Realistic Choice of Annual Matrices Contracts the Range of λS Estimates



Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Study of Androsace albana
- -
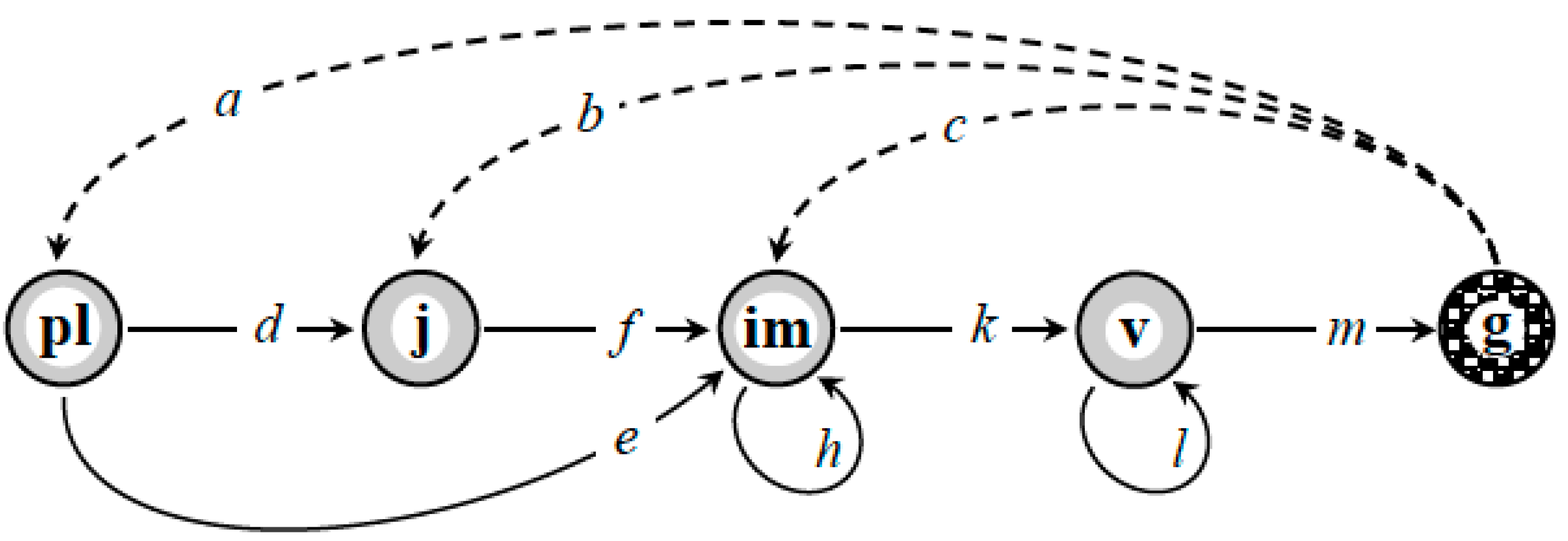
- delays in stages im and v, which can be explained by the fact that the harsh conditions of the highlands force the plants to resort to the “space-holder strategy” [30], i.e., staying or growing in one place for as long as possible. Poor soil quality also results in some virginal plants accumulating resources for fruiting longer than one year [30,31,32,33,34];
- -
- accelerated transitions
![Mathematics 08 02252 i001]() as a manifestation of polyvariant ontogeny in A. albana under the alpine belt conditions in South-Western Caucasus.
as a manifestation of polyvariant ontogeny in A. albana under the alpine belt conditions in South-Western Caucasus.

