
The First Thousand Days: Kidney Health and Beyond


1
Department of Pharmacy, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan
2
School of Pharmacy, Kaohsiung Medical University, Kaohsiung 807, Taiwan
3
Department of Pediatrics, Kaohsiung Chang Gung Memorial Hospital and Chang Gung University College of Medicine, Kaohsiung 833, Taiwan
4
Institute for Translational Research in Biomedicine, Kaohsiung Chang Gung Memorial Hospital and Chang Gung University College of Medicine, Kaohsiung 833, Taiwan
*
Author to whom correspondence should be addressed.
Healthcare 2021, 9(10), 1332; https://doi.org/10.3390/healthcare9101332
Submission received: 22 August 2021
/
Revised: 25 September 2021
/
Accepted: 3 October 2021
/
Published: 6 October 2021
(This article belongs to the Special Issue The First 1000 Days of Infant)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The global burden of chronic kidney disease (CKD) is rising. A superior strategy to advance global kidney health is required to prevent and treat CKD early. Kidney development can be impacted during the first 1000 days of life by numerous factors, including malnutrition, maternal illness, exposure to chemicals, substance abuse, medication use, infection, and exogenous stress. In the current review, we summarize environmental risk factors reported thus far in clinical and experimental studies relating to the programming of kidney disease, and systematize the knowledge on common mechanisms underlying renal programming. The aim of this review is to discuss the primary and secondary prevention actions for enhancing kidney health from pregnancy to age 2. The final task is to address the potential interventions to target renal programming through updating animal studies. Together, we can enhance the future of global kidney health in the first 1000 days of life.
1. Introduction
The first 1000 days of life, the period from conception to two years of age, is crucial for the individual’s later development. Our body can adapt in response to stimuli from the environment through alterations of structure or function, namely, developmental plasticity [1]. Suboptimal environmental conditions in this unique period can cause adverse long-term health outcomes. This theory has now evolved into the Developmental Origin of Health and Disease (DOHaD) theory [2]. These environmental factors include, but are not limited to, nutrition, maternal illness, environmental chemicals, substance abuse, medication use, infection, and exogenous stress [3]. Recent advances in epidemiological and experimental studies have offered considerable insight into how various environmental influences during early development increase the risk for developing chronic, especially non-communicable, disease (NCD), in later life [4].
Chronic kidney disease (CKD) is one of the most prevalent NCDs [5]. More importantly, CKD is a key determinant of poor health outcomes for major NCD [6]. The global burden of CKD is rising and now affects 10% of the world’s adult population [7]. CKD can originate in early life. A variety of adverse environmental conditions are associated with the programming of kidney disease [8,9,10]. We now know that programming processes before disease becomes apparent are modifiable by shifting the therapeutic approach from adulthood to early life, namely, reprogramming [11,12]. Accordingly, this vision proposes that greater attention is needed on global kidney health policy, particularly focus on the prevention of kidney disease in the earliest stage, not just the treatment of established CKD [13]. Thus, this review places specific emphasis on gaining a greater understanding of the pathophysiological phenomenon behind the programming of kidney disease and current evidence relating to preventing CKD in the first 1000 days of life by developing a potential reprogramming strategy. Our search strategy was designed to retrieve literature relating to DOHaD and kidney disease from PubMed/MEDLINE databases. Specific emphasis was placed on environmental insult exposure during pregnancy, lactation, and infant stages. Additional studies targeting the pathogenesis of developmental programming of kidney disease were also considered.
2. Biological Processes Shaping Kidney Development
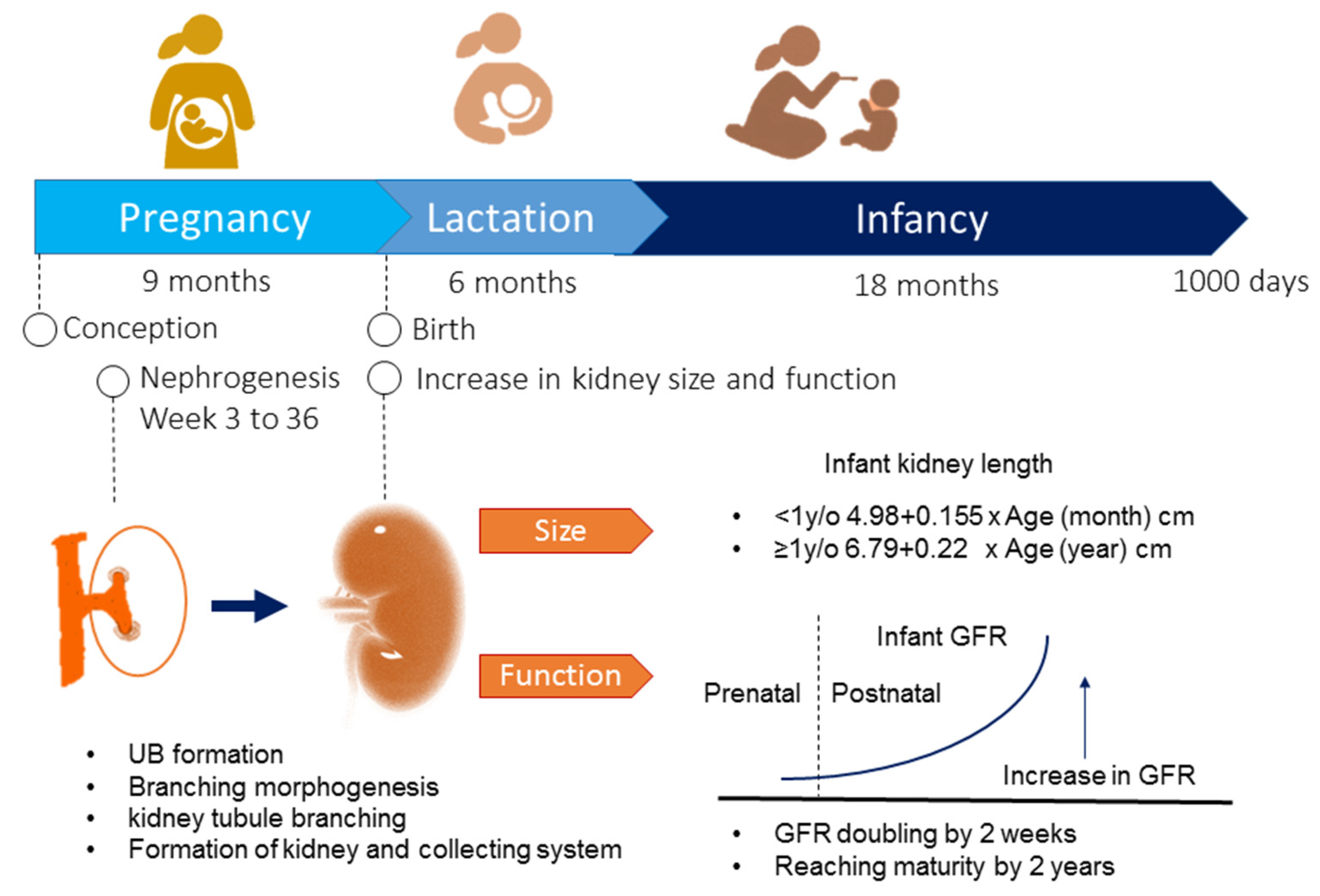
Figure 1 illustrates the biological processes of kidney development during the first 1000 days of life. In humans, kidney development begins at week three and ends at around 36 weeks of gestation [14]. The metanephric kidney is initiated when the ureteric bud (UB) forms and elongates to invade the adjacent metanephric mesenchyme (MM) [15]. The MM forms nephrons, while the UB tip branches serially to form the collecting duct. The renal vesicles form by a mesenchyme to epithelium conversion and are the precursors of the nephrons. Branching morphogenesis establishes an extensive ureteric bud arborization [15], which eventually differentiates into the collecting duct system and leads to the formation of the nephrons. A nephron is the basic functional unit of the kidney. The human kidneys are composed of approximately 1 million nephrons, with a 10-fold interindividual variability [14]. An exponential increase in nephrons occurs between 18 and 32 weeks of gestation. By the end of gestation, nephron development is complete [16]. In general, nephrogenesis is complete by term birth.
After birth, the kidney continues to grow in size. For infants younger than 1 year, the equation is as follows: renal length (cm) = 4.98 + 0.155 X age (months) [17]. For babies older than 1 year, the regression equation is as follows: renal length (cm) = 6.79 + 0.22 X age (years) [17]. Regarding renal function, the glomerular filtration rate (GFR) doubles in the first 2 weeks of life from a value of 20 mL/min/1.73 m2 at birth in full-term neonates. The GFR continues to increase after birth and reaches adult values by two years of age [18].
3. Risk Factors Influencing Kidney Health and Development
Branching morphogenesis is critical for a normal nephron number [15]. Impaired branching morphogenesis and nephrogenesis could cause a reduced nephron number and a broad spectrum of malformed kidneys, namely, congenital anomalies of the kidney and urinary tract (CAKUT) [19]. Developing kidneys are vulnerable to environmental risk factors that impair development during pregnancy: a severe renal maldevelopment occurs during early pregnancy, while kidney defects that occur later are generally less severe [19]. A case-control study recruiting more than 1.6 million infants demonstrated that risk factors for CAKUT consist of prematurity, low birth weight (LBW), male sex, maternal gestational diabetes, maternal thalassemia, oligohydramnios or polyhydramnios, and first parity [20].
Human and experimental studies suggest genetic factors, including chromosomal anomalies, copy number variants, and monogenic mutations/deletions, likely contribute to approximately 45% of CAKUT. Environmental factors and not yet identified genetic factors contribute to the remainder [19,21,22]. Although hundreds of candidate genes have been identified, CAKUT cannot be attributed to a monogenic cause in more than 80% of cases [19]. Considering the phenotypes from genetic defects vary considerably, this suggests that many cases of CAKUT are polygenic. Notably, gene–gene and gene–environment interactions have also contributed to CAKUT [23]. One example is apolipoprotein L1 (APOL1) variants [24,25]. Prior research suggests that environmental stressors and APOL1 may contribute to the CKD phenotype variance associated with APOL1 risk alleles. Under basal conditions, genotypes carrying risk alleles appear to have a subtle phenotype that is not disease causing. However, high-risk genotypes cannot adapt to the stresses and lose renal function, resulting in CKD throughout the entire lifespan [25]. Additionally, genes are vulnerable to epigenetic modification in response to adverse conditions during the first 1000 days of life. Likewise, epigenetic changes may provide a mechanistic link whereby early life exposures lead to long-term increased risk of kidney disease in adulthood.
