.; Melling, N.; Simon, R.; Sauter, G.; Bokemeyer, C.; Lebok, P.; Terracciano, L.M.; Izbicki, J.R.; Marx, A.H. High intratumoral FOXP3⁺ T regulatory cell (Tregs) density is an independent good prognosticator in nodal negative colorectal cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 8227–8235. [Google Scholar]
- Liu, Z.; Huang, Q.; Liu, G.; Dang, L.; Chu, D.; Tao, K.; Wang, W. Presence of FOXP3(+)Treg cells is correlated with colorectal cancer progression. Int. J. Clin. Exp. Med. 2014, 7, 1781–1785. [Google Scholar]
- Carluccio, S.; Delbue, S.; Signorini, L.; Setola, E.; Bagliani, A.; Della Valle, A.; Galli, A.; Ferrante, P.; Bregni, M. Generation of tumor-specific cytotoxic T-lymphocytes from the peripheral blood of colorectal cancer patients for adoptive T-cell transfer. J. Cell. Physiol. 2015, 230, 1457–1465. [Google Scholar] [CrossRef]
- Aparicio, C.; Belver, M.; Enríquez, L.; Espeso, F.; Núñez, L.; Sánchez, A.; de la Fuente, M.; González-Vallinas, M. Cell Therapy for Colorectal Cancer: The Promise of Chimeric Antigen Receptor (CAR)-T Cells. Int. J. Mol. Sci. 2021, 22, 11781. [Google Scholar] [CrossRef]
- Erreni, M.; Bianchi, P.; Laghi, L.; Mirolo, M.; Fabbri, M.; Locati, M.; Mantovani, A.; Allavena, P. Chapter 5 Expression of Chemokines and Chemokine Receptors in Human Colon Cancer. Methods Enzymol. 2009, 460, 105–121. [Google Scholar] [CrossRef]
- Shamoun, L.; Landerholm, K.; Ramilo, A.B.; E Andersson, R.; Dimberg, J.; Wågsäter, D. Association of gene and protein expression and genetic polymorphism of CC chemokine ligand 4 in colorectal cancer. World J. Gastroenterol. 2021, 27, 5076–5087. [Google Scholar] [CrossRef]
- Siddiqui, I.; Erreni, M.; van Brakel, M.; Debets, R.; Allavena, P. Enhanced recruitment of genetically modified CX3CR1-positive human T cells into Fractalkine/CX3CL1 expressing tumors: Importance of the chemokine gradient. J. Immunother. Cancer 2016, 4, 21. [Google Scholar] [CrossRef]
- Erreni, M.; Siddiqui, I.; Marelli, G.; Grizzi, F.; Bianchi, P.; Morone, D.; Marchesi, F.; Celesti, G.; Pesce, S.; Doni, A.; et al. The Fractalkine-Receptor Axis Improves Human Colorectal Cancer Prognosis by Limiting Tumor Metastatic Dissemination. J. Immunol. 2015, 196, 902–914. [Google Scholar] [CrossRef] [Green Version]
- Hojo, S.; Koizumi, K.; Tsuneyama, K.; Arita, Y.; Cui, Z.; Shinohara, K.; Minami, T.; Hashimoto, I.; Nakayama, T.; Sakurai, H.; et al. High-Level Expression of Chemokine CXCL16 by Tumor Cells Correlates with a Good Prognosis and Increased Tumor-Infiltrating Lymphocytes in Colorectal Cancer. Cancer Res. 2007, 67, 4725–4731. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, Y.; Sun, X.; Deng, G.; Huang, W.; Wu, X.; Gu, Y.; Tian, Z.; Fan, Z.; Xu, Q.; et al. CXCR6 is required for antitumor efficacy of intratumoral CD8+ T cell. J. Immunother. Cancer 2021, 9, e003100. [Google Scholar] [CrossRef]
- Nagasawa, S.; Tsuchida, K.; Shiozawa, M.; Hiroshima, Y.; Kimura, Y.; Hashimoto, I.; Watanabe, H.; Kano, K.; Numata, M.; Aoyama, T.; et al. Clinical Significance of Chemokine Receptor CXCR4 and CCR7 mRNA Expression in Patients With Colorectal Cancer. Anticancer Res. 2021, 41, 4489–4495. [Google Scholar] [CrossRef]
- Wang, D.; Yang, L.; Yu, W.; Wu, Q.; Lian, J.; Li, F.; Liu, S.; Li, A.; He, Z.; Liu, J.; et al. Colorectal cancer cell-derived CCL20 recruits regulatory T cells to promote chemoresistance via FOXO1/CEBPB/NF- κB signaling. J. Immunother. Cancer. 2019, 7, 215. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Han, X.; Yan, J.; Pan, Y.; Gong, J.; Di, J.; Cheng, Z.; Jin, Z.; Wang, Z.; Zheng, Q.; et al. The prognostic significance of chemokine receptor CXCR3 expression in colorectal carcinoma. Biomed. Pharmacother. 2012, 66, 373–377. [Google Scholar] [CrossRef]
- Kim, J.; Takeuchi, H.; Lam, S.T.; Turner, R.R.; Wang, H.-J.; Kuo, C.; Foshag, L.; Bilchik, A.J.; Hoon, D.S. Chemokine Receptor CXCR4 Expression in Colorectal Cancer Patients Increases the Risk for Recurrence and for Poor Survival. J. Clin. Oncol. 2005, 23, 2744–2753. [Google Scholar] [CrossRef]
- Mlecnik, B.; Tosolini, M.; Charoentong, P.; Kirilovsky, A.; Bindea, G.; Berger, A.; Camus, M.; Gillard, M.; Bruneval, P.; Fridman, W.H.; et al. Biomolecular Network Reconstruction Identifies T-Cell Homing Factors Associated With Survival in Colorectal Cancer. Gastroenterology 2010, 138, 1429–1440. [Google Scholar] [CrossRef]