2.2. Local Meteodata, Statistical Treatment
2.3. Revealing the Pattern of Transition Matrix and Estimating Its Elements
2.4. Estimating λS by the Direct MC Method with a Markov Chain
3. Results
3.1. The Pattern of P = [pij] and the Estimates of pij
3.2. Estimates of λS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Tossing a 10-Face Dice with Unequal Face Probabilities
Appendix B. Calculating the Final Term of a Given-Length Sequence
| function [lamS] = lamS_Ana_num(All,Tau, coef) % function lamS_Ana_num calculates stochastic lambda following definition (2) % input All is a 3D array of all numeric annual matrices available in the study. % in 2020, all = 10; Tau= Length of the product % random choice is governed by a Markov chain with an m-by-m matrix P (see below) % transition matrix Pnum is a global numeric variable. % @ Logofet D.O., 2020 global Pnum; size(All);m=ans(3); % removed all checks for correct arguments! tau=1; Column=Pnum(:,1); xPROD = [37 110 99 35 13]’;% initial vector = x2009 for A.albana while (tau <= Tau), % length of the product = Tau row = (m+1)-sum(rand<=cumsum(Column)); % random choice from ‘m’ annual matrices Ltau = All(:,:,row); % randomly (MC) chosen current matrix. xPROD = double(Ltau * xPROD/coef); % double precision calculation. tau=tau+1;Column=Pnum(:,row); end; lamS = (exp(log(norm(xPROD, 1))/Tau + log(coef))); end |
Appendix C. Getting a Given Number of Random Realizations
| ffunction [lamSmin, lamSmax] = repAnaMCnum(repeat, AllLnum, Tau, coef) % repeats calculation of stochastic lambda for ‘repeat’ times (=13, 33, …) % and detects the range of variations for A.albana (‘Ana’ in the function name); % the rest 2 input arguments are the same as in function ‘lamS_Ana_num’(MC). % @ Logofet D.O., 2020 size(repeat); rng(‘default’); % to reproduce the results. if any(size(repeat)~=[1 1]), error ‘Incorrect size of the input!’, end; first=[]; for rep=1:repeat, lamS=lamS_Ana_num(AllLnum,Tau, coef);% next lambdaS, which may happen = 0; if lamS==0, lamS=[]; end;% excludes 0 lambdaS from accumulation in ‘first’ first=[first; lamS];% adjoins the next nonzero lambdaS; if rem(rep,10)==0, rep, min(first),max(first),end,% for vision during the long calculation end; format long; lamSmin=min(first); lamSmax=max(first);%bounds of the range end |
References
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Logofet, D.O. Projection matrices revisited: A potential-growth indicator and the merit of indication. J. Math. Sci. 2013, 193, 671–686. [Google Scholar] [CrossRef]
- Harary, F.; Norman, R.Z.; Cartwright, D. Structural Models: An Introduction to the Theory of Directed Graphs; John Wiley: New York, NY, USA, 1965. [Google Scholar]
- Horn, R.A.; Johnson, C.R. Matrix Analysis; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Logofet, D.O. Matrices and Graphs: Stability Problems in Mathematical Ecology; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Gantmacher, F.R. Matrix Theory; Chelsea Publ.: New York, NY, USA, 1959. [Google Scholar]
- Logofet, D.O.; Ulanova, N.G.; Belova, I.N. Adaptation on the ground and beneath: Does the local population maximize its λ1? Ecol. Complex. 2014, 20, 176–184. [Google Scholar] [CrossRef]
- Logofet, D.O. Projection matrices in variable environments: λ1 in theory and practice. Ecol. Model. 2013, 251, 307–311. [Google Scholar] [CrossRef]
- Furstenberg, H.; Kesten, H. Products of random matrices. Ann. Math. Stat. 1960, 31, 457–469. [Google Scholar] [CrossRef]
- Oseledec, V.I. A multiplicative ergodic theorem: Ljapunov characteristic numbers for dynamical systems. Trans. Mosc. Math. Soc. 1968, 19, 197–231. [Google Scholar]
- Cohen, J.E. Ergodicity of age structure in populations with Markovian vital rates, I: Countable states. J. Amer. Stat. Ass. 1976, 71, 335–339. [Google Scholar] [CrossRef]
- Tuljapurkar, S.D. Population Dynamics in Variable Environments; Springer: New York, NY, USA, 1990. [Google Scholar]
- Logofet, D.O. Does averaging overestimate or underestimate population growth? It depends. Ecol. Model. 2019, 411, 108744. [Google Scholar] [CrossRef]
- Richardson, C.W. Stochastic simulation of daily precipitation, temperature and solar radiation. Water Resour. Res. 1981, 17, 182–190. [Google Scholar] [CrossRef]