As nephrogenesis is completed by full-term birth, premature infants tend to develop low nephron endowment. Additionally, a low nephron number is related to compromised pregnancy, low birth weight, intrauterine growth retardation, inadequacy of postnatal nutrition, and treatment with nephrotoxic drugs after birth, etc. [9,16,26]. The role of low nephron number in renal programming is gaining attention as it can cause glomerular hyperfiltration and compensatory glomerular hypertrophy, and can initiate a vicious cycle of further reductions in nephrons [16]. As kidney disease is possibly the result of interactions among multiple hits [27], a low nephron number may create a first hit to the kidney, which increases the vulnerability of remaining nephrons to develop CKD when facing other environmental insults as a second hit in later life.
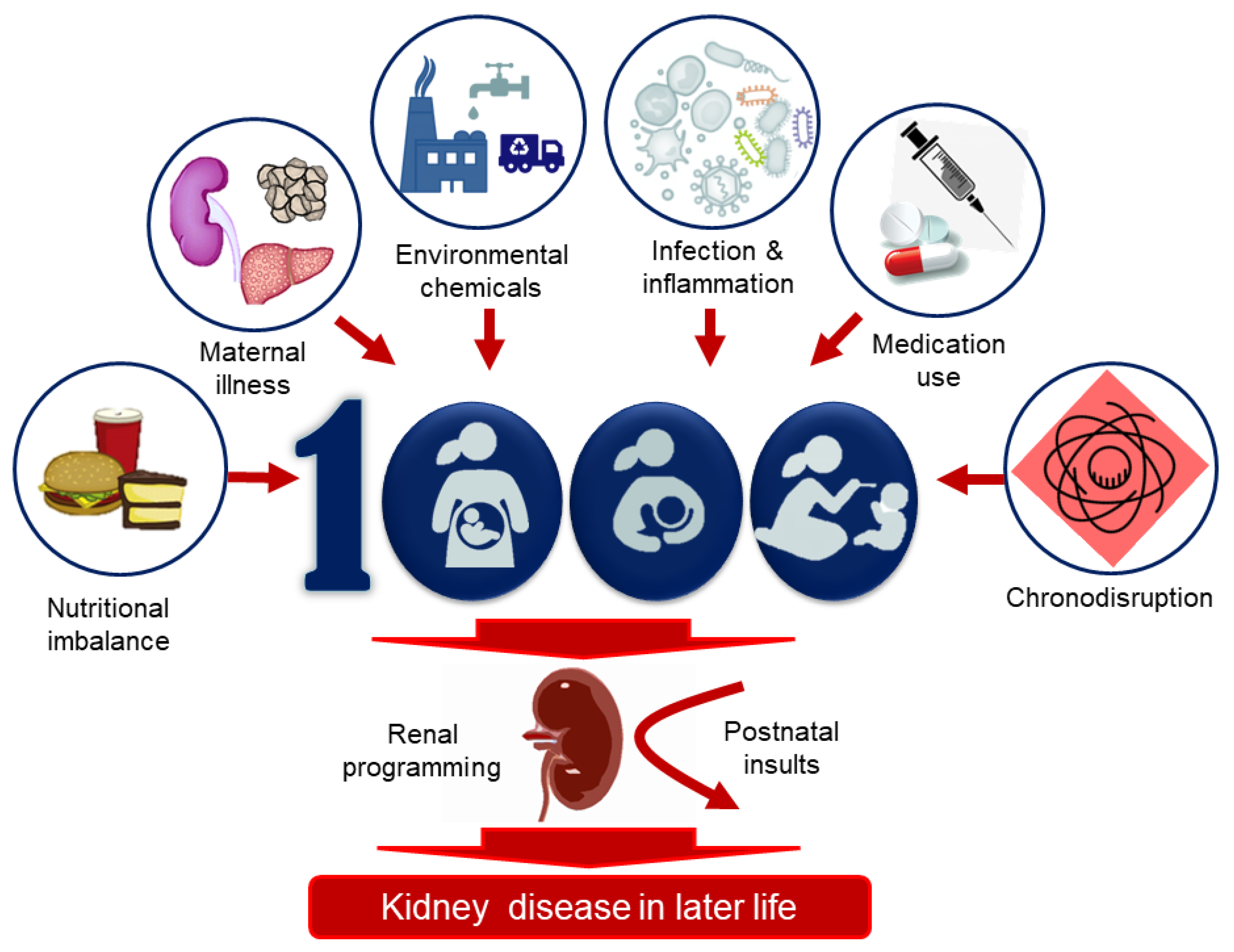
Thus far, several environmental risk factors have been linked to the programming of kidney disease, including nutritional imbalance, maternal illness, environmental chemicals, substance abuse, medication use, infection, and exogenous stress (Figure 2). These are discussed in turn.
3.1. Nutritional Imbalance
The insufficient or excessive consumption of certain nutrients has been linked to the developmental programming of kidney disease [28,29,30]. Important support was first provided by the Dutch famine birth cohort study for renal programming, which demonstrated that undernutrition during pregnancy has long-lasting consequences for adult health, including kidney disease [31]. Several epidemiological studies have associated maternal nutrition during pregnancy with offspring kidney structure and function in humans, as reviewed elsewhere [29]. Deficiencies in maternal folate [32], vitamin A [33], and total energy [31] during pregnancy were associated with negative impacts on kidney structure and function, measured by kidney volume, proteinuria, and renal function in the offspring [29].
There is convincing evidence from animal studies of nutritional imbalance during pregnancy and lactation affecting renal programming, resulting in kidney disease in adult offspring. Various nutritional factors have been related to renal programming, including calorie restriction [34], protein restriction [35], low-salt intake [36], magnesium-deficient diet [37], high sucrose consumption [38], high-fructose diet [39], high-fat diet [40], and high-salt diet [41].
Additionally, nutritional imbalance during pregnancy and lactation is also associated with a reduced nephron number, a key determinant of adulthood kidney disease. In rat models, nutritional insults last only for a brief period, as little as 1–2 days, but can impair kidney development, resulting a permanent low nephron endowment [9]. Currently, a variety of animal models of early-life suboptimal nutrition, such as maternal caloric restriction [42], low protein diet [43], vitamin A deficiency [44], multi-deficient diet [45], iron restriction diet [46], low-salt diet [47], and high-salt diet [47], have been reported to impair nephrogenesis, resulting in a reduced nephron number.
3.2. Maternal Illness
Maternal illnesses and complications during pregnancy can drive renal programming and increase the risk for developing kidney disease later in life. Several animal models resembling human illnesses and pregnancy complications have been established to study renal programming-related offspring outcomes, such as hypertensive disorders of pregnancy [48], preeclampsia [49], CKD [50], diabetes [51], and sleep disorder [52]. Hypertension affects up to 10% of pregnancies [53]. In spontaneously hypertensive rats, maternal hypertension is related to renal programming and hypertension in the adult offspring [48]. Another study showed pregnant rats treated with NG-nitro-L-arginine-methyl ester (L-NAME, a nitric oxide synthase inhibitor) to mimic maternal preeclampsia caused elevated blood pressure (BP) and renal programming in their adult offspring [49]. To study the influence of maternal CKD on offspring’s renal outcome, we used an adenine-induced maternal CKD model. Our findings indicated that uremia-related adverse outcomes in adult offspring included renal hypertrophy and hypertension [54].
Diabetes in pregnancy is also thought to impair nephrogenesis. Human studies have demonstrated that adults born to mothers with gestational diabetes have an increased risk of CAKUT [20,55] and kidney disease [56]. Another observational study showed that diabetes during pregnancy influences fetal kidney growth, indicating a negative effect on nephrogenesis [56]. In a streptozotocin-induced diabetes rat model, offspring born to diabetic dams developed a reduced nephron number, renal hypertension, and kidney injury [51]. Moreover, sleep disorder in pregnancy also affects kidney development. In a maternal sleep restriction model, adult offspring displayed an enlarged glomeruli diameter and a reduced number of glomeruli coinciding with hypertension at 2 months of age [52]. These findings suggest that sleep restriction during pregnancy impairs nephrogenesis, resulting in renal programming-related disorders in offspring.
From human and animal studies, maternal obesity is another risk factor for CKD in offspring [57]. An observational study recruiting 3093 CAKUT cases showed a positive association between maternal obesity and CAKUT in offspring [58]. Another meta-analysis study supports the notion that maternal obesity adversely impacts renal programming in offspring, with an increased risk of kidney disease in adulthood [59]. In various animal models of maternal obesity, offspring from obese mothers had higher serum creatinine levels, a 24 h urinary albumin to creatinine ratio, and worse renal tubular injury and glomerulosclerosis scores [40,60,61]. In view of the alarming increase in global obesity, more attention should be paid to studying how maternal obesity influences CKD development in offspring.
3.3. Environmental Chemicals
Numerous environmental chemicals pose a wide range of adverse effects on the kidney [62]. During kidney development, some chemicals can impair nephrogenesis, leading to low nephron endowment and CAKUT [63]. Accordingly, developmental nephrotoxic effects can be expected during the environmental chemical exposure of pregnant women. After birth, infants can still be at an increased risk of nephrotoxicity to elemental (e.g., mercury) or organic contaminants (e.g., melamine) [64,65].
There are several observational studies addressing the implication of maternal environmental chemical exposure in offspring’s renal outcome. Two studies investigated the associations between maternal lead levels and renal outcomes in offspring [66,67]. One study found there were no associations between maternal lead levels and estimated GFR (eGFR) at 8–12 years of age. However, they observed that the maternal lead level was negatively associated with kidney volume in children [66]. Another study reported there was an inverse association between maternal blood lead levels and eGFR in overweight children at 8–12 years of age [67]. Epidemiological studies revealed that maternal exposure to polycyclic aromatic hydrocarbon, per- and polyfluoroalkyl substances, phthalates, polycyclic aromatic hydrocarbon, and PM2.5/PM10 associated with preterm birth and LBW [68,69,70,71,72], are both risk factors related to a low nephron number.
Evidence from experimental studies also support that exposure during pregnancy can affect kidney development, resulting in renal programming. Maternal exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) or bisphenol A causes a rise in BP in adult rat offspring [73,74], which is relevant to renal programming. Additionally, hydronephrosis was described in rat offspring prenatally exposed to TCDD [75]. Animal studies of the implication of maternal heavy metal exposure in the kidneys of offspring suggested that cadmium is the main cause of adverse renal outcomes [76].