- Kistner, L.; Doll, D.; Holtorf, A.; Nitsche, U.; Janssen, K.-P. Interferon-inducible CXC-chemokines are crucial immune modulators and survival predictors in colorectal cancer. Oncotarget 2017, 8, 89998–90012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.-N.; Li, M.-Q.; Deng, S.-H.; Chen, C.; Ni, Y.; Cui, B.-B.; Liu, Y.-L. Prognostic Immune-Related Analysis Based on Differentially Expressed Genes in Left- and Right-Sided Colon Adenocarcinoma. Front. Oncol. 2021, 11, 640196. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, T.; Moehler, M.; Gockel, I.; Sgourakis, G.G.; Biesterfeld, S.; Müller, M.; Berger, M.R.; Lang, H.; Galle, P.R.; Schimanski, C.C. Low expression of chemokine receptor CCR5 in human colorectal cancer correlates with lymphatic dissemination and reduced CD8+ T-cell infiltration. Int. J. Color. Dis. 2010, 25, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Karin, N. CXCR3 Ligands in Cancer and Autoimmunity, Chemoattraction of Effector T Cells, and Beyond. Front. Immunol. 2020, 11, 976. [Google Scholar] [CrossRef]
- Zumwalt, T.J.; Arnold, M.; Goel, A.; Boland, C.R. Active secretion of CXCL10 and CCL5 from colorectal cancer microenvironments associates with GranzymeB+ CD8+ T-cell infiltration. Oncotarget 2014, 6, 2981–2991. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.; Santoni, A.; Bernardini, G. Tumor inhibition or tumor promotion? The duplicity of CXCR3 in cancer. J. Leukoc. Biol. 2020, 108, 673–685. [Google Scholar] [CrossRef]
- Kawada, K.; Hosogi, H.; Sonoshita, M.; Sakashita, H.; Manabe, T.; Shimahara, Y.; Sakai, Y.; Takabayashi, A.; Oshima, M.; Taketo, M.M. Chemokine receptor CXCR3 promotes colon cancer metastasis to lymph nodes. Oncogene 2007, 26, 4679–4688. [Google Scholar] [CrossRef]
- Reynders, N.; Abboud, D.; Baragli, A.; Noman, M.Z.; Rogister, B.; Niclou, S.P.; Heveker, N.; Janji, B.; Hanson, J.; Szpakowska, M.; et al. The Distinct Roles of CXCR3 Variants and Their Ligands in the Tumor Microenvironment. Cells 2019, 8, 613. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Deng, Q.; Zhang, Y.; Ye, H.; Yu, X.; Zhang, Y.; Han, G.Y.; Luo, P.; Wu, M.; Yu, Y.; et al. Non-platelet-derived CXCL4 differentially regulates cytotoxic and regulatory T cells through CXCR3 to suppress the immune response to colon cancer. Cancer Lett. 2018, 443, 1–12. [Google Scholar] [CrossRef]
- Abron, J.D.; Singh, N.P.; Murphy, A.E.; Mishra, M.K.; Price, R.L.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P. Differential role of CXCR3 in inflammation and colorectal cancer. Oncotarget 2018, 9, 17928–17936. [Google Scholar] [CrossRef] [Green Version]
- La Porta, C.A. CXCR6: The role of environment in tumor progression. Challenges for therapy. Stem Cell Rev. Rep. 2012, 8, 1282–1285. [Google Scholar] [CrossRef]
- Kee, J.-Y.; Ito, A.; Hojo, S.; Hashimoto, I.; Igarashi, Y.; Tsukada, K.; Irimura, T.; Shibahara, N.; Nakayama, T.; Yoshie, O.; et al. Chemokine CXCL16 suppresses liver metastasis of colorectal cancer via augmentation of tumor-infiltrating natural killer T cells in a murine model. Oncol. Rep. 2013, 29, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Hoki, T.; Oba, T.; Saito, H.; Attwood, K.; Sabel, M.S.; Chang, A.E.; Odunsi, K.; Ito, F. CX3CR1–CD8+ T cells are critical in antitumor efficacy but functionally suppressed in the tumor microenvironment. JCI Insight 2020, 5, e133920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, M.; Tanaka, F.; Yamaguchi, H.; Sadanaga, N.; Inoue, H.; Mori, M. The high expression of Fractalkine results in a better prognosis for colorectal cancer patients. Int. J. Oncol. 2005, 26, 41–47. [Google Scholar] [CrossRef]
- Menten, P.; Wuyts, A.; Van Damme, J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev. 2002, 13, 455–481. [Google Scholar] [CrossRef]
- Kulkarni, N.; Pathak, M.; Lal, G. Role of chemokine receptors and intestinal epithelial cells in the mucosal inflammation and tolerance. J. Leukoc. Biol. 2016, 101, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.T.; Li, K.K.; Hepburn, E.; Weston, C.J.; Curbishley, S.M.; Reynolds, G.M.; Hejmadi, R.K.; Bicknell, R.; Eksteen, B.; Ismail, T.; et al. The effects of CCR5 inhibition on regulatory T-cell recruitment to colorectal cancer. Br. J. Cancer 2014, 112, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Yu, X.; Xue, L.; Ge, X.; Zhao, W.; Peng, W. Intrinsic beta-catenin signaling suppresses CD8(+) T-cell infiltration in colorectal cancer. Biomed. Pharmacother. 2019, 115, 108921. [Google Scholar] [CrossRef]
- Spranger, S.; Bao, R.; Gajewski, T.F. Melanoma-intrinsic beta-catenin signalling prevents anti-tumour immunity. Nature 2015, 523, 231–235. [Google Scholar] [CrossRef]