- Johnson, G.L.; Hanson, C.L.; Hardegree, S.P.; Ballard, E.B. Stochastic weather simulation—Overview and analysis of two commonly used models. J. Appl. Meteorol. 1996, 35, 1878–1896. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.L.; Daly, C.; Taylor, G.H.; Hanson, C.L. Spatial variability and interpolation of stochastic weather simulation model parameters. J. Appl. Meteorol. 2000, 39, 778–796. [Google Scholar] [CrossRef]
- Dubrovsky, M.; Zalud, Z.; Stastna, M. Sensitivity of CERES–Maize yields to statistical structure of daily weather series. Clim. Chang. 2000, 46, 447–472. [Google Scholar] [CrossRef]
- Cohen, J.E. Ergodicity of age structure in populations with Markovian vital rates, II: General states. Adv. Appl. Probab. 1977, 9, 18–37. [Google Scholar] [CrossRef]
- Steinsaltz, D.; Tuljapurkar, S.; Horvitz, C. Derivatives of the stochastic growth rate. Theor. Popul. Biol. 2011, 80, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, L. Conditions for growth and extinction in matrix models with environmental stochasticity. Ecol. Model. 2019, 411, 108797. [Google Scholar] [CrossRef]
- Morris, W.F.; Tuljapurkar, S.; Haridas, C.V.; Menges, E.S.; Horvitz, C.C.; Pfister, C.A. Sensitivity of the population growth rate to demographic variability within and between phases of the disturbance cycle. Ecol. Lett. 2006, 9, 1331–1341. [Google Scholar] [CrossRef]
- Rees, M.; Ellner, S.P. Integral projection models for populations in temporally varying environments. Ecol. Monogr. 2009, 79, 575–594. [Google Scholar] [CrossRef] [Green Version]
- Ozgul, A.; Childs, D.Z.; Oli, M.K.; Armitage, K.B.; Blumstein, D.T.; Olson, L.E.; Tuljapurkar, S.; Coulson, T. Coupled dynamics of body mass and population growth in response to environmental change. Nature 2010, 466, 482–485. [Google Scholar] [CrossRef] [Green Version]
- Williams, H.J.; Jacquemyn, H.; Ochocki, B.M.; Brys, R.; Miller, T.E.X. Life history evolution under climate change and its influence on the population dynamics of a long-lived plant. J. Ecol. 2015, 103, 798–808. [Google Scholar] [CrossRef]
- Red Book of the Adygea Republic: Rare and Endangered Objects of Fauna and Flora: In 2 Parts, Part 1: Plants and Fungi, 2nd ed.; Zamotaylov, A.S. (Ed.) Kachestvo: Maykop, Russia, 2012. (In Russian) [Google Scholar]
- Red Book of the Krasnodar Territory (Plants and Mushrooms), 3nd ed.; Litvinskaya, S.A. (Ed.) Design Bureau No. 1: Krasnodar, Russia, 2017. (In Russian) [Google Scholar]
- Kazantseva, E.S. Population Dynamics and Seed Productivity of Short-Lived Alpine Plants in the North-West Caucasus. Ph.D Thesis, Moscow State University, Moscow, Russia, 2016. (In Russian). [Google Scholar]
- Logofet, D.O.; Kazantseva, E.S.; Belova, I.N.; Onipchenko, V.G. How long does a short-lived perennial live? A modelling approach. Biol. Bull. Rev. 2018, 8, 406–420. [Google Scholar] [CrossRef]
- Logofet, D.O.; Kazantseva, E.S.; Belova, I.N.; Onipchenko, V.G. Local population of Eritrichium caucasicum as an object of mathematical modelling. I. Life cycle graph and a nonautonomous matrix model. Biol. Bull. Rev. 2017, 7, 415–427. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Rabonov, T.A. Life cycle of perennial herbaceous plants in meadow phytocoenoses. Trudi Bot. Inst. Acad. Nauk SSSR Ser. 3. Geobot. 1950, 6, 7–204. (In Russian) [Google Scholar]
- Rabotnov, T.A. Fitotsenologiya (Phytocenology); Moscow State Univ. Publ.: Moscow, Russia, 1978. (In Russian) [Google Scholar]
- Bender, M.H.; Baskin, J.M.; Baskin, C.C. Age of maturity and life span in herbaceous, polycarpic perennials. Bot. Rev. 2000, 66, 311–349. [Google Scholar] [CrossRef]
- Keller, R.; Vittoz, P. Clonal growth and demography of a hemicryptophyte alpine plant: Leontopodium alpinum Cassini. Alp. Bot. 2015, 125, 31–40. [Google Scholar] [CrossRef]
- Logofet, D.O.; Kazantseva, E.S.; Onipchenko, V.G. Seed bank as a persistent problem in matrix population models: From uncertainty to certain bounds. Ecol. Model. 2020, 438, 109284. [Google Scholar] [CrossRef]
- Logofet, D.O.; Kazantseva, E.S.; Belova, I.N.; Onipchenko, V.G. Backward prediction confirms the conclusion on local plant population viability. Zhurnal Obs. Biol. (J. Gen. Biol.) 2020, 81, 257–271. (In Russian) [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-128. Available online: http://CRAN.R-project.org/package=nlme (accessed on 20 October 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 20 October 2020).