3.4. Substance Abuse
As with nutrient and chemical effects during kidney development, substance abuse is also a major maternal insult. In the United States, roughly 6–16% of pregnant women are cigarette smokers, alcohol abusers, or illicit drug users [77]. An observational study has shown that maternal alcohol exposure has a dose-dependent adverse effect on renal function in overweight and obese children [78]. Another cohort study revealed that maternal alcohol exposure is associated with the development of mild CKD in their offspring at 30 years [79]. Similarly, in a maternal ethanol exposure rat model, reduced nephron number and renal function were reported in adult offspring, possibly as a result of inhibited ureteric branching morphogenesis [80].
In humans, maternal smoking during pregnancy is associated with fetal and infant kidney volume [81]. Likewise, prior research on animal models demonstrated that maternal nicotine exposure adversely affected fetal kidney development, resulting in CKD in offspring [82,83,84]. Though illicit drug use is associated with a higher risk of CKD progression [85], whether maternal illicit drug use affects offspring’s renal outcomes remains largely unknown.
3.5. Infection and Inflammation
Intrauterine infection is a crucial and frequent mechanism leading to preterm birth [86]. Microbial endotoxins and proinflammatory cytokines stimulate the production of prostaglandins, resulting in uterine contractility [87]. Furthermore, emerging evidence indicates that maternal infections contribute to poor birth outcomes, such as LBW and preterm birth, by the inflammation-mediated disruption of placental development and function [88].
The influence of infection during gestation on offspring’s renal outcomes has been studied in animal models. Maternal exposure to lipopolysaccharide (LPS) caused offspring hypertension coinciding with renal programming [89,90]. Another study revealed that prenatal LPS exposure augmented neonatal hyperoxia-induced kidney injury [91].
After birth, urinary tract infection (UTI) is one of the major infections contributing to adverse renal outcomes [92]. Pyelonephritis can lead to renal scarring and result in hypertension and even kidney failure. Approximately 30 % of children who develop a UTI are subsequently diagnosed with vesicoureteral reflux [93]. Reflux nephropathy is reported as the fourth most frequent cause of end-stage kidney disease in the pediatric population [92].
3.6. Medication Use
The existing literature suggests that a number of drugs administrated to pregnant women may affect kidney development, leading to CAKUT [84]. These medications include, but are not limited to, aminoglycosides, cyclosporine A, NSAIDs, ACE inhibitor (ACEI)/angiotensin receptor blockers (ARBs), dexamethasone, furosemide, anti-epileptic drugs, Adriamycin, and cyclophosphamide [94]. In various animal models, cyclosporine A [95], gentamicin [96], and glucocorticoid [97,98,99] have been associated with a low nephron number and renal programming [9].
Most nephrotoxic drugs in mature kidneys may also have toxic effects on developing kidneys. It is worth noting that drugs that are not nephrotoxic in fully developed kidneys may impair the balance of growth factors that are crucial for kidney development. For example, ACEI/ARBs are well known to exert renoprotective benefits [100]. However, these drugs have been avoided in pregnant women due to ACEI/ARB fetopathy and renal maldevelopment [101]. The reason for this is that the suppression of the intrarenal RAS contributes to altered structural development of the kidney [102]. Another example is glucocorticoid. Currently, antenatal glucocorticoid administration is recommended in women at risk of preterm birth to accelerate fetal lung maturation [103]. In normal pregnancy, the fetus is protected by the placental inactivation of active glucocorticoids [104]. Accordingly, excessive glucocorticoid through exogenous administration has been related to renal programming, resulting in a low nephron number [104]. In addition to exogenous administration, a developing fetus is likely to be exposed to excessive glucocorticoids of maternal origin (e.g., due to a stressed pregnancy). In rats, repeated dexamethasone administration on embryonic days 15 and 16 [97], from gestational days 16 to 22 [98], or from postnatal days 1 to 3 [99] was associated with reduced nephron numbers and resulted in hypertension in adult rats’ offspring.
3.7. Chronodisruption
Human studies have shown a link between gestational chronodisruption and adverse pregnancy outcomes [105,106]. In pregnant women, the disruption of circadian rhythms can occur through shift work, jet travel across time zones, or exposure to light at night [107]. A meta-analysis study recruiting 196,989 women reported that working rotating shifts is associated with preterm birth and small for gestational age (SGA), both risk factors for a low nephron number [106]. In rats, chronic photophase shifts throughout pregnancy program adult offspring to display renal dysfunction and hypertension [108].
4. Behind the First 1000 Days of Life
As summarized above, a diversity of environmental risk factors is associated with the programming of kidney disease during the first 1000 days of life. Based on what is now known about the magnitude of kidney development in the first 1000 days of life, it is not surprising that more work is needed to understand the underlying mechanisms behind the pathophysiology of kidney disease programming. A better understanding of these mechanisms will help in targeted therapy and prevention.
4.1. Mechanisms of Later Kidney Disease of Developmental Origin
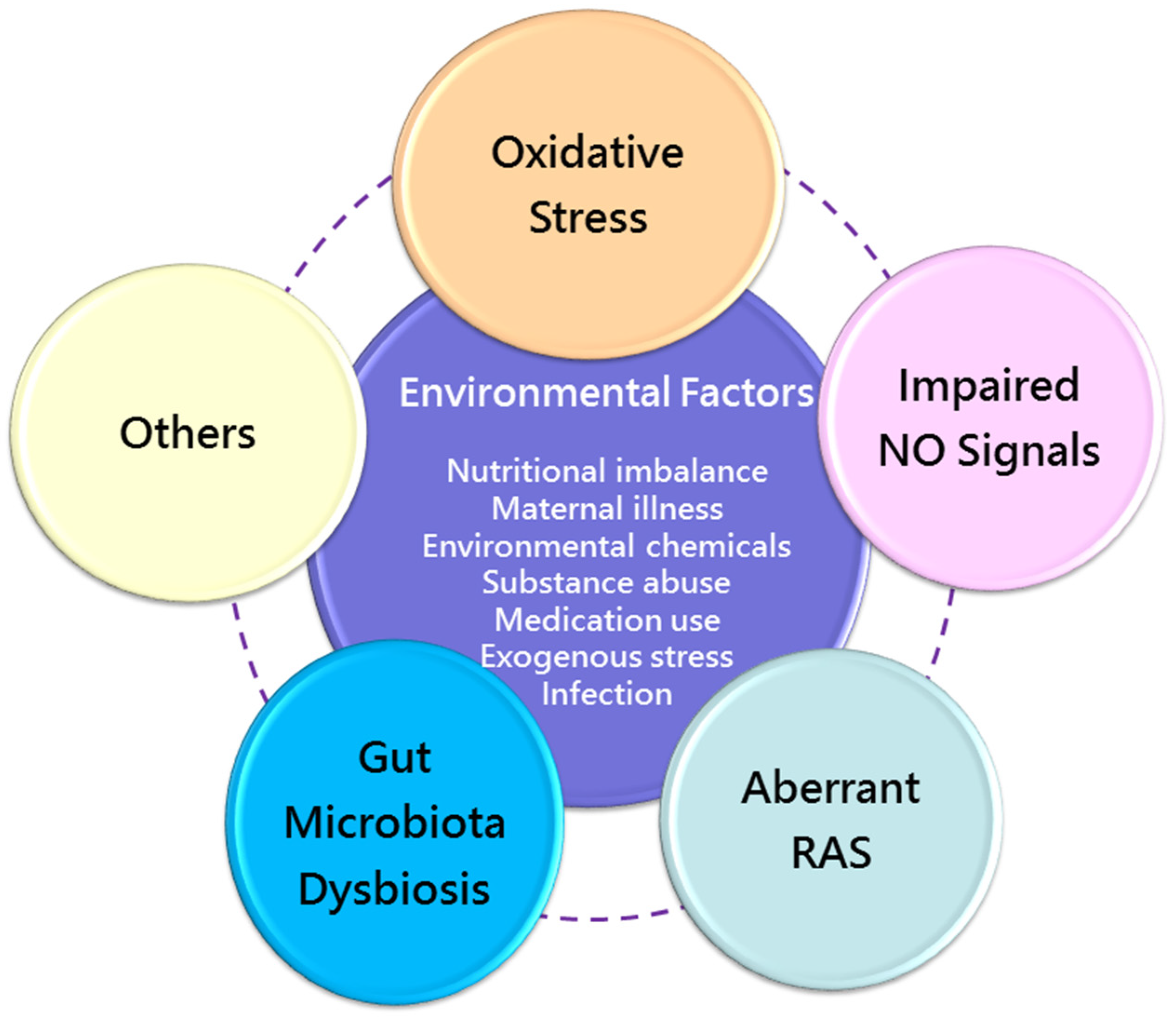
Despite various early-life environmental factors related to CKD in later life, current evidence suggests that there may be common mechanisms behind renal programming. Although the complete mechanisms remain inconclusive, prior research has provided important information on certain molecular mechanisms, including oxidative stress [109], nitric oxide (NO) signaling [110], aberrant renin–angiotensin system (RAS) [111], and gut microbiota dysbiosis [112]. A summary of the integrated mechanisms of renal programming in response to various maternal insults for kidney disease of developmental origin is depicted in Figure 3.
Oxidative stress is considered to play a critical role in fetal programming [113]. Several mechanisms of oxidative stress have been related to renal programming, including the increased production of reactive oxygen species (ROS), antioxidant system dysfunction, and increased oxidative damage. As reviewed elsewhere, a number of animal models demonstrate oxidative stress involved in renal programming [105]. Nutritional imbalance during pregnancy and lactation is the most common factor to induce the programming of kidney disease. For example, calorie restriction [42] and increased consumption of a high-fat diet [40], fructose [114], or methyl donors [115] have been addressed previously. Other environmental factor associated with renal programming, such as environmental chemicals [73], substance abuse [82], maternal illness [50], inflammation [91], and medication use [97,98], have all been linked to oxidative stress.
A reduced nephron number induced by oxidative stress has been reported in the caloric restriction model [42], streptozotocin-induced diabetes [51], and maternal smoking [116]. As we reviewed elsewhere [94], renal programming induced by a variety of maternal insults is associated with increased F2-isoprostanes [49], malondialdehyde (markers of lipid peroxidation) [117], and 8-hydroxydeoxyguanosine (8-OHdG, an oxidative DNA damage marker) [49]. Conversely, the perinatal use of antioxidants has shown benefits against oxidative stress-induced renal programming in various animal models [109].