- Lillard, J.W., Jr.; Singh, U.P.; Boyaka, P.N.; Singh, S.; Taub, D.D.; McGhee, J.R. MIP-1alpha and MIP-1beta differentially mediate mucosal and systemic adaptive immunity. Blood 2003, 101, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.Y.; Lin, Y.C.; Mahalingam, J.; Huang, C.T.; Chen, T.W.; Kang, C.W.; Peng, H.M.; Chu, Y.Y.; Chiang, J.M.; Dutta, A.; et al. Tumor-derived chemokine CCL5 enhances TGF-beta-mediated killing of CD8(+) T cells in colon cancer by T-regulatory cells. Cancer research. 2012, 72, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhong, M.; Wang, C.; Xu, Y.; Gao, W.-Q.; Zhang, Y. CCL5-deficiency enhances intratumoral infiltration of CD8+ T cells in colorectal cancer. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.H.; Lee, K.Y.; Chang, Y.L.; Chan, Y.F.; Kuo, L.W.; Lin, T.Y.; Chung, F.T.; Kuo, C.S.; Yu, C.T.; Lin, S.M.; et al. CD14(+)S100A9(+) monocytic myeloid-derived suppressor cells and their clinical relevance in non-small cell lung cancer. Am. J. Respir. Crit. Care Med. 2012, 186, 1025–1036. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; Baba, T.; Shinagawa, K.; Matsushima, K.; Mukaida, N. Crucial involvement of the CCL3-CCR5 axis-mediated fibroblast accumulation in colitis-associated carcinogenesis in mice. Int. J. Cancer 2014, 135, 1297–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Zhou, Y.; Li, C.; Rychahou, P.; Zhang, S.; Titlow, W.B.; Bauman, G.; Wu, Y.; Liu, J.; Wang, C.; et al. Ketogenesis Attenuates KLF5-Dependent Production of CXCL12 to Overcome the Immunosuppressive Tumor Microenvironment in Colorectal Cancer. Cancer Res. 2022, 82, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Banerjee, D.; Ben-Baruch, A. Chemokines at the crossroads of tumor-fibroblast interactions that promote malignancy. J. Leukoc. Biol. 2010, 89, 31–39. [Google Scholar] [CrossRef]
- Halama, N.; Zoernig, I.; Berthel, A.; Kahlert, C.; Klupp, F.; Suarez-Carmona, M.; Suetterlin, T.; Brand, K.; Krauss, J.; Lasitschka, F.; et al. Tumoral Immune Cell Exploitation in Colorectal Cancer Metastases Can Be Targeted Effectively by Anti-CCR5 Therapy in Cancer Patients. Cancer Cell 2016, 29, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Ghadjar, P.; Coupland, S.; Na, I.-K.; Noutsias, M.; Letsch, A.; Stroux, A.; Bauer, S.; Buhr, H.J.; Thiel, E.; Scheibenbogen, C.; et al. Chemokine Receptor CCR6 Expression Level and Liver Metastases in Colorectal Cancer. J. Clin. Oncol. 2006, 24, 1910–1916. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, N.; Li, Q.; Zhang, W.; Ke, F.; Leng, Q.; Wang, H.; Chen, J.; Wang, H. Tumor-Associated Macrophages Recruit CCR6+ Regulatory T Cells and Promote the Development of Colorectal Cancer via Enhancing CCL20 Production in Mice. PLoS ONE 2011, 6, e19495. [Google Scholar] [CrossRef]
- Sun, Z.; Du, C.; Xu, P.; Miao, C. Surgical trauma-induced CCL18 promotes recruitment of regulatory T cells and colon cancer progression. J. Cell. Physiol. 2019, 234, 4608–4616. [Google Scholar] [CrossRef] [Green Version]
- Märkl, F.; Huynh, D.; Endres, S.; Kobold, S. Utilizing chemokines in cancer immunotherapy. Trends Cancer 2022, 8, 670–682. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Song, Z.; Chu, J.; Qu, X. Deficiency of Interferon-Gamma or Its Receptor Promotes Colorectal Cancer Development. J. Interf. Cytokine Res. 2015, 35, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Schwartzkopff, J.; Pradera, F.; Kammertoens, T.; Seliger, B.; Pircher, H.; Blankenstein, T. A critical requirement of interferon gamma-mediated angiostasis for tumor rejection by CD8+ T cells. Cancer Res. 2003, 63, 4095–4100. [Google Scholar]
- Burke, J.D.; Young, H.A. IFN-γ: A cytokine at the right time, is in the right place. Semin. Immunol. 2019, 43, 101280. [Google Scholar] [CrossRef] [PubMed]
- Ling, A.; Lundberg, I.V.; Eklöf, V.; Wikberg, M.L.; Öberg, Å.; Edin, S.; Palmqvist, R. The infiltration, and prognostic importance, of Th1 lymphocytes vary in molecular subgroups of colorectal cancer. J. Pathol. Clin. Res. 2016, 2, 21–31. [Google Scholar] [CrossRef]
- Coppola, A.; Arriga, R.; Lauro, D.; Del Principe, M.I.; Buccisano, F.; Maurillo, L.; Palomba, P.; Venditti, A.; Sconocchia, G. NK Cell Inflammation in the Clinical Outcome of Colorectal Carcinoma. Front. Med. 2015, 2, 33. [Google Scholar] [CrossRef] [Green Version]
- Katlinski, K.V.; Gui, J.; Katlinskaya, Y.V.; Ortiz, A.; Chakraborty, R.; Bhattacharya, S.; Carbone, C.J.; Beiting, D.P.; Girondo, M.A.; Peck, A.R.; et al. Inactivation of Interferon Receptor Promotes the Establishment of Immune Privileged Tumor Microenvironment. Cancer Cell 2017, 31, 194–207. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Li, Y.; Zhang, J.; Zhang, B. PD-L1 expression increased by IFN-γ via JAK2-STAT1 signaling and predicts a poor survival in colorectal cancer. Oncol. Lett. 2020, 20, 1127–1134. [Google Scholar] [CrossRef]