- Kemeny, J.G.; Snell, J.L. Finite Markov Chains; Van Nostrand: Princeton, NJ, USA, 1960. [Google Scholar]
- Logofet, D.O.; Kazantseva, E.S.; Belova, I.N.; Onipchenko, V.G. Local population of Eritrichium caucasicum as an object of mathematical modelling. III. Population growth in the random environment. Biol. Bull. Rev. 2019, 9, 453–464. [Google Scholar] [CrossRef]
- Nguyen, V.; Buckley, Y.M.; Salguero-Gomez, R.; Wardle, G.M. Consequences of neglecting cryptic life stages from demographic models. Ecol. Model. 2019, 408, 108723. [Google Scholar] [CrossRef]
- Kendall, B.E.; Fujiwara, M.; Diaz-Lopez, J.; Schneider, S.; Voigt, J.; Wiesner, S. Persistent problems in the construction of matrix population models. Ecol. Model. 2019, 406, 33–43. [Google Scholar] [CrossRef]
- Che-Castaldo, J.; Jones, O.; Kendall, B.E.; Burns, J.H.; Childs, D.Z.; Ezard, T.H.; Hernandez-Yanez, H.; Hodgson, D.J.; Jongejans, E.; Knight, T.; et al. Comments to “Persistent problems in the construction of matrix population models”. Ecol. Model. 2020, 416, 108913. [Google Scholar] [CrossRef] [Green Version]
- Tuljapurkar, S.D.; Stanford University, Stanford, CA, USA. Personal Communication, 2020.
- Tuljapurkar, S.D.; Haridas, C.V. Temporal autocorrelation and stochastic population growth. Ecol. Lett. 2006, 9, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Morris, W.F.; Pfister, C.A.; Tuljapurkar, S.; Haridas, C.V.; Boggs, C.L.; Boyce, M.S.; Bruna, E.M.; Church, D.R.; Coulson, T.; Doak, D.F.; et al. Longevity can buffer plant and animal populations against changing climatic variability. Ecology 2008, 89, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, C.P.; Casella, G. Monte Carlo Statistical Methods; Springer: New York, NY, USA, 2004. [Google Scholar]
- Mathworks Documentation. Available online: https://www.mathworks.com/help/matlab/ref/rand.html?s_tid=srchtitle (accessed on 20 October 2020).
- Mathworks Documentation. Available online: https://www.mathworks.com/help/matlab/ref/global.html (accessed on 20 October 2020).
- Mathworks Documentation. Available online: https://www.mathworks.com/help/matlab/ref/rng.html (accessed on 20 October 2020).




{kind=link}
{kind=link}
{kind=link}
| Stage | Stage Group Sizes (in Absolute Numbers) at the Year of Observation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | |
| pl | 37 | 30 | 19 | 49 | 19 | 16 | 4 | 10 | 3 | 12 | 13 |
| j | 110 | 48 | 45 | 86 | 137 | 98 | 19 | 29 | 8 | 23 | 38 |
| im | 99 | 55 | 43 | 87 | 95 | 34 | 10 | 13 | 4 | 13 | 2 |
| v | 35 | 26 | 57 | 58 | 73 | 50 | 20 | 16 | 18 | 23 | 23 |
| g | 13 | 1 | 1 | 4 | 6 | 3 | 4 | 2 | 1 | 2 | 1 |
| Census Year, t | Matrix L(t): t → t + 1 | λ1(L(t)) | Vector x*, % |
|---|---|---|---|
| 2009 | 0.5661 | ||
| 2010 | 1.2283 | ||
| 2011 | 1.5779 | ||
| 2012 | 1.2641 | ||
| 2013 | 0.6345 | ||
| 2014 | 0.3988 | ||
| 2015 | 1.0679 | ||
| 2016 | 0. 9611 | ||
| 2017 | 1.1206 | ||
| 2018 | 0.9617 |
| Incoming States | Outgoing States | Eigenvector, ss* | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | ||
| 2009 | 0 | 1/8 | 0 | 1/9 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0359 |
| 2010 | 1/2 | 1/4 | 0 | 1/9 | 1/3 | 0 | 0 | 1/9 | 0 | 1/3 | 0.1437 |