Renal programming, in addition to from oxidative stress, has been associated with impaired NO signals [110]. NO, a potent vasodilator, plays a key role in fetal development during pregnancy [118]. Nitric oxide synthase (NOS) catalyzes L-arginine to generate NO. However, in certain conditions, such as inhibition by NOS inhibitor asymmetric dimethylarginine (ADMA) [119], uncoupled NOS produces superoxide, consequently resulting in peroxynitrite formation. Accordingly, reduced NO bioavailability as a result of NOS uncoupling has been linked to kidney disease of developmental origin [110]. Moreover, our prior research showed that nephrogenesis was inhibited by ADMA, a ROS inducer, as well as an endogenous NOS inhibitor, consequently leading to a reduction in the nephron number [51].
Numerous interventions targeting the NO pathway in pregnancy to protect offspring against kidney disease have been reported [110]. These interventions include supplementation with substrates for NOS, NO donors, ADMA-lowering agents, and the enhancement expression/activity of NOS enzymes.
Similar to oxidative stress and NO, aberrant RAS appears to be involved in the pathogenesis of renal programming [115]. In the developing kidney, RAS genes are highly expressed and essential for mediating the proper formation of the renal structure and function [120]. Mutations in RAS genes are associated with kidney malformation in humans [121], which is in agreement with studies positing that the RAS is directly blocked by ACEI/ARBs [101]. Likewise, genetic inactivation of the angiotensinogen, renin, angiotensin converting enzyme (ACE), angiotensin II type 1 (AT1R) or type 2 receptor in mice leads to a broad phenotypic spectrum of CAKUT [121].
Angiotensin II (Ang II), the major player in the RAS, can mediate several key events of the inflammatory processes via AT1R stimulation [122,123]. These processes include triggering endothelial dysfunction, stimulating the release of cytokine/chemokines, activating NAD(P)H oxidase to produce ROS, and promoting pro-fibrotic growth factors, all contributing to kidney damage.
As reviewed elsewhere [111], most environmental influences that can program the kidney, resulting in adulthood kidney disease, such as nutritional imbalance, maternal illness, substance abuse, environmental chemical exposure, and medication use, are associated with the aberrant activation of the RAS. On the other hand, early blockade of the classical RAS axis appears to reprogram the inappropriately activated RAS to prevent the programming of kidney disease in various animal models.
Gut microbiota and their derived metabolites can affect the function of various target organs through circulation, including the renal systems [124]. Several adverse environmental factors in early life can shape the offspring’s gut microbial composition, leading to consequent adverse offspring outcomes [125]. Conversely, maternal microbiota-targeted interventions have shown benefits against renal programming [112,126]. Importantly, several gut microbiota-derived uremic toxins are associated with cardiovascular disease (CVD) in CKD via the activation of the aryl hydrocarbon receptor (AHR) [127]. Considering that several environmental factors (e.g., nutrition, environmental chemicals, and inflammation) are related to AHR activation, the interplay among gut microbiota, AHR, and the kidney has attracted the attention of researchers for investigating the mechanisms underlying the more thorough programming of kidney disease. Gut microbiota dysbiosis has been linked to hypertension by modulating the gut RAS [128]. On the other hand, ACE2, one component of the RAS, can mediate antimicrobial peptide secretion in the gut, leading to altered gut microbiota composition [129]. These findings indicate that there might be an interconnection between gut microbiota and the RAS behind the pathogenesis of renal programming.
Notably, environmental factors might display other potential mechanisms corresponding to renal programming, such as epigenetic regulation [130], dysregulated nutrient-sensing signals [131], and sex differences [132]. Although there are multiple mechanistic pathways outlined above, they might be interconnected to one another to drive renal programming, resulting in kidney disease. Better understanding the interaction between these common mechanisms and identifying new potential pathways to develop prevention interventions are key in the early prevention of kidney disease.
4.2. Prevention Actions
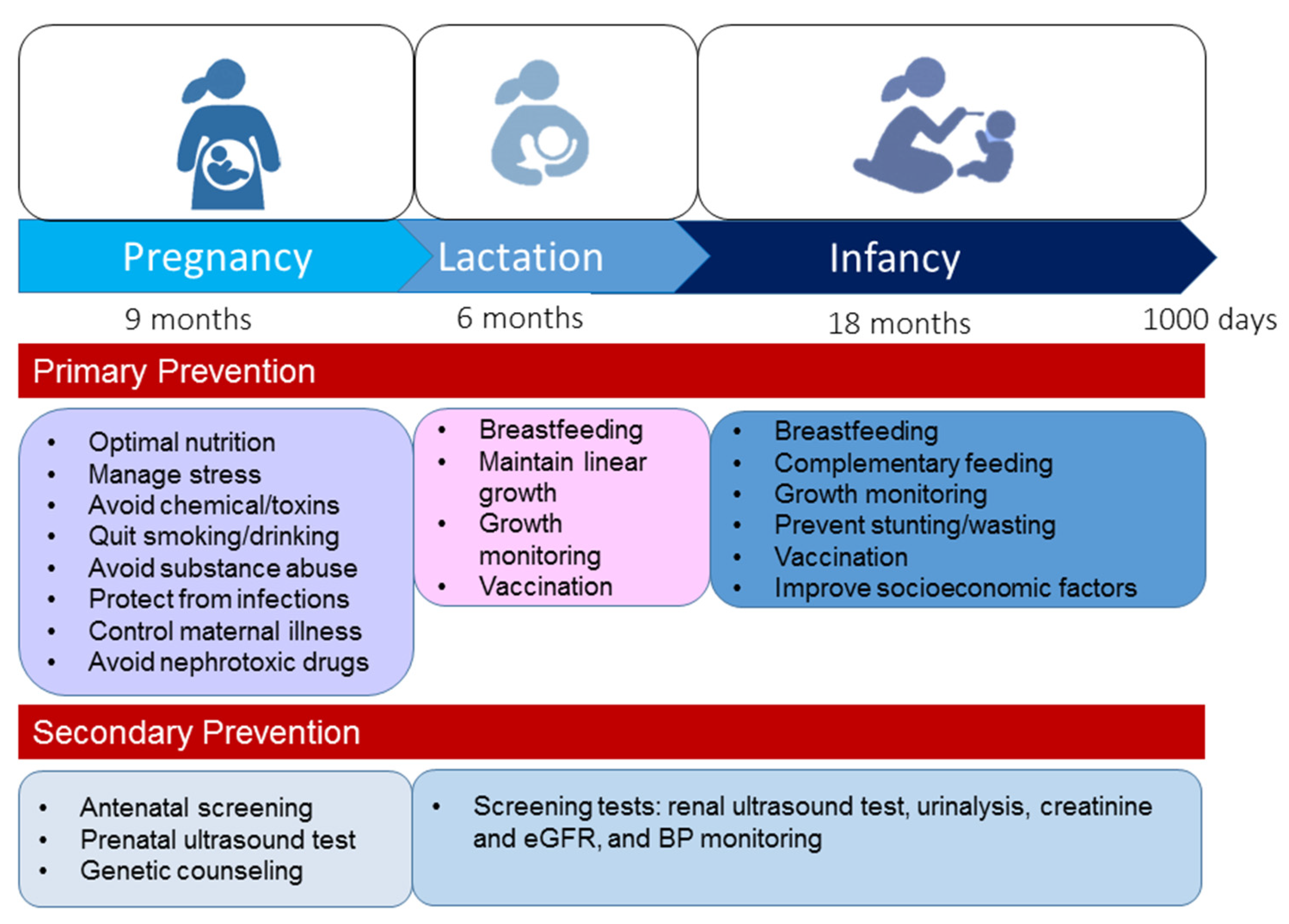
In 2020, World Kidney Day informed the public about the importance of preventive interventions—primary, secondary, or tertiary [13]. In view of the complex nature of CKD, a holistic approach is required to positively impact kidney health. Tertiary prevention aims to manage advanced CKD and related comorbidities, which are rare during the early stage of life. Considering the prevention strategy from a DOHaD perspective, primary and secondary preventions seem to be the best strategy to improve global kidney health during the first 1000 days of life. Figure 4 illustrates the recommended primary and secondary prevention strategies from pregnancy to age 2.
First, primary prevention aims to prevent kidney disease before it occurs. The modifiable risk factors illustrated in Figure 1 should be avoided during the first 1000 days of life. During pregnancy through to early childhood, optimal nutrition is essential for supporting kidney health [133]. Neonates and young infants are particularly vulnerable to infections as they have naïve immune systems. As vaccination is one of the most cost-effective ways of preventing infection, enhanced early life immunity via taking necessary vaccines is essential to protect from infection [134]. Other key points are summarized in Figure 4. Moreover, additional attention is required to improve socioeconomic factors, e.g., access to family planning, equity and education for women, and reduction in poverty [7].
Secondary prevention suggests preventive measures that lead to screening for the early identification of disease and prompt treatment of kidney disease in the earliest stages. Although the early detection of CKD has the potential to yield marked public health benefits, most countries have inadequate CKD detection and surveillance systems to achieve this goal [135]. Some important services are required to screen for and detect kidney disease during the first 1000 days of life, such as antenatal screening, prenatal ultrasound testing, genetic counseling, renal ultrasound testing, urinalysis, creatinine and eGFR, and BP monitoring.
Considering CAKUT have a genetic basis, key nephrogenesis genes may form the basis of genetic screening tests for the future development of novel genetic therapies. In recent years, several potential biomarkers for the early detection of kidney damage have been introduced, and each of these biomarkers has advantages and disadvantages [136,137]. Nevertheless, currently, there is still no ideal biomarker of acute kidney injury. Furthermore, the search for an ideal biomarker predicting the progression of CKD in children with CAKUT is still ongoing. Although neutrophil gelatinase-associated lipocalin, and trefoil family factors (TFF) 1 and 3 have shown the potential to predict CKD progression in children with CAKUT [138,139], they still await more thorough validation. As precursor cell technology has been applied to generate new kidney tissues, more attention will need to be paid to the use of genetically altered metanephric precursor cells to differentiate into functioning kidney tissue for regenerative medicine therapies [140].
Given the advances in the DOHaD research field, it has become clear that kidney disease of developmental origin can be prevented in the earliest stage by reprogramming [11,12]. Prior animal studies have provided essential information in regard to reprogramming strategies. Considering that oxidative stress is a crucial mechanism implicated in renal programming, many natural antioxidants have been used as reprogramming strategies to prevent kidney disease in various animal models [109]: vitamin E and selenium [141], folate [142], L-taurine [143], L-tryptophan [144], N-acetylcysteine [48,49], resveratrol [73,74], and melatonin [44,100]. These findings support the notion that maternal nutrition is a double-edged sword for fetal programming: maternal malnutrition programs many NCDs, whereas nutritional intervention can also be advantageous to prevent adulthood NCDs [145].
