
Vitamin E and Non-Communicable Diseases: A Review
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Metabolism
3. Serum Metabolites
4. Vitamin E Effects in Human Health and Diseases
4.1. Antioxidant Activity
4.2. Anti-Inflammatory Activity
4.3. Interaction of Vitamin E and Gut Microbiota
4.4. Vitamin E and Cardiovascular Diseases
4.5. Vitamin E and Diabetes Mellitus
4.6. Vitamin E and Asthma
4.7. Vitamin E and Cancer
5. Toxicity and Health Risk of Vitamin E
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evans, H.M.; Bishop, K.S. On the existence of a hitherto unrecognized dietary factor essential for reproduction. Science 1922, 56, 650–651. [Google Scholar] [CrossRef] [Green Version]
- Niki, E.; Abe, K. Vitamin E: Structure, Properties and Functions. In Vitamin E: Chemistry and Nutritional Benefits, 1st ed.; Niki, E., Ed.; Royal Society of Chemistry: London, UK, 2019; Volume 11, pp. 1–11. [Google Scholar]
- Jiang, Q. Metabolism of natural forms of vitamin E and biological actions of vitamin E metabolites. Free Radic. Biol. Med. 2022, 179, 375–387. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Torquato, P.; Ripa, O.; Giusepponi, D.; Galarini, R.; Bartolini, D.; Wallert, M.; Pellegrino, R.; Cruciani, G.; Lorkowski, S.; Birringer, M.; et al. Analytical strategies to assess the functional metabolome of vitamin E. J. Pharm. Biomed. Anal. 2016, 124, 399–412. [Google Scholar] [CrossRef]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory Redox Interactions. IUBMB Life 2019, 71, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Kluge, S.; Schmölz, L.; Wallert, M.; Galli, F.; Birringer, M.; Lorkowski, S. Long-Chain Metabolites of Vitamin E: Metabolic Activation as a General Concept for Lipid-Soluble Vitamins? Antioxidants 2018, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Ciffolilli, S.; Wallert, M.; Bartolini, D.; Krauth, V.; Werz, O.; Piroddi, M.; Sebastiani, B.; Torquato, P.; Lorkowski, S.; Birringer, M.; et al. Human serum determination and in vitro anti-inflammatory activity of the vitamin E metabolite α-(13’-hydroxy)-6-hydroxychroman. Free Radic. Biol. Med. 2015, 89, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Graf, P.; Brigelius-Flohé, R.; Wechter, W.; Sies, H. Quantification of the alpha- and gamma-tocopherol metabolites 2,5,7, 8-tetramethyl-2-(2’-carboxyethyl)-6-hydroxychroman and 2,7, 8-trimethyl-2-(2’-carboxyethyl)-6-hydroxychroman in human serum. Anal. Biochem. 1999, 275, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Han, S.N. Vitamin E: Regulatory role on gene and protein expression and metabolomics profiles. IUBMB Life 2019, 71, 442–455. [Google Scholar] [CrossRef] [PubMed]
- Sozen, E.; Demirel, T.; Ozer, N.K. Vitamin E: Regulatory role in the cardiovascular system. IUBMB Life 2019, 71, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Wallert, M.; Mosig, S.; Rennert, K.; Funke, H.; Ristow, M.; Pellegrino, R.M.; Cruciani, G.; Galli, F.; Lorkowski, S.; Birringer, M. Long-chain metabolites of α-tocopherol occur in human serum and inhibit macrophage foam cell formation in vitro. Free Radic. Biol. Med. 2014, 68, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Martens, L.G.; Luo, J.; Meulmeester, F.L.; Ashrafi, N.; van Eekelen, E.W.; de Mutsert, R.; Mook-Kanamori, D.O.; Rosendaal, F.R.; van Dijk, K.W.; Mills, K.; et al. Associations between Lifestyle Factors and Vitamin E Metabolites in the General Population. Antioxidants 2020, 9, 1280. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Fasciolo, G.; Di Meo, S.; Venditti, P. Vitamin E Supplementation and Mitochondria in Experimental and Functional Hyperthyroidism: A Mini-Review. Nutrients 2019, 11, 2900. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.Y.; Han, S.N. The Role of Vitamin E in Immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Khadangi, F.; Azzi, A. Vitamin E—The Next 100 Years. IUBMB Life 2019, 71, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.D.; Meydani, S.N.; Wu, D. Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 2019, 71, 487–494. [Google Scholar] [CrossRef]