| 2011 | 0 | 1/4 | 0 | 0 | 0 | 0 | 0 | 0 | 1/2 | 0 | 0.0359 |
| 2012 | 0 | 0 | 1/2 | 1/9 | 1/3 | 1/4 | 0 | 0 | 0 | 1/3 | 0.1613 |
| 2013 | 0 | 0 | 0 | 1/9 | 0 | 1/8 | 1/7 | 0 | 0 | 0 | 0.0519 |
| 2014 | 0 | 1/8 | 1/2 | 1/9 | 1/3 | 0 | 2/7 | 1/9 | 0 | 1/6 | 0.1389 |
| 2015 | 0 | 1/8 | 0 | 1/9 | 0 | 1/8 | 0 | 1/3 | 0 | 1/6 | 0.1164 |
| 2016 | 1/2 | 0 | 0 | 0 | 0 | 1/4 | 2/7 | 1/3 | 0 | 0 | 0.1289 |
| 2017 | 0 | 0 | 0 | 1/9 | 0 | 1/8 | 0 | 1/9 | 1/4 | 0 | 0.0661 |
| 2018 | 0 | 1/8 | 0 | 2/9 | 0 | 1/8 | 2/7 | 0 | 1/4 | 0 | 0.1210 |
| Column Sum | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Product “Length” 1 | Number of Realizations | Range of Variations in the Estimates of λS; Range Length in the Following Series: | ||
|---|---|---|---|---|
| Markov Chain | Equiprobable iid | ss* iid | ||
| 1 × 105 | 13 | [0.924332, 0.928207]; 0.003876 | [0.935773, 0.939521]; 0.003749 | [0.940098, 0.945336]; 0.005237 |
| 33 | [0.924332, 0.928207]; 0.003876 | [0.933281, 0.939521]; 0.006240 | [0.939897, 0.945336]; 0.005439 | |
| 100 | [0.923628, 0.928371]; 0.004743 | [0.933281, 0.940469]; 0.007188 | [0.939383, 0.945336]; 0.005953 | |
| 2 × 105 | 13 | [0.925053, 0.927760]; 0.002707 | [0.935810, 0.938245]; 0.002434 | [0.940534, 0.943727]; 0.003193 |
| 33 | [0.924339, 0.927760]; 0.003420 | [0.935694, 0.93465]; 0.002770 | [0.940534, 0.943727]; 0.003193 | |
| 100 | [0.924046, 0.927760]; 0.003714 | [0.935120, 0.939514]; 0.004394 | [0.940331, 0.943907]; 0.003571 | |
| 3 × 105 | 13 | [0.925153, 0.927434]; 0.002281 | [0.935323, 0.938044]; 0.002721 | [0.940505, 0.942882]; 0.002377 |
| 33 | [0.924483, 0.927434]; 0.002950 | [0.935323, 0.938272]; 0.002949 | [0.940505, 0.942919]; 0.002415 | |
| 100 | [0.924483, 0.927434]; 0.002950 | [0.934746, 0.938613]; 0.003867 | [0.939569, 0.943653]; 0.004084 | |
| 5 × 105 | 13 | [0.924771, 0.926431]; 0.001660 | [0.936047, 0.937644]; 0.001597 | [0.941309, 0.942485]; 0.001176 |
| 33 | [0.924771, 0.926558]; 0.001787 | [0.936047, 0.937886]; 0.001838 | [0.941043, 0.943080]; 0.002037 | |
| 100 | [0.924714, 0.926724; 0.002009 | [0.935679, 0.938194]; 0.002515 | [0.941012, 0.943089]; 0.002077 | |
| 1 × 106 | 13 | [0. 925045, 0.925313]; 0.000676 | [0.936453, 0.937261]; 0.000807 | [0.941301, 0.942233]; 0.000933 |
| 33 | [0. 925045, 0.926010]; 0.000965 | [0.936453, 0.937261]; 0.000808 | [0.941301, 0.942275]; 0.000974 | |
| 100 | [0.925045, 0.926048]; 0.001003 | [0.936341, 0.937473]; 0.001133 | [0.941218, 0.942491]; 0.001273 | |
| 1000 | [0.924874, 0.926079]; 0.001205 | [0.936297, 0.937635]; 0.001339 | [0.941195, 0.942521]; 0.001326 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Logofet, D.O.; Golubyatnikov, L.L.; Ulanova, N.G. Realistic Choice of Annual Matrices Contracts the Range of λS Estimates. Mathematics 2020, 8, 2252. https://doi.org/10.3390/math8122252
Logofet DO, Golubyatnikov LL, Ulanova NG. Realistic Choice of Annual Matrices Contracts the Range of λS Estimates. Mathematics. 2020; 8(12):2252. https://doi.org/10.3390/math8122252
Chicago/Turabian StyleLogofet, Dmitrii O., Leonid L. Golubyatnikov, and Nina G. Ulanova. 2020. "Realistic Choice of Annual Matrices Contracts the Range of λS Estimates" Mathematics 8, no. 12: 2252. https://doi.org/10.3390/math8122252