- Ren, J.; Li, N.; Pei, S.; Lian, Y.; Li, L.; Peng, Y.; Liu, Q.; Guo, J.; Wang, X.; Han, Y.; et al. Histone methyltransferase WHSC1 loss dampens MHC-I antigen presentation pathway to impair IFN-γ-stimulated antitumor immunity. J. Clin. Investig. 2022, 132, e153167. [Google Scholar] [CrossRef]
- Hurtado, C.G.; Wan, F.; Housseau, F.; Sears, C.L. Roles for Interleukin 17 and Adaptive Immunity in Pathogenesis of Colorectal Cancer. Gastroenterology 2018, 155, 1706–1715. [Google Scholar] [CrossRef]
- Cui, G.; Yuan, A.; Goll, R.; Florholmen, J. IL-17A in the tumor microenvironment of the human colorectal adenoma–carcinoma sequence. Scand. J. Gastroenterol. 2012, 47, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- De Simone, V.; Pallone, F.; Monteleone, G.; Stolfi, C. Role of TH17 cytokines in the control of colorectal cancer. Oncoimmunology 2013, 2, e26617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Pan, F.; Guo, H.; Bu, F.; Xin, T.; Chen, S.; Guo, Y. Feedback mechanisms between M2 macrophages and Th17 cells in colorectal cancer patients. Tumor Biol. 2016, 37, 12223–12230. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yu, W.; Lian, J.; Wu, Q.; Liu, S.; Yang, L.; Li, F.; Huang, L.; Chen, X.; Zhang, Z.; et al. Th17 cells inhibit CD8+ T cell migration by systematically downregulating CXCR3 expression via IL-17A/STAT3 in advanced-stage colorectal cancer patients. J. Hematol. Oncol. 2020, 13, 1–15. [Google Scholar] [CrossRef]
- Amicarella, F.; Muraro, M.G.; Hirt, C.; Cremonesi, E.; Padovan, E.; Mele, V.; Governa, V.; Han, J.; Huber, X.; Droeser, R.A.; et al. Dual role of tumour-infiltrating T helper 17 cells in human colorectal cancer. Gut 2017, 66, 692–704. [Google Scholar] [CrossRef] [Green Version]
- Punkenburg, E.; Vogler, T.; Büttner, M.; Amann, K.; Waldner, M.; Atreya, R.; Abendroth, B.; Mudter, J.; Merkel, S.; Gallmeier, E.; et al. Batf-dependent Th17 cells critically regulate IL-23 driven colitis-associated colon cancer. Gut 2015, 65, 1139–1150. [Google Scholar] [CrossRef]
- Rizzo, A.; De Mare, V.; Rocchi, C.; Stolfi, C.; Colantoni, A.; Neurath, M.F.; Macdonald, T.T.; Pallone, F.; Monteleone, G.; Fantini, M.C. Smad7 induces plasticity in tumor-infiltrating Th17 cells and enables TNF-alpha-mediated killing of colorectal cancer cells. Carcinogenesis 2014, 35, 1536–1546. [Google Scholar] [CrossRef] [Green Version]
- Groth, C.; Weber, R.; Lasser, S.; Özbay, F.G.; Kurzay, A.; Petrova, V.; Altevogt, P.; Utikal, J.; Umansky, V. Tumor promoting capacity of polymorphonuclear myeloid-derived suppressor cells and their neutralization. Int. J. Cancer 2021, 149, 1628–1638. [Google Scholar] [CrossRef]
- Johdi, N.A.; Ait-Tahar, K.; Sagap, I.; Jamal, R. Molecular Signatures of Human Regulatory T Cells in Colorectal Cancer and Polyps. Front. Immunol. 2017, 8, 620. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.; Dalgleish, A.G.; Kumar, D. Review article: Immune suppression and colorectal cancer. Aliment. Pharmacol. Ther. 2006, 24, 1163–1177. [Google Scholar] [CrossRef]
- Giacomelli, L.; Gianni, W.; Belfiore, C.; Gandini, O.; Repetto, L.; Filippini, A.; Frati, L.; Aglianò, A.M.; Gazzaniga, P. Persistence of epidermal growth factor receptor and interleukin 10 in blood of colorectal cancer patients after surgery identifies patients with high risk to relapse. Clin. Cancer Res. 2003, 9, 2678–2682. [Google Scholar]
- Galizia, G.; Orditura, M.; Romano, C.; Lieto, E.; Castellano, P.; Pelosio, L.; Imperatore, V.; Catalano, G.; Pignatelli, C.; De Vita, F. Prognostic Significance of Circulating IL-10 and IL-6 Serum Levels in Colon Cancer Patients Undergoing Surgery. Clin. Immunol. 2002, 102, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Rossowska, J.; Anger, N.; Szczygieł, A.; Mierzejewska, J.; Pajtasz-Piasecka, E. Reprogramming the murine colon cancer microenvironment using lentivectors encoding shRNA against IL-10 as a component of a potent DC-based chemoimmunotherapy. J. Exp. Clin. Cancer Res. 2018, 37, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Huang, L.; Zhao, H.; Yan, Y.; Lu, J. The Role of Interleukins in Colorectal Cancer. Int. J. Biol. Sci. 2020, 16, 2323–2339. [Google Scholar] [CrossRef]
- Erdman, S.E.; Sohn, J.J.; Rao, V.P.; Nambiar, P.R.; Ge, Z.; Fox, J.G.; Schauer, D.B. CD4+CD25+ Regulatory Lymphocytes Induce Regression of Intestinal Tumors in ApcMin/+ Mice. Cancer Res. 2005, 65, 3998–4004. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, A.; Pallone, F.; Monteleone, G.; Fantini, M.C. Intestinal inflammation and colorectal cancer: A double-edged sword? World J. Gastroenterology. 2011, 17, 3092–3100. [Google Scholar]