- Pirhadi-Tavandashti, N.; Imani, H.; Ebrahimpour-Koujan, S.; Samavat, S.; Hakemi, M.S. The effect of vitamin E supplementation on biomarkers of endothelial function and inflammation among hemodialysis patients: A double-blinded randomized clinical trial. Complement. Ther. Med. 2020, 49, 102357. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Kattoor, A.J.; Saldeen, T.; Mehta, J.L. Vitamin E and its anticancer effects. Crit. Rev. Food Sci. Nutr. 2019, 59, 2831–2838. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar] [PubMed]
- Gao, B.; Chi, L.; Mahbub, R.; Bian, X.; Tu, P.; Ru, H.; Lu, K. Multi-Omics Reveals that Lead Exposure Disturbs Gut Microbiome Development, Key Metabolites, and Metabolic Pathways. Chem. Res. Toxicol. 2017, 30, 996–1005. [Google Scholar] [CrossRef] [Green Version]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [Green Version]
- Stacchiotti, V.; Rezzi, S.; Eggersdorfer, M.; Galli, F. Metabolic and functional interplay between gut microbiota and fat-soluble vitamins. Crit. Rev. Food Sci. Nutr. 2021, 61, 3211–3232. [Google Scholar] [CrossRef]
- Walker, W.A. Dysbiosis. In The Microbiota in Gastrointestinal Pathophysiology, 1st ed.; Floch, M.H., Yehuda Ringel, Y., Walker, W.A., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 227–232. [Google Scholar]
- Pham, V.T.; Fehlbaum, S.; Seifert, N.; Richard, N.; Bruins, M.J.; Sybesma, W.; Rehman, A.; Steinert, R.E. Effects of colon-targeted vitamins on the composition and metabolic activity of the human gut microbiome—A pilot study. Gut Microbes 2021, 13, 1875774. [Google Scholar] [CrossRef]
- Vernia, F.; Longo, S.; Stefanelli, G.; Viscido, A.; Latella, G. Dietary Factors Modulating Colorectal Carcinogenesis. Nutrients 2021, 13, 143. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.; Kim, S.; Lee, J.; Ha, J.; Oh, H.; Lee, Y.; Kim, Y.; Yoon, Y. Vitamin E (α-tocopherol) consumption influences gut microbiota composition. Int. J. Food Sci. Nutr. 2020, 71, 221–225. [Google Scholar] [CrossRef]
- Liu, K.Y.; Nakatsu, C.H.; Jones-Hall, Y.; Kozik, A.; Jiang, Q. Vitamin E alpha- and gamma-tocopherol mitigate colitis, protect intestinal barrier function and modulate the gut microbiota in mice. Free Radic. Biol. Med. 2021, 163, 180–189. [Google Scholar] [CrossRef]
- Ran, L.; Liu, A.B.; Lee, M.J.; Xie, P.; Lin, Y.; Yang, C.S. Effects of antibiotics on degradation and bioavailability of different vitamin E forms in mice. Biofactors 2019, 45, 450–462. [Google Scholar] [CrossRef]
- Traber, M.G.; Buettner, G.R.; Bruno, R.S. The relationship between vitamin C status, the gut-liver axis, and metabolic syndrome. Redox Biol. 2019, 21, 101091. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Xie, J.; Bai, H.; Tian, T.; Zou, T.; Chen, J.J. Gut Microbiota-Derived Inflammation-Related Serum Metabolites as Potential Biomarkers for Major Depressive Disorder. J. Inflamm. Res. 2021, 14, 3755–3766. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhao, Y.; Im, S.; Nakatsu, C.; Jones-Hall, Y.; Jiang, Q. Vitamin E delta-tocotrienol and metabolite 13’-carboxychromanol inhibit colitis-associated colon tumorigenesis and modulate gut microbiota in mice. J. Nutr. Biochem. 2021, 89, 108567. [Google Scholar] [CrossRef] [PubMed]
- Wallert, M.; Ziegler, M.; Wang, X.; Maluenda, A.; Xu, X.; Yap, M.L.; Witt, R.; Giles, C.; Kluge, S.; Hortmann, M.; et al. α-Tocopherol preserves cardiac function by reducing oxidative stress and inflammation in ischemia/reperfusion injury. Redox Biol. 2019, 26, 101292. [Google Scholar] [CrossRef] [PubMed]
- Wallert, M.; Börmel, L.; Lorkowski, S. Inflammatory Diseases and Vitamin E-What Do We Know and Where Do We Go? Mol. Nutr. Food Res. 2021, 65, e2000097. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. S1), S67–S74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 17 August 2022).