Additionally, there are several reprogramming interventions targeting specific signaling pathways giving rise to benefits against renal programming. Targeting of the NO pathway in early life has been employed in various animal models to prevent the development of kidney disease in adult progeny. As reviewed elsewhere [95], these interventions include the supplementation of NO substrate, agents that lower ADMA, NO donors, and the enhancement of NOS expression. Likewise, RAS-based interventions have also shown promising results in protecting against renal programming and related diseases, such as renin inhibitor, ACE inhibitor, ACE-2 (ACE2) activator, and ARBs [111]. Furthermore, reprogramming interventions targeting the hydrogen sulfide (H2S) pathway [146] and nutrient-sensing signals [147] have also shown benefits with regard to kidney disease of developmental origin. Although significant advances have been made from animal research, the need for meaningful clinical translation remains a research priority.
5. Conclusions and Perspectives
Healthy people, living healthy lives on a healthy and peaceful planet were the ultimate goals stated by the United Nations in 2015, to be achieved by 2030 [148]. However, much remains to be accomplished to tackle the challenges of NCDs, in particular, kidney disease [5,6,7]. The concept of the first 1000 days of life allowed us to analyze the literature to determine the causes that could influence kidney development, identify the underling mechanisms of renal programming, and develop potential prevention strategies.
Though various modifiable early-life risk factors have been identified to date, preventive efforts should continue to discover other possible risk factors. Another important aspect is that current preventive strategies mainly focus on promoting a healthy lifestyle and avoiding exposure to risk agents. However, the translation of effective reprogramming interventions from animal studies into clinical practice has been far slower than expected. On all fronts, holistic and multilateral action is essential. Kidney health should be an imperative policy, which can be successfully achieved by the collaboration of doctors, nurses, allied health professionals, researchers, policy makers, and social workers. Only through collaboration can we implement not only patient but also global perspectives toward CKD prevention and the commencement of global kidney health futures in the first 1000 days of life.
Author Contributions
C.-N.H. contributed to the concept generation, the drafting of the manuscript, data interpretation, the critical revision of the manuscript, and the approval of the article; Y.-L.T. contributed to the drafting of the manuscript, data interpretation, the critical revision of the manuscript, concept generation, and the approval of the article. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grant MOST 10-2314-B-182-020-MY3 from the Ministry of Science and Technology, Taiwan, and grants CFRPG8K0021 and CORPG8L0551 from Chang Gung Memorial Hospital, Kaohsiung, Taiwan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gluckman, P.D.; A Hanson, M.; Bateson, P.; Beedle, A.S.; Law, C.M.; A Bhutta, Z.; Anokhin, K.V.; Bougnères, P.; Chandak, G.R.; Dasgupta, P.; et al. Towards a new developmental synthesis: Adaptive developmental plasticity and human disease. Lancet 2009, 373, 1654–1657. [Google Scholar] [CrossRef]
- Haugen, A.C.; Schug, T.T.; Collman, G.; Heindel, J.J. Evolution of DOHaD: The impact of environmental health sciences. J. Dev. Orig. Heal. Dis. 2014, 6, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Barouki, R.; Gluckman, P.D.; Grandjean, P.; Hanson, M.; Heindel, J.J. Developmental origins of non-communicable disease: Implications for research and public health. Environ. Health 2012, 11, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, M.A.; Gluckman, P.D. Early developmental conditioning of later health and disease: Physiology or pathophysiology? Physiol. Rev. 2014, 94, 1027–1076. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.; Tonelli, M.; Bonventre, J.; Coresh, J.; Donner, J.-A.; Fogo, A.B.; Fox, C.S.; Gansevoort, R.T.; Heerspink, H.J.L.; Jardine, M.; et al. Global kidney health 2017 and beyond: A roadmap for closing gaps in care, research, and policy. Lancet 2017, 390, 1888–1917. [Google Scholar] [CrossRef]
- Woo, K.-T.; Choong, H.L.; Wong, K.-S.; Tan, H.B.; Chan, C.-M. The contribution of chronic kidney disease to the global burden of major noncommunicable diseases. Kidney Int. 2012, 81, 1044–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luyckx, V.A.; Tonelli, M.; Stanifer, J.W. The global burden of kidney disease and the sustainable development goals. Bull. World Heal. Organ. 2018, 96, 414–422D. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.A.; Bertram, J.F.; Brenner, B.M.; Fall, C.; E Hoy, W.; E Ozanne, S.; E Vikse, B. Effect of fetal and child health on kidney development and long-term risk of hypertension and kidney disease. Lancet 2013, 382, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsu, C.N. Developmental origins of chronic kidney disease: Should we focus on early life? Int. J. Mol. Sci. 2017, 18, 381. [Google Scholar] [CrossRef] [Green Version]
- Chong, E.; Yosypiv, I.V. Developmental programming of hypertension and kidney disease. Int. J. Nephrol. 2012, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tain, Y.-L.; Joles, J.A. Reprogramming: A preventive strategy in hypertension focusing on the kidney. Int. J. Mol. Sci. 2016, 17, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paauw, N.D.; Van Rijn, B.B.; Lely, A.T.; Joles, J.A. Pregnancy as a critical window for blood pressure regulation in mother and child: Programming and reprogramming. Acta Physiol. 2016, 219, 241–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.K.-T.; Garcia-Garcia, G.; Lui, S.-F.; Andreoli, S.; Fung, W.W.-S.; Hradsky, A.; Kumaraswami, L.; Liakopoulos, V.; Rakhimova, Z.; Saadi, G.; et al. Kidney health for everyone everywhere—From prevention to detection and equitable access to care. Pediatr. Nephrol. 2020, 35, 1801–1810. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.A.; Brenner, B.M. The clinical importance of nephron mass. J. Am. Soc. Nephrol. 2010, 21, 898–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.M.; Sampogna, R.V.; Sakurai, H.; Bush, K.T.; Nigam, S.K. Branching morphogenesis and kidney disease. Development 2004, 131, 1449–1462. [Google Scholar] [CrossRef] [Green Version]
- Bertram, J.F.; Douglas-Denton, R.N.; Diouf, B.; Hughson, M.; Hoy, W. Human nephron number: Implications for health and disease. Pediatr. Nephrol. 2011, 26, 1529–1533. [Google Scholar] [CrossRef]
- Rosenbaum, D.M.; Korngold, E.; Teele, R.L. Sonographic assessment of renal length in normal children. Am. J. Roentgenol. 1984, 142, 467–469. [Google Scholar] [CrossRef]
- Filler, G.; Lopes, L.; Awuku, M. The importance of accurately assessing renal function in the neonate and infant. Adv. Clin. Chem. 2015, 71, 141–156. [Google Scholar] [CrossRef]
- Murugapoopathy, V.; Gupta, I.R. A primer on congenital anomalies of the kidneys and urinary tracts (CAKUT). Clin. J. Am. Soc. Nephrol. 2020, 15, 723–731. [Google Scholar] [CrossRef]
- Tain, Y.L.; Luh, H.; Lin, C.Y.; Hsu, C.N. Incidence and risks of congenital anomalies of kidney and urinary tract in newborns: A population-based case-control study in Taiwan. Medicine 2016, 95, e2659. [Google Scholar] [CrossRef]
- Rosenblum, S.; Pal, A.; Reidy, K. Renal development in the fetus and premature infant. Semin. Fetal Neonatal Med. 2017, 22, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Uy, N.; Reidy, K. Developmental genetics and congenital anomalies of the kidney and urinary tract. J. Pediatr. Genet. 2015, 5, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaou, N.; Renkema, K.; Bongers, E.M.H.F.; Giles, R.H.; Knoers, N.V.A.M. Genetic, environmental, and epigenetic factors involved in CAKUT. Nat. Rev. Nephrol. 2015, 11, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.; Pollak, M.R. APOL1and kidney disease: From genetics to biology. Annu. Rev. Physiol. 2020, 82, 323–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruggeman, L.A.; O’Toole, J.F.; Sedor, J.R. APOL1 polymorphisms and kidney disease: Loss-of-function or gain-of-function? Am. J. Physiol. Physiol. 2019, 316, F1–F8. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, V.A.; Brenner, B.M. Birth weight, malnutrition and kidney-associated outcomes—A global concern. Nat. Rev. Nephrol. 2015, 11, 135–149. [Google Scholar] [CrossRef]
- Nenov, V.D.; Taal, M.; Sakharova, O.V.; Brenner, B.M. Multi-hit nature of chronic renal disease. Curr. Opin. Nephrol. Hypertens. 2000, 9, 85–97. [Google Scholar] [CrossRef]
- Wood-Bradley, R.J.; Barrand, S.; Giot, A.; Armitage, J. Understanding the role of maternal diet on kidney development; an opportunity to improve cardiovascular and renal health for future generations. Nutrients 2015, 7, 1881–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.Q.; Collins, C.E.; Gordon, A.; Rae, K.M.; Pringle, K.G. the relationship between maternal nutrition during pregnancy and offspring kidney structure and function in humans: A systematic review. Nutrients 2018, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-N.; Tain, Y.-L. The good, the bad, and the ugly of pregnancy nutrients and developmental programming of adult disease. Nutrients 2019, 11, 894. [Google Scholar] [CrossRef] [Green Version]
- Painter, R.C.; Roseboom, T.J.; van Montfrans, G.A.; Bossuyt, P.M.; Krediet, R.T.; Osmond, C.; Barker, D.J.; Bleker, O.P. Microalbuminuria in adults after prenatal exposure to the Dutch famine. J. Am. Soc. Nephrol. 2005, 16, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Miliku, K.; Mesu, A.; Franco, O.; Hofman, A.; Steegers, E.A.; Jaddoe, V.W. Maternal and fetal folate, vitamin B 12, and homocysteine concentrations and childhood kidney outcomes. Am. J. Kidney Dis. 2017, 69, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodyer, P.; Kurpad, A.; Rekha, S.; Muthayya, S.; Dwarkanath, P.; Iyengar, A.; Philip, B.; Mhaskar, A.; Benjamin, A.; Maharaj, S.; et al. Effects of maternal vitamin A status on kidney development: A pilot study. Pediatr. Nephrol. 2007, 22, 209–214. [Google Scholar] [CrossRef]