- Li, J.; Xu, J.; Yan, X.; Jin, K.; Li, W.; Zhang, R. Targeting Interleukin-6 (IL-6) Sensitizes Anti-PD-L1 Treatment in a Colorectal Cancer Preclinical Model. Med Sci. Monit. 2018, 24, 5501–5508. [Google Scholar] [CrossRef]
- Koper-Lenkiewicz, O.; Dymicka-Piekarska, V.; Milewska, A.; Zińczuk, J.; Kamińska, J. The Relationship between Inflammation Markers (CRP, IL-6, sCD40L) and Colorectal Cancer Stage, Grade, Size and Location. Diagnostics 2021, 11, 1382. [Google Scholar] [CrossRef]
- Lehtomäki, K.; Mustonen, H.; Kellokumpu-Lehtinen, P.-L.; Joensuu, H.; Hermunen, K.; Soveri, L.-M.; Boisen, M.; Dehlendorff, C.; Johansen, J.; Haglund, C.; et al. Lead Time and Prognostic Role of Serum CEA, CA19-9, IL-6, CRP, and YKL-40 after Adjuvant Chemotherapy in Colorectal Cancer. Cancers 2021, 13, 3892. [Google Scholar] [CrossRef]
- Lu, C.-C.; Kuo, H.-C.; Wang, F.-S.; Jou, M.-H.; Lee, K.-C.; Chuang, J.-H. Upregulation of TLRs and IL-6 as a Marker in Human Colorectal Cancer. Int. J. Mol. Sci. 2014, 16, 159–177. [Google Scholar] [CrossRef] [Green Version]
- Ohno, Y.; Toyoshima, Y.; Yurino, H.; Monma, N.; Xiang, H.; Sumida, K.; Kaneumi, S.; Terada, S.; Hashimoto, S.; Ikeo, K.; et al. Lack of interleukin-6 in the tumor microenvironment augments type-1 immunity and increases the efficacy of cancer immunotherapy. Cancer Sci. 2017, 108, 1959–1966. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wu, Z.; Meng, W.; Zhang, C.; Cheng, M.; Chen, Y.; Zou, Y.; Li, K.; Lin, S.; Xiong, W.; et al. Blockade of IL-6 inhibits tumor immune evasion and improves anti–PD-1 immunotherapy. Cytokine 2022, 158, 155976. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Fantini, M.C.; Schramm, C.; Lehr, H.A.; Wirtz, S.; Becker, A.; Burg, J.; Strand, S.; Kiesslich, R.; Huber, S.; et al. TGF-beta suppresses tumor progression in colon cancer by inhibition of IL–6 trans-signaling. Immunity 2004, 42, 139. [Google Scholar] [CrossRef]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2017, 18, 1271. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, K.; Popp, V.; Wirtz, S.; Al-Saifi, R.; Acera, M.G.; Atreya, R.; Dregelies, T.; Vieth, M.; Fichtner-Feigl, S.; McKenzie, A.N.J.; et al. PU.1-driven Th9 Cells Promote Colorectal Cancer in Experimental Colitis Models Through Il-6 Effects in Intestinal Epithelial Cells. J. Crohn’s Colitis 2022. [Google Scholar] [CrossRef]
- Neurath, M.F.; Finotto, S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor Rev. 2011, 22, 83–89. [Google Scholar] [CrossRef]
- Wunderlich, C.M.; Ackermann, P.J.; Ostermann, A.L.; Adams-Quack, P.; Vogt, M.C.; Tran, M.L.; Nikolajev, A.; Waisman, A.; Garbers, C.; Theurich, S.; et al. Obesity exacerbates colitis-associated cancer via IL-6-regulated macrophage polarisation and CCL-20/CCR-6-mediated lymphocyte recruitment. Nat. Commun. 2018, 9, 1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, C.; Wang, L.; Hu, S.; Huang, C.; Xia, Z.; Liao, J.; Yi, W.; Chen, J. Poly(I:C) enhances the efficacy of phagocytosis checkpoint blockade immunotherapy by inducing IL-6 production. J. Leukoc. Biol. 2021, 110, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.-C.; Zhang, Z.; Li, T.-Y.; Liang, Y.-F.; Wang, H.-M.; Bao, J.-J.; Zhang, J.-A.; Wang, W.-D.; Xiang, W.-Y.; Kong, B.; et al. Assessing the role of IL-35 in colorectal cancer progression and prognosis. Int. J. Clin. Exp. Pathol. 2013, 6, 1806–1816. [Google Scholar]
- Wang, Z.; Liu, J.-Q.; Liu, Z.; Shen, R.; Zhang, G.; Xu, J.; Basu, S.; Feng, Y.; Bai, X.-F. Tumor-Derived IL-35 Promotes Tumor Growth by Enhancing Myeloid Cell Accumulation and Angiogenesis. J. Immunol. 2013, 190, 2415–2423. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.A.; Neurath, M.F. Anticancer properties of the IL-12 family--focus on colorectal cancer. Curr. Med. Chem. 2010, 17, 3303–3308. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Kuchroo, V.K. IL-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, H.; Clark, S.C.; Hamaoka, T. Cellular and Molecular Mechanisms Underlying IL-12-Induced Tumor Regression. Ann. New York Acad. Sci. 1996, 795, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Yang, P.; Zhang, E.; Gu, J.; Xu, H.; Li, M.; Gao, X.; Li, X.; Zhang, Y.; Xu, H.; et al. Significantly increased anti-tumor activity of carcinoembryonic antigen-specific chimeric antigen receptor T cells in combination with recombinant human IL-12. Cancer Med. 2019, 8, 4753–4765. [Google Scholar] [CrossRef]