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk factors contributing to type 2 diabetes and recent advances in the treatment and prevention. Int. J. Med. Sci. 2014, 11, 1185–1200. [Google Scholar] [CrossRef] [Green Version]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxid. Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef] [Green Version]
- Balbi, M.E.; Tonin, F.S.; Mendes, A.M.; Borba, H.H.; Wiens, A.; Fernandez-Llimos, F.; Pontarolo, R. Antioxidant effects of vitamins in type 2 diabetes: A meta-analysis of randomized controlled trials. Diabetol. Metab. Syndr. 2018, 10, 18. [Google Scholar] [CrossRef]
- Kataja-Tuomola, M.; Sundell, J.R.; Männistö, S.; Virtanen, M.J.; Kontto, J.; Albanes, D.; Virtamo, J. Effect of alpha-tocopherol and beta-carotene supplementation on the incidence of type 2 diabetes. Diabetologia 2008, 51, 47–53. [Google Scholar] [CrossRef]
- Mayer-Davis, E.J.; Costacou, T.; King, I.; Zaccaro, D.J.; Bell, R.A. Insulin Resistance and Atherosclerosis Study (IRAS). Plasma and dietary vitamin E in relation to incidence of type 2 diabetes: The Insulin Resistance and Atherosclerosis Study (IRAS). Diabetes Care 2002, 25, 2172–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Lee, I.M.; Song, Y.; Van Denburgh, M.; Cook, N.R.; Manson, J.E.; Buring, J.E. Vitamin E and risk of type 2 diabetes in the women’s health study randomized controlled trial. Diabetes 2006, 55, 2856–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazdro, R.; Burgess, J.R. The role of vitamin E and oxidative stress in diabetes complications. Mech. Ageing. Dev. 2010, 131, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Shirai, Y.; Matsubara, T.; Sanse, K.; Kuriyama, M.; Oshiro, N.; Yoshino, K.; Yonezawa, K.; Ono, Y.; Saito, N. Phosphorylation and up-regulation of diacylglycerol kinase gamma via its interaction with protein kinase C gamma. J. Biol. Chem. 2006, 281, 31627–31637. [Google Scholar] [CrossRef]
- Poznyak, A.; Grechko, A.V.; Poggio, P.; Myasoedova, V.A.; Alfieri, V.; Orekhov, A.N. The Diabetes Mellitus-Atherosclerosis Connection: The Role of Lipid and Glucose Metabolism and Chronic Inflammation. Int. J. Mol. Sci. 2020, 21, 1835. [Google Scholar] [CrossRef] [Green Version]
- Bursell, S.E.; King, G.L. Can protein kinase C inhibition and vitamin E prevent the development of diabetic vascular complications? Diabetes Res. Clin. Pract. 1999, 45, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, S.; Jialal, I. Alpha tocopherol supplementation decreases serum C-reactive protein and monocyte interleukin-6 levels in normal volunteers and type 2 diabetic patients. Free Radic. Biol. Med. 2000, 29, 790–792. [Google Scholar] [CrossRef]
- Xu, Y.J.; Tappia, P.S.; Neki, N.S.; Dhalla, N.S. Prevention of diabetes-induced cardiovascular complications upon treatment with antioxidants. Heart Fail. Rev. 2014, 19, 113–121. [Google Scholar] [CrossRef]
- Samsu, N. Diabetic Nephropathy: Challenges in Pathogenesis, Diagnosis, and Treatment. Biomed. Res. Int. 2021, 2021, 1497449. [Google Scholar] [CrossRef]