- Gilbert, J.S.; Lang, A.L.; Grant, A.R.; Nijland, M.J. Maternal nutrient restriction in sheep: Hypertension and decreased nephron number in offspring at 9 months of age. J. Physiol. 2005, 565, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Woods, L.L.; Weeks, D.A.; Rasch, R. Programming of adult blood pressure by maternal protein restriction: Role of nephro-genesis. Kidney Int. 2004, 65, 1339–1348. [Google Scholar] [CrossRef] [Green Version]
- Battista, M.-C.; Oligny, L.L.; St-Louis, J.; Brochu, M. Intrauterine growth restriction in rats is associated with hypertension and renal dysfunction in adulthood. Am. J. Physiol. Metab. 2002, 283, E124–E131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlegel, R.N.; Moritz, K.M.; Paravicini, T.M. Maternal hypomagnesemia alters renal function but does not program changes in the cardiovascular physiology of adult offspring. J. Dev. Orig. Heal. Dis. 2016, 7, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Shi, A.; Zhu, D.; Bo, L.; Zhong, Y.; Wang, J.; Xu, Z.; Mao, C. High sucrose intake during gestation increases angiotensin II type 1 receptor-mediated vascular contractility associated with epigenetic alterations in aged offspring rats. Peptides 2016, 86, 133–144. [Google Scholar] [CrossRef]
- Tain, Y.L.; Wu, K.L.; Lee, W.C.; Leu, S.; Chan, J.Y. Maternal fructose-intake-induced renal programming in adult male off-spring. J. Nutr. Biochem. 2015, 26, 642–650. [Google Scholar] [CrossRef]
- Tain, Y.-L.; Lin, Y.-J.; Sheen, J.-M.; Yu, H.-R.; Tiao, M.-M.; Chen, C.-C.; Tsai, C.-C.; Huang, L.-T.; Hsu, C.-N. High fat diets sex-specifically affect the renal transcriptome and program obesity, kidney injury, and hypertension in the offspring. Nutrients 2017, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- da Silva, A.A.; de Noronha, I.L.; de Oliveira, I.B.; Malheiros, D.M.; Heimann, J.C. Renin-angiotensin system function and blood pressure in adult rats after perinatal salt overload. Nutr. Metab. Cardiovasc. Dis. 2003, 13, 133–139. [Google Scholar] [PubMed]
- Tain, Y.-L.; Hsieh, C.-S.; Lin, I.-C.; Chen, C.-C.; Sheen, J.-M.; Huang, L.-T. Effects of maternal l-citrulline supplementation on renal function and blood pressure in offspring exposed to maternal caloric restriction: The impact of nitric oxide pathway. Nitric Oxide 2010, 23, 34–41. [Google Scholar] [CrossRef]
- Luzardo, R.; Silva, P.A.; Einicker-Lamas, M.; Ortiz-Costa, S.; Carmo, M.D.G.T.D.; Vieira-Filho, L.D.; Paixão, A.D.O.; Lara, L.S.; Vieyra, A. Metabolic programming during lactation stimulates renal na+ transport in the adult offspring due to an early impact on local angiotensin II pathways. PLoS ONE 2011, 6, e21232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelièvre-Pégorier, M.; Vilar, J.; Ferrier, M.-L.; Moreau, E.; Freund, N.; Gilbert, T.; Merlet-Bénichou, C. Mild vitamin A deficiency leads to inborn nephron deficit in the rat. Kidney Int. 1998, 54, 1455–1462. [Google Scholar] [CrossRef] [Green Version]
- Paixão, A.D.; Maciel, C.R.; Teles, M.B.B.; Figueiredo-Silva, J. Regional Brazilian diet-induced low birth weight is correlated with changes in renal hemodynamics and glomerular morphometry in adult age. Biol. Neonate 2001, 80, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Lisle, S.J.M.; Lewis, R.M.; Petry, C.J.; Ozanne, S.E.; Hales, C.N.; Forhead, A.J. Effect of maternal iron restriction during pregnancy on renal morphology in the adult rat offspring. Br. J. Nutr. 2003, 90, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koleganova, N.; Piecha, G.; Ritz, E.; Becker, L.E.; Müller, A.; Weckbach, M.; Nyengaard, J.R.; Schirmacher, P.; Gross-Weissmann, M.-L. Both high and low maternal salt intake in pregnancy alter kidney development in the offspring. Am. J. Physiol. Physiol. 2011, 301, F344–F354. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-N.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.; Tain, Y.-L. Maternal N-acetylcysteine therapy prevents hypertension in spontaneously hypertensive rat offspring: Implications of hydrogen sulfide-generating pathway and gut microbiota. Antioxidants 2020, 9, 856. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sul-fide-generating pathway and renal transcriptome to prevent prenatal N(G)-Nitro-L-arginine methyl ester (L-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obstet. Gynecol. 2016, 215, 636. [Google Scholar] [CrossRef]
- Hsu, C.-N.; Yang, H.-W.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.; Tain, Y.-L. Maternal adenine-induced chronic kidney disease programs hypertension in adult male rat offspring: Implications of nitric oxide and gut microbiome derived metabolites. Int. J. Mol. Sci. 2020, 21, 7237. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.-L.; Lee, W.-C.; Hsu, C.-N.; Lee, W.-C.; Huang, L.-T.; Lee, C.-T.; Lin, C.-Y. Asymmetric dimethylarginine is associated with developmental programming of adult kidney disease and hypertension in offspring of streptozotocin-treated mothers. PLoS ONE 2013, 8, e55420. [Google Scholar] [CrossRef] [PubMed]
- Thomal, M.J.T.; Palma, B.D.; Ponzio, M.B.F.; Franco, M.D.C.; Zaladek-Gil, F.; Fortes, Z.B.; Tufik, S.; Gomes, G.N. Sleep restriction during pregnancy: Hypertension and renal abnormalities in young offspring rats. Sleep 2010, 33, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.; Kitt, J.; Leeson, P.; Aye, C.Y.L.; Lewandowski, A.J. Preeclampsia: Risk factors, diagnosis, management, and the car-diovascular impact on the offspring. J. Clin. Med. 2019, 8, 1625. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.M.; Peck, J.D.; Thompson, D.; Wild, R.A.; Langlois, P. Maternal diabetes and renal agenesis/dysgenesis. Birth Defects Res. Part. A Clin. Mol. Teratol. 2010, 88, 722–727. [Google Scholar] [CrossRef]
- Nelson, R.G.; Morgenstern, H.; Bennett, P.H. Intrauterine diabetes exposure and the risk of renal disease in diabetic Pima Indians. Diabetes 1998, 47, 1489–1493. [Google Scholar] [CrossRef]
- Brennan, S.; Kandasamy, Y.; Rudd, D.M.; Schneider, M.E.; Jones, R.E.; Watson, D.L. The effect of diabetes during pregnancy on fetal renal parenchymal growth. J. Nephrol. 2020, 33, 1079–1089. [Google Scholar] [CrossRef]
- Wong, M.G.; The, N.L.; Glastras, S. Maternal obesity and offspring risk of chronic kidney disease. Nephrology 2018, 23, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.Q.; Collins, C.E.; Gordon, A.; Rae, K.; Pringle, K.G. The relationship between maternal obesity and diabetes during pregnancy on offspring kidney structure and function in humans: A systematic review. J. Dev. Orig. Heal. Dis. 2018, 10, 406–419. [Google Scholar] [CrossRef]
- Macumber, I.; Schwartz, S.; Leca, N. Maternal obesity is associated with congenital anomalies of the kidney and urinary tract in offspring. Pediatr. Nephrol. 2016, 32, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Glastras, S.J.; Tsang, M.; Teh, R.; Chen, H.; McGrath, R.; Zaky, A.A.; Pollock, C.A.; Saad, S. Maternal obesity promotes diabetic nephropathy in rodent offspring. Sci. Rep. 2016, 6, 27769. [Google Scholar] [CrossRef]
- Glastras, S.J.; Chen, H.; Teh, R.; McGrath, R.; Chen, J.; Pollock, C.A.; Wong, M.G.; Saad, S. Mouse models of diabetes, obesity and related kidney disease. PLoS ONE 2016, 11, e0162131. [Google Scholar] [CrossRef] [PubMed]
- Kataria, A.; Trasande, L.; Trachtman, H. The effects of environmental chemicals on renal function. Nat. Rev. Nephrol. 2015, 11, 610–625. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, M.J.; Bolger, P.M.; A Jose, P. The developing kidney and environmental toxins. Pediatrics 2004, 113, 1084–1091. [Google Scholar]
- Weidemann, D.K.; Weaver, V.M.; Fadrowski, J.J. Toxic environmental exposures and kidney health in children. Pediatr. Nephrol. 2015, 31, 2043–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalal, R.P.; Goldfarb, D. Melamine-related kidney stones and renal toxicity. Nat. Rev. Nephrol. 2011, 7, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Skröder, H.; Hawkesworth, S.; Moore, S.E.; Wagatsuma, Y.; Kippler, M.; Vahter, M. Prenatal lead exposure and childhood blood pressure and kidney function. Environ. Res. 2016, 151, 628–634. [Google Scholar] [CrossRef]
- Saylor, C.; Tamayo-Ortiz, M.; Pantic, I.; Amarasiriwardena, C.; McRae, N.; Estrada-Gutierrez, G.; Parra-Hernandez, S.; Tolentino, M.C.; Baccarelli, A.A.; Fadrowski, J.J.; et al. Prenatal blood lead levels and reduced preadolescent glomerular filtration rate: Modification by body mass index. Environ. Int. 2021, 154, 106414. [Google Scholar] [CrossRef]
- Sol, C.M.; Santos, S.; Asimakopoulos, A.G.; Martinez-Moral, M.P.; Duijts, L.; Kannan, K.; Trasande, L.; Jaddoe, V.W. Associations of maternal phthalate and bisphenol urine concentrations during pregnancy with childhood blood pressure in a population-based pro-spective cohort study. Environ. Int. 2020, 138, 105677. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ni, W.; Zhu, S.; Wu, Y.; Cui, Y.; Ma, J.; Liu, Y.; Qiao, J.; Ye, Y.; Yang, P.; et al. Per- and polyfluoroalkyl substances exposure during pregnancy and adverse pregnancy and birth outcomes: A systematic review and meta-analysis. Environ. Res. 2021, 201, 111632. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Wang, L.; Lin, X.; Spengler, J.D.; Perera, F.P. Fetal window of vulnerability to airborne polycyclic aromatic hydro-carbons on proportional intrauterine growth restriction. PLoS ONE 2012, 7, e35464. [Google Scholar]