- Uemura, A.; Takehara, T.; Miyagi, T.; Suzuki, T.; Tatsumi, T.; Ohkawa, K.; Kanto, T.; Hiramatsu, N.; Hayashi, N. Natural killer cell is a major producer of interferon gamma that is critical for the IL-12-induced anti-tumor effect in mice. Cancer Immunol. Immunother. CII 2010, 59, 453–463. [Google Scholar] [CrossRef]
- Rodolfo, M.; Zilocchi, C.; Melani, C.; Cappetti, B.; Arioli, I.; Parmiani, G.; Colombo, M.P. Immunotherapy of experimental metastases by vaccination with interleukin gene-transduced adenocarcinoma cells sharing tumor-associated antigens. Comparison between IL-12 and IL-2 gene-transduced tumor cell vaccines. J. Immunol. 1996, 157, 5536–5542. [Google Scholar]
- Tannenbaum, C.S.; Wicker, N.; Armstrong, D.; Tubbs, R.; Finke, J.; Bukowski, R.M.; A Hamilton, T. Cytokine and chemokine expression in tumors of mice receiving systemic therapy with IL-12. J. Immunol. 1996, 156, 693–699. [Google Scholar]
- Nastala, C.L.; Edington, H.D.; McKinney, T.G.; Tahara, H.; A Nalesnik, M.; Brunda, M.J.; Gately, M.K.; Wolf, S.F.; Schreiber, R.D.; Storkus, W.J. Recombinant IL-12 administration induces tumor regression in association with IFN-gamma production. J. Immunol. 1994, 153, 1697–1706. [Google Scholar]
- Liu, X.; Gao, X.; Zheng, S.; Wang, B.; Li, Y.; Zhao, C.; Muftuoglu, Y.; Chen, S.; Li, Y.; Yao, H.; et al. Modified nanoparticle mediated IL-12 immunogene therapy for colon cancer. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1993–2004. [Google Scholar] [CrossRef]
- Rodriguez-Madoz, J.R.; Prieto, J.; Smerdou, C. Semliki forest virus vectors engineered to express higher IL-12 levels induce efficient elimination of murine colon adenocarcinomas. Mol. Ther. 2005, 12, 153–163. [Google Scholar] [CrossRef]
- Quetglas, J.I.; Rodriguez-Madoz, J.R.; Bezunartea, J.; Ruiz-Guillen, M.; Casales, E.; Medina-Echeverz, J.; Prieto, J.; Berraondo, P.; Hervas-Stubbs, S.; Smerdou, C. Eradication of liver-implanted tumors by Semliki Forest virus expressing IL-12 requires efficient long-term immune responses. J. Immunol. 2013, 190, 2994–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gri, G.; Chiodoni, C.; Gallo, E.; Stoppacciaro, A.; Liew, F.Y.; Colombo, M.P. Antitumor effect of interleukin (IL)-12 in the absence of endogenous IFN-gamma: A role for intrinsic tumor immunogenicity and IL-15. Cancer Res. 2002, 62, 4390–4397. [Google Scholar]
- Malvicini, M.; Ingolotti, M.; Piccioni, F.; Garcia, M.; Bayo, J.; Atorrasagasti, C.; Alaniz, L.; Aquino, J.B.; Espinoza, J.A.; Gidekel, M.; et al. Reversal of gastrointestinal carcinoma-induced immunosuppression and induction of antitumoural immunity by a combination of cyclophosphamide and gene transfer of IL-12. Mol. Oncol. 2011, 5, 242–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.X.; Gao, X.; Janakiraman, N.; Chapman, R.A.; Gautam, S.C. IL-12 Gene Therapy of Leukemia with Hematopoietic Progenitor Cells without the Toxicity of Systemic IL-12 Treatment. Clin. Immunol. 2001, 98, 180–189. [Google Scholar] [CrossRef]
- Lenzi, R.; Rosenblum, M.; Verschraegen, C.; Kudelka, A.P.; Kavanagh, J.J.; Hicks, M.E.; Lang, E.A.; Nash, M.A.; Levy, L.B.; Garcia, M.E.; et al. Phase I study of intraperitoneal recombinant human interleukin 12 in patients with Mullerian carcinoma, gastrointestinal primary malignancies, and mesothelioma. Clin. Cancer Res. 2002, 8, 3686–3695. [Google Scholar]
- Hewitt, S.L.; Bailey, D.; Zielinski, J.; Apte, A.; Musenge, F.; Karp, R.; Burke, S.; Garcon, F.; Mishra, A.; Gurumurthy, S.; et al. Intratumoral IL12 mRNA Therapy Promotes TH1 Transformation of the Tumor Microenvironment. Clin. Cancer Res. 2020, 26, 6284–6298. [Google Scholar] [CrossRef] [PubMed]
- Chiyo, M.; Shimozato, O.; Yu, L.; Kawamura, K.; Iizasa, T.; Fujisawa, T.; Tagawa, M. Expression of IL-27 in murine carcinoma cells produces antitumor effects and induces protective immunity in inoculated host animals. Int. J. Cancer 2005, 115, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Morishima, N.; Owaki, T.; Asakawa, M.; Kamiya, S.; Mizuguchi, J.; Yoshimoto, T. Augmentation of Effector CD8+ T Cell Generation with Enhanced Granzyme B Expression by IL-27. J. Immunol. 2005, 175, 1686–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, N.; Canavan, J.B.; Macdonald, T.T.; Lord, G. Transcriptional regulation of the mucosal immune system mediated by T-bet. Mucosal Immunol. 2010, 3, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Atreya, I.; Schimanski, C.C.; Becker, C.; Wirtz, S.; Dornhoff, H.; Schnürer, E.; Berger, M.R.; Galle, P.R.; Herr, W.; Neurath, M.F. The T-box transcription factor eomesodermin controls CD8 T cell activity and lymph node metastasis in human colorectal cancer. Gut 2007, 56, 1572–1578. [Google Scholar] [CrossRef] [Green Version]