- Khatami, P.G.; Soleimani, A.; Sharifi, N.; Aghadavod, E.; Asemi, Z. The effects of high-dose vitamin E supplementation on biomarkers of kidney injury, inflammation, and oxidative stress in patients with diabetic nephropathy: A randomized, double-blind, placebo-controlled trial. J. Clin. Lipidol. 2016, 10, 922–929. [Google Scholar] [CrossRef]
- Tan, G.C.J.; Tan, S.M.Q.; Phang, S.C.W.; Ng, Y.T.; Ng, E.Y.; Ahmad, B.; Palamisamy, U.D.M.; Kadir, K.A. Tocotrienol-rich vitamin E improves diabetic nephropathy and persists 6-9 months after washout: A phase IIa randomized controlled trial. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018819895462. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wang, P.; Airen, S.; Brown, C.; Liu, Z.; Townsend, J.H.; Wang, J.; Jiang, H. Nutritional and medical food therapies for diabetic retinopathy. Eye Vis. 2020, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei-Malazy, O.; Ardeshirlarijani, E.; Namazi, N.; Nikfar, S.; Jalili, R.B.; Larijani, B. Dietary antioxidative supplements and diabetic retinopathy; a systematic review. J. Diabetes Metab. Disord. 2019, 18, 705–716, Erratum in J. Diabetes Metab. Disord. 2019, 18, 717. [Google Scholar] [CrossRef] [PubMed]
- She, C.; Shang, F.; Cui, M.; Yang, X.; Liu, N. Association between dietary antioxidants and risk for diabetic retinopathy in a Chinese population. Eye 2021, 35, 1977–1984. [Google Scholar] [CrossRef]
- Ho, J.I.; Ng, E.Y.; Chiew, Y.; Koay, Y.Y.; Chuar, P.F.; Phang, S.C.W.; Ahmad, B.; Kadir, K.A. The effects of vitamin E on non-proliferative diabetic retinopathy in type 2 diabetes mellitus: Are they sustainable with 12 months of therapy. SAGE Open Med. 2022, 10, 20503121221095324. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Silveira, J.S.; Antunes, G.L.; Kaiber, D.B.; da Costa, M.S.; Marques, E.P.; Ferreira, F.S.; Gassen, R.B.; Breda, R.V.; Wyse, A.T.S.; Pitrez, P.; et al. Reactive oxygen species are involved in eosinophil extracellular traps release and in airway inflammation in asthma. J. Cell. Physiol. 2019, 234, 23633–23646. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, C.; Wang, Y.; Li, Y. Does vitamin E prevent asthma or wheeze in children: A systematic review and meta-analysis. Paediatr. Respir. Rev. 2018, 27, 60–68. [Google Scholar] [CrossRef]
- Devereux, G.; Craig, L.; Seaton, A.; Turner, S. Maternal vitamin D and E intakes in pregnancy and asthma to age 15 years: A cohort study. Pediatr. Pulmonol. 2019, 54, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Alwarith, J.; Kahleova, H.; Crosby, L.; Brooks, A.; Brandon, L.; Levin, S.M.; Barnard, N.D. The role of nutrition in asthma prevention and treatment. Nutr. Rev. 2020, 78, 928–938. [Google Scholar] [CrossRef]
- Strait, R.T.; Camargo, C.A. Vitamin E and the risk of childhood asthma. Expert Rev. Respir. Med. 2016, 10, 881–890. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 19 August 2022).