- Kumar, S.N.; Saxena, P.; Patel, R.; Sharma, A.; Pradhan, D.; Singh, H.; Deval, R.; Bhardwaj, S.K.; Borgohain, D.; Akhtar, N.; et al. Predicting risk of low birth weight offspring from maternal features and blood polycyclic aromatic hydrocarbon concentration. Reprod. Toxicol. 2020, 94, 92–100. [Google Scholar] [CrossRef]
- Uwak, I.; Olson, N.; Fuentes, A.; Moriarty, M.; Pulczinski, J.; Lam, J.; Xu, X.; Taylor, B.D.; Taiwo, S.; Koehler, K.; et al. Application of the navigation guide systematic review methodology to evaluate prenatal exposure to particulate matter air pollution and infant birth weight. Environ. Int. 2021, 148, 106378. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, Y.J.; Lu, P.C.; Tain, Y.L. Maternal resveratrol therapy protects male rat offspring against programmed hyper-tension induced by TCDD and dexamethasone exposures: Is it relevant to aryl hydrocarbon receptor? Int. J. Mol. Sci. 2018, 19, 2459. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Lin, Y.J.; Tain, Y.L. Maternal exposure to bisphenol A combined with high-fat diet-induced programmed hyper-tension in adult male rat offspring: Effects of resveratrol. Int. J. Mol. Sci. 2019, 20, 4382. [Google Scholar] [CrossRef] [Green Version]
- Aragon, A.C.; Kopf, P.G.; Campen, M.J.; Huwe, J.K.; Walker, M.K. In Utero and Lactational 2,3,7,8-Tetrachlorodibenzo-p-dioxin exposure: Effects on fetal and adult cardiac gene expression and adult cardiac and renal morphology. Toxicol. Sci. 2007, 101, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Jacquillet, G.; Barbier, O.; Rubera, I.; Tauc, M.; Borderie, A.; Namorado, M.C.; Martin, D.; Sierra, G.; Reyes, J.L.; Poujeol, P.; et al. Cadmium causes delayed effects on renal function in the offspring of cadmium-contaminated pregnant female rats. Am. J. Physiol. Physiol. 2007, 293, F1450–F1460. [Google Scholar] [CrossRef]
- A Slotkin, T. Cholinergic systems in brain development and disruption by neurotoxicants: Nicotine, environmental tobacco smoke, organophosphates. Toxicol. Appl. Pharmacol. 2004, 198, 132–151. [Google Scholar] [CrossRef]
- Correia-Costa, L.; Schaefer, F.; Afonso, A.C.; Correia, S.; Guimarães, J.T.; Guerra, A.; Barros, H.; Azevedo, A. Prenatal alcohol exposure affects renal function in overweight schoolchildren: Birth cohort analysis. Pediatr. Nephrol. 2019, 35, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; McIntyre, H.D.; Alati, R.; Al Mamun, A. Maternal alcohol consumption during pregnancy and its association with offspring renal function at 30 years: Observation from a birth cohort study. Nephrology 2017, 24, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, S.P.; Denton, K.; Cullen-McEwen, L.; Bertram, J.; Moritz, K.M. Prenatal exposure to alcohol reduces nephron number and raises blood pressure in progeny. J. Am. Soc. Nephrol. 2010, 21, 1891–1902. [Google Scholar] [CrossRef] [Green Version]
- Taal, H.R.; Geelhoed, J.J.M.; Steegers, E.A.P.; Hofman, A.; Moll, H.A.; Lequin, M.; Van Der Heijden, A.J.; Jaddoe, V.W.V. Maternal smoking during pregnancy and kidney volume in the offspring: The generation R study. Pediatr. Nephrol. 2011, 26, 1275–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, D.B.; Mesquita, F.F.; De Lima, I.P.; Boer, P.A.; Gontijo, J.A. Fetal kidney programming by maternal smoking exposure: Effects on kidney structure, blood pressure and urinary sodium excretion in adult offspring. Nephron 2015, 129, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M.; Chou, H.-C.; Huang, L.-T. Maternal nicotine exposure during gestation and lactation induces kidney injury and fibrosis in rat offspring. Pediatr. Res. 2014, 77, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Chan, Y.L.; Nguyen, L.T.; Mak, C.; Zaky, A.; Anwer, A.G.; Shi, Y.; Nguyen, T.; Pollock, C.A.; Oliver, B.G.; et al. Impact of maternal e-cigarette vapor exposure on renal health in the offspring. Ann. N.Y. Acad. Sci. 2019, 1452, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Bundy, J.D.; Bazzano, L.A.; Xie, D.; Cohan, J.; Dolata, J.; Fink, J.C.; Hsu, C.-Y.; Jamerson, K.; Lash, J.; Makos, G.; et al. Self-reported tobacco, alcohol, and illicit drug use and progression of chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2018, 13, 993–1001. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W. Intrauterine infection and preterm delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG Int. J. Obstet. Gynaecol. 2006, 113, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Weckman, A.M.; Ngai, M.; Wright, J.; McDonald, C.R.; Kain, K.C. The impact of infection in pregnancy on placental vascular development and adverse birth outcomes. Front. Microbiol. 2019, 10, 1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Cui, J.; Chen, R.; Deng, Y.; Liao, X.; Wei, Y.; Li, X.; Su, M.; Yu, J.; Yi, P. prenatal exposure to lipopolysaccharide alters renal DNA methyltransferase expression in rat offspring. PLoS ONE 2017, 12, e0169206. [Google Scholar] [CrossRef]
- Farias, J.S.; Santos, K.M.; Lima, N.K.; Cabral, E.V.; Aires, R.S.; Veras, A.C.; Paixão, A.D.; Vieira, L.D. Maternal endotoxemia induces renal collagen deposition in adult offspring: Role of NADPH oxidase/TGF-β1/MMP-2 signaling pathway. Arch. Biochem. Biophys. 2020, 684, 108306. [Google Scholar] [CrossRef]
- Chen, C.-M.; Chou, H.-C. Maternal inflammation exacerbates neonatal hyperoxia-induced kidney injury in rat offspring. Pediatr. Res. 2019, 86, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Fillion, M.-L.; Watt, C.; Gupta, I.R. Vesicoureteric reflux and reflux nephropathy: From mouse models to childhood disease. Pediatr. Nephrol. 2014, 29, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.; Fletcher, J.T.; Alexander, S.I.; Craig, J.C. Vesicoureteral reflux. J. Am. Soc. Nephrol. 2008, 19, 847–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreuder, M.F.; Bueters, R.R.; Huigen, M.C.; Russel, F.G.; Masereeuw, R.; van den Heuvel, L.P. Effect of drugs on renal de-velopment. Clin. J. Am. Soc. Nephrol. 2011, 6, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slabiak-Blaz, N.; Adamczak, M.; Gut, N.; Grajoszek, A.; Nyengaard, J.R.; Ritz, E.; Wiecek, A. Administration of cyclosporine a in pregnant rats—The effect on blood pressure and on the glomerular number in their offspring. Kidney Blood Press. Res. 2015, 40, 413–423. [Google Scholar] [CrossRef]
- Gilbert, T.; Lelievre-Pegorier, M.; Merlet-Benichou, C. Immediate and long-term renal effects of fetal exposure to gentamicin. Pediatr. Nephrol. 1990, 4, 445–450. [Google Scholar] [CrossRef]
- Tain, Y.-L.; Sheen, J.M.; Chen, C.C.; Yu, H.-R.; Tiao, M.-M.; Kuo, H.C.; Huang, L.T. Maternal citrulline supplementation prevents prenatal dexamethasone-induced programmed hypertension. Free. Radic. Res. 2014, 48, 580–586. [Google Scholar] [CrossRef]
- Tai, I.-H.; Sheen, J.-M.; Lin, Y.-J.; Yu, H.-R.; Tiao, M.-M.; Chen, C.-C.; Huang, L.-T.; Tain, Y.-L. Maternal N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and prevents programmed hypertension in male offspring exposed to prenatal dexamethasone and postnatal high-fat diet. Nitric Oxide 2015, 53, 6–12. [Google Scholar] [CrossRef]
- Chang, H.-Y.; Tain, Y.-L. Postnatal dexamethasone-induced programmed hypertension is related to the regulation of melatonin and its receptors. Steroids 2016, 108, 1–6. [Google Scholar] [CrossRef]
- Berl, T. Review: Renal protection by inhibition of the renin-angiotensin-aldosterone system. J. Renin-Angiotensin-Aldosterone Syst. 2009, 10, 1–8. [Google Scholar] [CrossRef]
- Quan, A. Fetopathy associated with exposure to angiotensin converting enzyme inhibitors and angiotensin receptor antago-nists. Early Hum. Dev. 2006, 82, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kett, M.M.; Denton, K. Renal programming: Cause for concern? Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R791–R803. [Google Scholar] [CrossRef] [PubMed]
- McGoldrick, E.; Stewart, F.; Parker, R.; Dalziel, S.R. Antenatal corticosteroids for accelerating fetal lung maturation for women at risk of preterm birth. Cochrane Database Syst. Rev. 2020, 2021, CD004454. [Google Scholar] [CrossRef]
- Moisiadis, V.G.; Matthews, S. Glucocorticoids and fetal programming part 2: Mechanisms. Nat. Rev. Endocrinol. 2014, 10, 403–411. [Google Scholar] [CrossRef]
- Bonzini, M.; Palmer, K.T.; Coggon, D.; Carugno, M.; Cromi, A.; Ferrario, M.M. Shift work and pregnancy outcomes: A sys-tematic review with meta-analysis of currently available epidemiological studies. Int. J. Obstet. Gynaecol. 2011, 118, 1429–1437. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Vandermeer, B.; Khurana, R.; Nerenberg, K.; Featherstone, R.; Sebastianski, M.; Davenport, M.H. The impact of oc-cupational shift work and working hours during pregnancy on health outcomes: A systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2019, 221, 563–576. [Google Scholar] [CrossRef]
- Hsu, C.-N.; Tain, Y.-L. Light and circadian signaling pathway in pregnancy: Programming of adult health and disease. Int. J. Mol. Sci. 2020, 21, 2232. [Google Scholar] [CrossRef] [Green Version]
- Mendez, N.; Torres-Farfan, C.; Salazar, E.; Bascur, P.; Bastidas, C.; Vergara, K.; Spichiger, C.; Halabi, D.; Vio, C.; Richter, H.G. Fetal programming of renal dysfunction and high blood pressure by chronodisruption. Front. Endocrinol. 2019, 10, 362. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Developmental origins of kidney disease: Why oxidative stress matters? Antioxidants 2021, 10, 33. [Google Scholar] [CrossRef]
- Hsu, C.-N.; Tain, Y.-L. Regulation of nitric oxide production in the developmental programming of hypertension and kidney disease. Int. J. Mol. Sci. 2019, 20, 681. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-N.; Tain, Y.-L. Targeting the renin–angiotensin–aldosterone system to prevent hypertension and kidney disease of developmental origins. Int. J. Mol. Sci. 2021, 22, 2298. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-N.; Hou, C.-Y.; Hsu, W.-H.; Tain, Y.-L. Cardiovascular diseases of developmental origins: Preventive aspects of gut microbiota-targeted therapy. Nutrients 2021, 13, 2290. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.P.; Al-Hasan, Y. Impact of oxidative stress in fetal programming. J. Pregnancy 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lee, W.C.; Wu, K.L.H.; Leu, S.; Chan, J.Y.H. Targeting arachidonic acid pathway to prevent programmed hyper-tension in maternal fructose-fed male adult rat offspring. J. Nutr. Biochem. 2016, 38, 86–92. [Google Scholar] [CrossRef]