- Neurath, M.F. IL-23 in inflammatory bowel diseases and colon cancer. Cytokine Growth Factor Rev. 2018, 45, 1–8. [Google Scholar] [CrossRef]
- Richter, C.; Herrero San Juan, M.; Weigmann, B.; Bergis, D.; Dauber, K.; Muders, M.H.; Baretton, G.B.; Pfeilschifter, J.M.; Bonig, H.; Brenner, S.; et al. Defective IL-23/IL-17 Axis Protects p47phox-/- Mice from Colon Cancer. Front. Immunol. 2017, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, F.; Zhang, L.; Wu, J.; Zhang, J.; Zhang, S.; Li, K.; Qi, Y.; Lin, P. IL-23/IL-23R: Potential mediator of intestinal tumor progression from adenomatous polyps to colorectal carcinoma. Int. J. Color. Dis. 2011, 26, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, J.; Madka, V.; Rai, R.; Morris, K.T.; Houchen, C.W.; Chandrakesan, P.; Rao, C.V. Inflammatory Mediators and Gut Microbial Toxins Drive Colon Tumorigenesis by IL-23 Dependent Mechanism. Cancers 2021, 13, 5159. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.Y.; Osterreicher, C.H.; Hung, K.E.; et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature 2012, 491, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Kvedaraite, E.; Lourda, M.; Ideström, M.; Chen, P.; Olsson-Åkefeldt, S.; Forkel, M.; Gavhed, D.; Lindforss, U.; Mjösberg, J.; Henter, J.-I.; et al. Tissue-infiltrating neutrophils represent the main source of IL-23 in the colon of patients with IBD. Gut 2015, 65, 1632–1641. [Google Scholar] [CrossRef]
- Zhou, L.; Ivanov, I.I.; Spolski, R.; Min, R.; Shenderov, K.; Egawa, T.; Levy, D.E.; Leonard, W.J.; Littman, D.R. IL-6 programs T(H)-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat. Immunol. 2007, 8, 967–974. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Chen, Y.; Tato, C.M.; Laurence, A.; Joyce-Shaikh, B.; Blumenschein, W.M.; McClanahan, T.K.; O’Shea, J.J.; Cua, D.J. The interleukin 23 receptor is essential for the terminal differentiation of interleukin 17–producing effector T helper cells in vivo. Nat. Immunol. 2009, 10, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Sawant, D.V.; Yano, H.; Chikina, M.; Zhang, Q.; Liao, M.; Liu, C.; Callahan, D.J.; Sun, Z.; Sun, T.; Tabib, T.; et al. Adaptive plasticity of IL-10+ and IL-35+ Treg cells cooperatively promotes tumor T cell exhaustion. Nat. Immunol. 2019, 20, 724–735. [Google Scholar] [CrossRef]
- Liu, K.; Huang, A.; Nie, J.; Tan, J.; Xing, S.; Qu, Y.; Jiang, K. IL-35 Regulates the Function of Immune Cells in Tumor Microenvironment. Front. Immunol. 2021, 12, 683332. [Google Scholar] [CrossRef]
- Tauriello, D.V.F.; Palomo-Ponce, S.; Stork, D.; Berenguer-Llergo, A.; Badia-Ramentol, J.; Iglesias, M.; Sevillano, M.; Ibiza, S.; Cañellas, A.; Hernando-Momblona, X.; et al. TGFbeta drives immune evasion in genetically reconstituted colon cancer metastasis. Nature 2018, 554, 538–543. [Google Scholar] [CrossRef]
- Stolfi, C.; Troncone, E.; Marafini, I.; Monteleone, G. Role of TGF-Beta and Smad7 in Gut Inflammation, Fibrosis and Cancer. Biomolecules 2020, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Owyang, S.Y.; Zhang, M.; Walkup, G.A.; Chen, G.E.; Grasberger, H.; El-Zaatari, M.; Kao, J.Y. The effect of CT26 tumor-derived TGF-beta on the balance of tumor growth and immunity. Immunol. Lett. 2017, 191, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Otegbeye, F.; Ojo, E.; Moreton, S.; Mackowski, N.; Lee, D.A.; de Lima, M.; Wald, D.N. Inhibiting TGF-beta signaling preserves the function of highly activated, in vitro expanded natural killer cells in AML and colon cancer models. PLoS ONE 2018, 13, e0191358. [Google Scholar]
- Fantini, M.C.; Becker, C.; Tubbe, I.; Nikolaev, A.; Lehr, H.A.; Galle, P.; Neurath, M.F. Transforming growth factor beta induced FoxP3+ regulatory T cells suppress Th1 mediated experimental colitis. Gut 2006, 55, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Gneo, L.; Rizkalla, N.; Hejmadi, R.; Mussai, F.; de Santo, C.; Middleton, G. TGF-beta orchestrates the phenotype and function of monocytic myeloid-derived suppressor cells in colorectal cancer. Cancer Immunol. Immunother. CII 2022, 71, 1583–1596. [Google Scholar] [CrossRef]
- Becker, C.; Fantini, M.; Neurath, M. TGF-beta as a T cell regulator in colitis and colon cancer. Cytokine Growth Factor Rev. 2006, 17, 97–106. [Google Scholar] [CrossRef]
- Horn, L.A.; Chariou, P.L.; Gameiro, S.R.; Qin, H.; Iida, M.; Fousek, K.; Meyer, T.J.; Cam, M.; Flies, D.; Langermann, S.; et al. Remodeling the tumor microenvironment via blockade of LAIR-1 and TGF-beta signaling enables PD-L1-mediated tumor eradication. J. Clin. Investig. 2022, 132, e155148. [Google Scholar] [CrossRef]