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.; Jiang, X.; Ben, S.; Yuan, Q.; Su, L.; Zhang, Z.; Christiani, D.C.; Du, M.; Wang, M. Association between circulating vitamin E and ten common cancers: Evidence from large-scale Mendelian randomization analysis and a longitudinal cohort study. BMC Med. 2022, 20, 168. [Google Scholar] [CrossRef]
- Fernandez-Lazaro, C.I.; Martínez-González, M.Á.; Aguilera-Buenosvinos, I.; Gea, A.; Ruiz-Canela, M.; Romanos-Nanclares, A.; Toledo, E. Dietary Antioxidant Vitamins and Minerals and Breast Cancer Risk: Prospective Results from the SUN Cohort. Antioxidants 2021, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Playdon, M.C.; Ziegler, R.G.; Sampson, J.N.; Stolzenberg-Solomon, R.; Thompson, H.J.; Irwin, M.L.; Mayne, S.T.; Hoover, R.N.; Moore, S.C. Nutritional metabolomics and breast cancer risk in a prospective study. Am. J. Clin. Nutr. 2017, 106, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Schabath, M.B.; Cote, M.L. Cancer Progress and Priorities: Lung Cancer. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Knekt, P.; Aromaa, A.; Maatela, J.; Aaran, R.K.; Nikkari, T.; Hakama, M.; Hakulinen, T.; Peto, R.; Teppo, L. Vitamin E and cancer prevention. Am. J. Clin. Nutr. 1991, 53 (Suppl. S1), 283S–286S. [Google Scholar] [CrossRef] [PubMed]
- Alpha-Tocopherol, Beta Carotene Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Weinstein, S.J.; Yu, K.; Männistö, S.; Albanes, D. A Prospective Study of Serum Vitamin E and 28-Year Risk of Lung Cancer. J. Natl. Cancer Inst. 2020, 112, 191–199. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, Y.; Shu, Y.; Chen, X.; Hu, J.; Zheng, R.; Ma, D.; Yang, C.; Guan, X. Link between risk of colorectal cancer and serum vitamin E levels: A meta-analysis of case-control studies. Medicine 2017, 96, e7470. [Google Scholar] [CrossRef] [PubMed]
- Chapelle, N.; Martel, M.; Toes-Zoutendijk, E.; Barkun, A.N.; Bardou, M. Recent advances in clinical practice: Colorectal cancer chemoprevention in the average-risk population. Gut 2020, 69, 2244–2255. [Google Scholar] [CrossRef]
- Yang, C.S.; Luo, P.; Zeng, Z.; Wang, H.; Malafa, M.; Suh, N. Vitamin E and cancer prevention: Studies with different forms of tocopherols and tocotrienols. Mol. Carcinog. 2020, 59, 365–389. [Google Scholar] [CrossRef]