- Tain, Y.L.; Chan, J.Y.H.; Lee, C.T.; Hsu, C.N. Maternal melatonin therapy attenuates methyl-donor diet-induced pro-grammed hypertension in male adult rat offspring. Nutrients 2018, 10, 1407. [Google Scholar] [CrossRef] [Green Version]
- Sukjamnong, S.; Chan, Y.L.; Zakarya, R.; Nguyen, L.T.; Anwer, A.G.; Zaky, A.A.; Santiyanont, R.; Oliver, B.G.; Goldys, E.; Pollock, C.A.; et al. MitoQ supplementation prevent long-term impact of maternal smoking on renal development, oxidative stress and mitochondrial density in male mice offspring. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jeje, S.; O Akindele, O.; Ushie, G.; Rajil, Y. Changes in kidney function and oxidative stress biomarkers in offspring from dams treated with dexamethasone during lactation in Wistar rats. Afr. J. Med. Med. Sci. 2016, 45, 237–242. [Google Scholar]
- Sladek, S.M.; Magness, R.R.; Conrad, K.P. Nitric oxide and pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 1997, 272, R441–R463. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.; Hsu, C. Toxic dimethylarginines: Asymmetric dimethylarginine (ADMA) and symmetric dimethylarginine (SDMA). Toxins 2017, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Gubler, M.C.; Antignac, C. Renin–angiotensin system in kidney development: Renal tubular dysgenesis. Kidney Int. 2010, 77, 400–406. [Google Scholar] [CrossRef] [Green Version]
- Yosypiv, I.V. Renin-angiotensin system in mammalian kidney development. Pediatr. Nephrol. 2020, 36, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Deelman, L.; Sharma, K. Mechanisms of kidney fibrosis and the role of antifibrotic therapies. Curr. Opin. Nephrol. Hypertens. 2009, 18, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Benigni, A.; Cassis, P.; Remuzzi, G. Angiotensin II revisited: New roles in inflammation, immunology and aging. EMBO Mol. Med. 2010, 2, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain–gut–kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Chang-Chien, G.P.; Lin, S.; Hou, C.Y.; Tain, Y.L. Targeting on gut microbial metabolite trimethylamine-N-oxide and short-chain fatty acid to prevent maternal high-fructose-diet-induced developmental programming of hypertension in adult male offspring. Mol. Nutr. Food Res. 2019, 63, e1900073. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.S.; Borges, N.A.; Esgalhado, M.; Magliano, D.C.; Soulage, C.O.; Mafra, D. Aryl hydrocarbon receptor activation in chronic kidney disease: Role of uremic toxins. Nephron 2017, 137, 1–7. [Google Scholar] [CrossRef]
- Richards, E.M.; Pepine, C.J.; Raizada, M.K.; Kim, S. The gut, its microbiome, and hypertension. Curr. Hypertens. Rep. 2017, 19, 36. [Google Scholar] [CrossRef] [Green Version]
- Oliveira Andrade, J.M.; de Farias Lelis, D.; Mafra, V.; Cota, J. The angiotensin converting enzyme 2 (ACE2), gut microbiota, and cardiovascular health. Protein Pept. Lett. 2017, 24, 827–832. [Google Scholar] [CrossRef]
- Bianco-Miotto, T.; Craig, J.M.; Gasser, Y.P.; van Dijk, S.J.; Ozanne, S.E. Epigenetics and DOHaD: From basics to birth and beyond. J. Dev. Orig. Health Dis. 2017, 8, 513–519. [Google Scholar] [CrossRef]
- Jansson, T.; Powell, T. Role of placental nutrient sensing in developmental programming. Clin. Obstet. Gynecol. 2013, 56, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Tomat, A.L.; Salazar, F.J. Mechanisms involved in developmental programming of hypertension and renal diseases. Gender differences. Horm. Mol. Biol. Clin. Investig. 2014, 18, 63–77. [Google Scholar] [CrossRef]
- Beluska-Turkan, K.; Korczak, R.; Hartell, B.; Moskal, K.; Maukonen, J.; Alexander, D.E.; Salem, N.; Harkness, L.; Ayad, W.; Szaro, J.; et al. Nutritional gaps and supplementation in the first 1000 days. Nutrients 2019, 11, 2891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollmann, T.R.; Marchant, A.; Way, S.S. Vaccination strategies to enhance immunity in neonates. Science 2020, 368, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Htay, H.; Alrukhaimi, M.; Ashuntantang, G.E.; Bello, A.K.; Bellorin-Font, E.; Gharbi, M.B.; Braam, B.; Feehally, J.; Harris, D.C.; Jha, V.; et al. Global access of patients with kidney disease to health technologies and medications: Findings from the Global Kidney Health Atlas project. Kidney Int. Suppl. 2018, 8, 64–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrezenmeier, E.V.; Barasch, J.; Budde, K.; Westhoff, T.; Schmidt-Ott, K.M. Biomarkers in acute kidney injury—Pathophysi-ological basis and clinical performance. Acta Physiol. 2017, 219, 554–572. [Google Scholar] [CrossRef]
- Ronco, C.; Bellomo, R.; Kellum, J.A. Acute kidney injury. Lancet 2019, 394, 1949–1964. [Google Scholar] [CrossRef]
- Parmaksız, G.; Noyan, A.; Dursun, H.; Ince, E.; Anarat, R.; Cengiz, N. Role of new biomarkers for predicting renal scarring in vesicoureteral reflux: NGAL, KIM-1, and L-FABP. Pediatr. Nephrol. 2015, 31, 97–103. [Google Scholar] [CrossRef]
- Anand, S.; Bajpai, M.; Khanna, T.; Kumar, A. Urinary biomarkers as point-of-care tests for predicting progressive deterioration of kidney function in congenital anomalies of kidney and urinary tract: Trefoil family factors (TFFs) as the emerging biomarkers. Pediatr. Nephrol. 2021, 36, 1465–1472. [Google Scholar] [CrossRef]
- Woolf, A.S. Growing a new human kidney. Kidney Int. 2019, 96, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Franco Mdo, C.; Ponzio, B.F.; Gomes, G.N.; Gil, F.Z.; Tostes, R.; Carvalho, M.H.; Fortes, Z.B. Micronutrient prenatal supple-mentation prevents the development of hypertension and vascular endothelial damage induced by intrauterine malnutrition. Life Sci. 2009, 85, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Torrens, C.; Brawley, L.; Anthony, F.W.; Dance, C.S.; Dunn, R.; Jackson, A.A.; Poston, L.; Hanson, M.A. Folate supplementa-tion during pregnancy improves offspring cardiovascular dysfunction induced by protein restriction. Hypertension 2006, 47, 982–987. [Google Scholar] [CrossRef] [Green Version]
- Thaeomor, A.; Teangphuck, P.; Chaisakul, J.; Seanthaweesuk, S.; Somparn, N.; Roysommuti, S. Perinatal taurine supplementation prevents metabolic and cardiovascular effects of maternal diabetes in adult rat offspring. Adv. Exp. Med. Biol. 2017, 975, 295–305. [Google Scholar] [PubMed]
- Hsu, C.N.; Lin, I.C.; Yu, H.R.; Huang, L.T.; Tiao, M.M.; Tain, Y.L. Maternal tryptophan supplementation protects adult rat offspring against hypertension programmed by maternal chronic kidney disease: Implication of tryptophan-metabolizing mi-crobiome and aryl hydrocarbon receptor. Int J. Mol. Sci. 2020, 21, 4552. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. The double-edged sword effects of maternal nutrition in the developmental programming of hyper-tension. Nutrients 2018, 10, 1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.-N.; Tain, Y.-L. Gasotransmitters for the therapeutic prevention of hypertension and kidney disease. Int. J. Mol. Sci. 2021, 22, 7808. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.-L.; Hsu, C.-N. AMP-activated protein kinase as a reprogramming strategy for hypertension and kidney disease of developmental origin. Int. J. Mol. Sci. 2018, 19, 1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United Nations. Report of the Secretary-General on SDG Progress Special Edition; United Nations: New York, NY, USA, 2019.
Figure 1.
Kidney development during the first 1000 days of life. UB = ureteric bud; GFR = glomerular filtration rate.
Figure 1.
Kidney development during the first 1000 days of life. UB = ureteric bud; GFR = glomerular filtration rate.

Figure 2.
Various environmental factors during the first 1000 days of life are associated with renal programming, resulting in kidney disease in later life. These risk factors include nutritional imbalance, maternal illness, environmental chemicals, infection and inflammation, medication use, and chronodisruption.
Figure 2.
Various environmental factors during the first 1000 days of life are associated with renal programming, resulting in kidney disease in later life. These risk factors include nutritional imbalance, maternal illness, environmental chemicals, infection and inflammation, medication use, and chronodisruption.

Figure 3.
Overview of the common mechanisms of renal programming in response to various environmental risk factors during the first 1000 days of life. NO = nitric oxide; RAS = renin–angiotensin system.
Figure 3.
Overview of the common mechanisms of renal programming in response to various environmental risk factors during the first 1000 days of life. NO = nitric oxide; RAS = renin–angiotensin system.

Figure 4.
A summary of prevention strategies to improve kidney health during the first 1000 days of life.
Figure 4.
A summary of prevention strategies to improve kidney health during the first 1000 days of life.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hsu, C.-N.; Tain, Y.-L. The First Thousand Days: Kidney Health and Beyond. Healthcare 2021, 9, 1332. https://doi.org/10.3390/healthcare9101332
AMA Style
Hsu C-N, Tain Y-L. The First Thousand Days: Kidney Health and Beyond. Healthcare. 2021; 9(10):1332. https://doi.org/10.3390/healthcare9101332
Chicago/Turabian StyleHsu, Chien-Ning, and You-Lin Tain. 2021. "The First Thousand Days: Kidney Health and Beyond" Healthcare 9, no. 10: 1332. https://doi.org/10.3390/healthcare9101332
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.