- Calon, A.; Lonardo, E.; Berenguer-Llergo, A.; Espinet, E.; Hernando-Momblona, X.; Iglesias, M.; Sevillano, M.; Palomo-Ponce, S.; Tauriello, D.V.F.; Byrom, D.; et al. Stromal gene expression defines poor-prognosis subtypes in colorectal cancer. Nat. Genet. 2015, 47, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Perez, L.G.; Kempski, J.; McGee, H.M.; Pelzcar, P.; Agalioti, T.; Giannou, A.; Konczalla, L.; Brockmann, L.; Wahib, R.; Xu, H.; et al. TGF-beta signaling in Th17 cells promotes IL-22 production and colitis-associated colon cancer. Nat. Commun. 2020, 11, 2608. [Google Scholar] [CrossRef]
- Zhuang, Y.; Peng, L.-S.; Zhao, Y.-L.; Shi, Y.; Mao, X.-H.; Guo, G.; Chen, W.; Liu, X.-F.; Zhang, J.-Y.; Liu, T.; et al. Increased intratumoral IL-22-producing CD4+ T cells and Th22 cells correlate with gastric cancer progression and predict poor patient survival. Cancer Immunol. Immunother. 2012, 61, 1965–1975. [Google Scholar] [CrossRef] [PubMed]
- Dornhoff, H.; Becker, C.; Wirtz, S.; Strand, D.; Tenzer, S.; Rosfa, S.; Neufert, C.; Mudter, J.; Markl, J.; Siebler, J.; et al. A variant of Smurf2 protects mice against colitis-associated colon cancer by inducing transforming growth factor beta signaling. Gastroenterology 2012, 142, 1183–1194.e4. [Google Scholar] [CrossRef] [PubMed]
- Malla, R.R.; Marni, R. Angiogenesis: Promising therapeutic target of metastatic colon cancer. Crit. Rev. Oncog. 2020, 25, 161–173. [Google Scholar] [CrossRef]
- Kampoli, K.; Foukas, P.G.; Ntavatzikos, A.; Arkadopoulos, N.; Koumarianou, A. Interrogating the interplay of angiogenesis and immunity in metastatic colorectal cancer. World J. Methodol. 2022, 12, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Voron, T.; Marcheteau, E.; Pernot, S.; Colussi, O.; Tartour, E.; Taieb, J.; Terme, M. Control of the immune response by pro-angiogenic factors. Front Oncol. 2014, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Terme, M.; Pernot, S.; Marcheteau, E.; Sandoval, F.; Benhamouda, N.; Colussi, O.; Dubreuil, O.; Carpentier, A.F.; Tartour, E.; Taieb, J. VEGFA-VEGFR Pathway Blockade Inhibits Tumor-Induced Regulatory T-cell Proliferation in Colorectal Cancer. Cancer Res. 2013, 73, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhu, Y.; Li, C.; Trinh, R.; Ren, X.; Sun, F.; Wang, Y.; Shang, P.; Wang, T.; Wang, M.; et al. Anti-VEGFR2-interferon-α2 regulates the tumor microenvironment and exhibits potent antitumor efficacy against colorectal cancer. OncoImmunology 2017, 6, e1290038. [Google Scholar] [CrossRef]
- Shang, P.; Gao, R.; Zhu, Y.; Zhang, X.; Wang, Y.; Guo, M.; Peng, H.; Wang, M.; Zhang, J. VEGFR2-targeted antibody fused with IFN mut regulates the tumor microenvironment of colorectal cancer and exhibits potent anti-tumor and anti-metastasis activity. Acta Pharm. Sin. B 2020, 11, 420–433. [Google Scholar] [CrossRef]
- Zundler, S.; Becker, E.; Weidinger, C.; Siegmund, B. Anti-Adhesion Therapies in Inflammatory Bowel Disease—Molecular and Clinical Aspects. Front. Immunol. 2017, 8, 891. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xie, R.; Zhang, H.; Zheng, Y.; Lin, C.; Yang, L.; Huang, M.; Li, M.; Song, F.; Lu, L.; et al. Integrin beta7 Inhibits Colorectal Cancer Pathogenesis via Maintaining Antitumor Immunity. Cancer Immunol. Res. 2021, 9, 967–980. [Google Scholar] [CrossRef]
- Das, S.; Donas, C.; Akeus, P.; Quiding-Jarbrink, M.; Mora, J.R.; Villablanca, E.J. beta7 integrins contribute to intestinal tumor growth in mice. PLoS ONE 2018, 13, e0204181. [Google Scholar] [CrossRef] [PubMed]
- Wiendl, M.; Becker, E.; Müller, T.M.; Voskens, C.J.; Neurath, M.F.; Zundler, S. Targeting Immune Cell Trafficking—Insights From Research Models and Implications for Future IBD Therapy. Front. Immunol. 2021, 12, 1546. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atreya, I.; Neurath, M.F. How the Tumor Micromilieu Modulates the Recruitment and Activation of Colorectal Cancer-Infiltrating Lymphocytes. Biomedicines 2022, 10, 2940. https://doi.org/10.3390/biomedicines10112940
Atreya I, Neurath MF. How the Tumor Micromilieu Modulates the Recruitment and Activation of Colorectal Cancer-Infiltrating Lymphocytes. Biomedicines. 2022; 10(11):2940. https://doi.org/10.3390/biomedicines10112940
Chicago/Turabian StyleAtreya, Imke, and Markus F. Neurath. 2022. "How the Tumor Micromilieu Modulates the Recruitment and Activation of Colorectal Cancer-Infiltrating Lymphocytes" Biomedicines 10, no. 11: 2940. https://doi.org/10.3390/biomedicines10112940