- Luo, H.; Fang, Y.J.; Lu, M.S.; Pan, Z.Z.; Huang, J.; Chen, Y.M.; Zhang, C.X. Dietary and serum vitamins A and E and colorectal cancer risk in Chinese population: A case-control study. Eur. J. Cancer Prev. 2019, 28, 268–277. [Google Scholar] [CrossRef]
- World Cancer Research Fund International Prostate Cancer Statistics. Available online: https://www.wcrf.org/cancer-trends/prostate-cancer-statistics/ (accessed on 22 August 2022).
- Cui, R.; Liu, Z.Q.; Xu, Q. Blood α-tocopherol, γ-tocopherol levels and risk of prostate cancer: A meta-analysis of prospective studies. PLoS ONE 2014, 9, e93044. [Google Scholar] [CrossRef]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2009, 301, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Traber, M.G. Mechanisms for the prevention of vitamin E excess. J. Lipid Res. 2013, 54, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Diab, L.; Krebs, N.F. Vitamin Excess and Deficiency. Pediatr. Rev. 2018, 39, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US) Panel on Dietary Antioxidants and Related Compounds. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press: Washington, DC, USA, 2000. [Google Scholar]
- Owen, K.N.; Dewald, O. Vitamin E Toxicity. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Freedman, J.E.; Keaney, J.F. Vitamin E inhibition of platelet aggregation is independent of antioxidant activity. J. Nutr. 2001, 131, 374S–377S. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.R.; Pastor-Barriuso, R.; Dalal, D.; Riemersma, R.A.; Appel, L.J.; Guallar, E. Meta-analysis: High-dosage vitamin E supplementation may increase all-cause mortality. Ann. Intern. Med. 2005, 142, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: Systematic review and meta-analysis. JAMA 2007, 297, 842–857. [Google Scholar] [CrossRef]
- Klein, E.A.; Thompson, I.M.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the risk of prostate cancer: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef] [Green Version]
- Lonn, E.; Bosch, J.; Yusuf, S.; Sheridan, P.; Pogue, J.; Arnold, J.M.; Ross, C.; Arnold, A.; Sleight, P.; Probstfield, J.; et al. Effects of long-term vitamin E supplementation on cardiovascular events and cancer: A randomized controlled trial. JAMA 2005, 293, 1338–1347. [Google Scholar] [CrossRef] [PubMed]



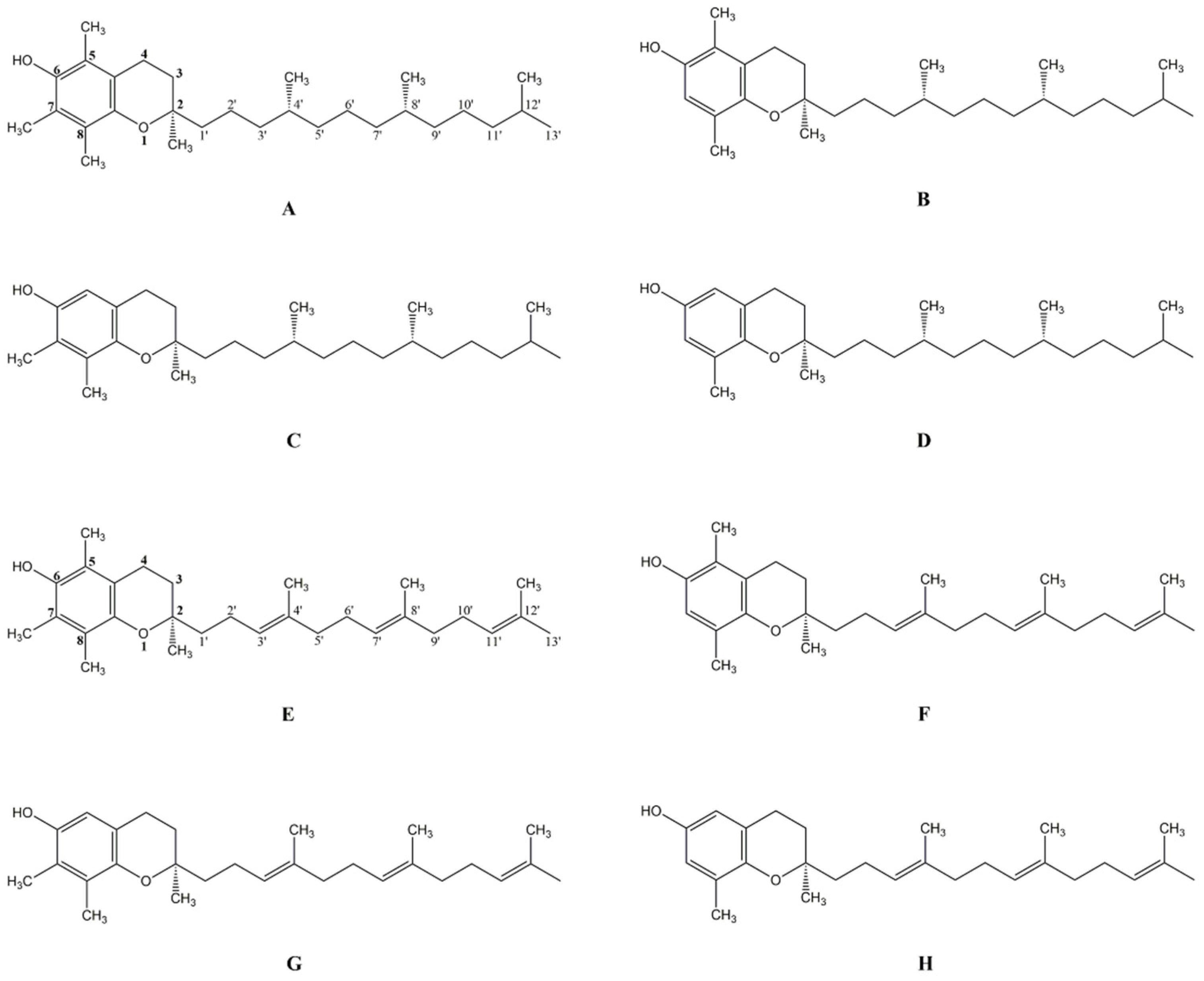{kind=link}
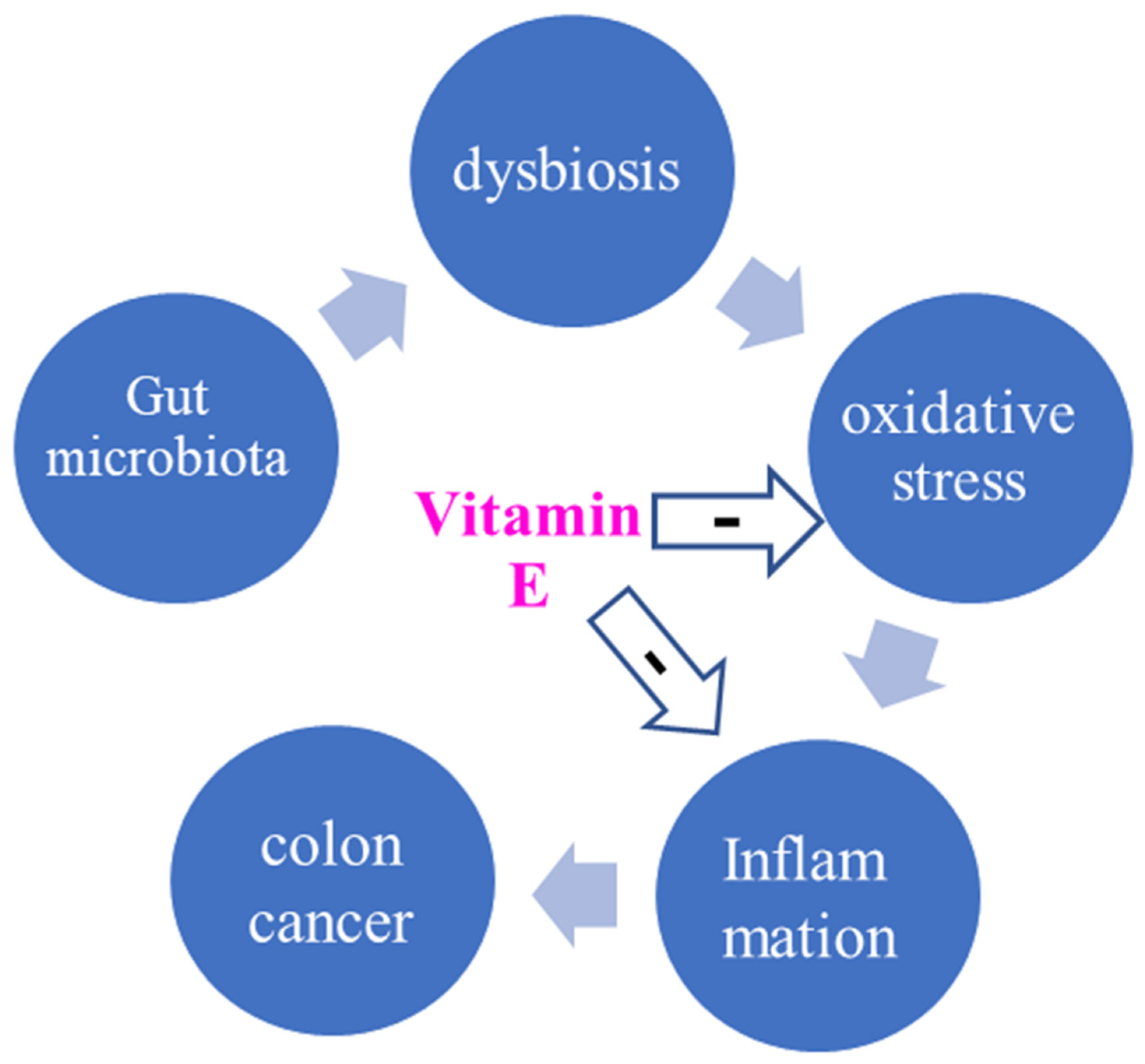{kind=link}
| Pathological Conditions | Reported Effect | References |
|---|---|---|
| Oxidative stress | Antioxidant by scavenging free radicals, especially peroxyl radicals, and singlet oxygen. | [2] |
| To maintain the integrity of long-chain polyunsaturated fatty acids in the membranes of cells and thus maintain their bioactivity. Maintaining the membranes integrity by inhibiting long-chain polyunsaturated fatty acids oxidation | [10] | |
| Preserving mitochondrial efficiency | [4,6,15] | |
| Inflammation | Inhibition of inflammation mediators and related enzymes | [3,4,18,19] |
| Dysbiosis | Modulatory effect on gut microbiota | [30,31] |
| CVD | Reduction in atherosclerotic plaque formation, CVD outcomes, myocardial infraction, and ischemic stroke risks | [12,36] |
| Lowering of CD36 expression | [3,12,37] | |
| Elevation of blood pressure by αT or mixed tocopherols | [4] | |
| Reduction in platelets coagulation and aggregation with αT or γT supplementation | [4] | |
| Reduction in atherosclerosis progression with vitamin E and coenzyme Q supplementation | [4] | |
| Reduction in oxidized HODE in the hearts of α-TOH-treated mice | [36] | |
| αT administration causes anti-oxidative response following I/R damage | [36] | |
| Protective effect of αT on mitochondrial integrity | [12,36] | |
| Inhibition of COXs and 5-LOX by 13′-COOHs and αT | [3,4,7] | |
| Reduction in lipids oxidation in the myocardium by αT | [36,37] | |
| Reduction in CVD risk factors with vitamin E, C and L-arginine | [12] | |
| Reduction in ischemic heart disease risk using vitamin E, C, and/or vitamin A | [12] | |
| Reduction in apoptotic activity with vitamin E | [12] | |
| Diabetes | No relationship with αT serum levels | [43] |
| Lower αT plasma levels in subjects with T2DM | [44] | |
| No protective effect of vitamin E supplementation | [43,44,45] | |
| Protective effect against diabetes-induced complications | [46] | |
| Protective effect against diabetes-induced atherosclerosis | [49,50] | |
| Conflicting result about the protective effect against diabetes-induced atherosclerosis | [51] | |
| Protective effect against diabetes-induced nephropathy | [53,54] | |
| Protective effect against diabetes-induced retinopathy | [55,56,57,58] | |
| Lower vitamin E serum levels in people with diabetes-induced retinopathy | [56] | |
| Asthma | Protective effect of vitamin E maternal intake | [19,61,62] |
| Protective effect of vitamin E | [19,63,64] | |
| Breast cancer | Possible protective effect of vitamin E | [67] |
| Inverse association with breast cancer risk only for vitamin E dietary intake in postmenopausal women | [68] | |
| Inverse relationship between αT serum levels and ER+ breast cancer risk, but positive relationship with serum ẟT and conjugated γT. | [69] | |
| Lung cancer | No relationship with αT serum levels | [71] |
| No protective effect of vitamin E supplementation | [72] | |
| Inverse relationship between αT serum basal levels and lung cancer risk | [73] | |
| No protective effect of vitamin E supplementation | [73] | |
| Colorectal cancer | Lower vitamin E plasma levels in subjects with colorectal cancer (for Caucasian population) | [74] |
| No protective effect of vitamin E | [75,76] | |
| Inverse relationship between vitamin E intake and colorectal cancer risk, but no relationship with αT serum levels | [77] | |
| Prostate Cancer | Inverse relationship between αT serum levels and prostate risk, no relationship with γT | [79] |
| No protective effect of αT supplementation | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciarcià, G.; Bianchi, S.; Tomasello, B.; Acquaviva, R.; Malfa, G.A.; Naletova, I.; La Mantia, A.; Di Giacomo, C. Vitamin E and Non-Communicable Diseases: A Review. Biomedicines 2022, 10, 2473. https://doi.org/10.3390/biomedicines10102473
Ciarcià G, Bianchi S, Tomasello B, Acquaviva R, Malfa GA, Naletova I, La Mantia A, Di Giacomo C. Vitamin E and Non-Communicable Diseases: A Review. Biomedicines. 2022; 10(10):2473. https://doi.org/10.3390/biomedicines10102473
Chicago/Turabian StyleCiarcià, Giulia, Simone Bianchi, Barbara Tomasello, Rosaria Acquaviva, Giuseppe Antonio Malfa, Irina Naletova, Alfonsina La Mantia, and Claudia Di Giacomo. 2022. "Vitamin E and Non-Communicable Diseases: A Review" Biomedicines 10, no. 10: 2473. https://doi.org/10.3390/biomedicines10102473