1. Introduction
The study of monitoring in population systems with tools of mathematical systems theory (MST) looks back to approximately three decades of history. A fundamental monograph on MST, dealing with basic concepts of controllability and observability, was authored by Kalman et al. [
1]. By population system, we mean either a) a single population structured in some way, or b) a set of interacting populations. To our knowledge, Varga [
2] was the first to apply the concept of observation systems of MST in order to recover the population genetic process from phenotypic observation. A recent continuation of this line of research was shown by Gámez et al. [
3]. Later on, this methodology was extended to the monitoring problem of a multispecies community, where, from the observation of certain species (indicators), we wanted to recover (estimate) the state process of the whole community.
For the monitoring of different ecological systems using the observer design methodology of Sundarapandian [
4], observer systems have previously been constructed, e.g., in Gámez et al. [
5,
6]. In particular, in work by López et al. [
7], the monitoring of Lotka–Volterra population systems was discussed. A review paper on the different applications of MST in population biology was authored by Varga [
8], and somewhat more recent surveys were published by Gámez [
9] and Varga et al. [
10]. In López et al.’s work [
11], by applying a technique of signed digraphs, a new, general approach to the design of a robust observer in continuous-time Lotka–Volterra models was proposed.
All the above references deal with classical continuous-time population models; however, for obvious reasons, in population biology, for practical purposes, discrete-time dynamics models should also be applied. First, in Gámez et al.’s work [
12], a discrete-time observer was constructed using the method of Sundarapandian [
13].
In the present paper, as a continuation of the above line of research, we adopt the latter observer design method to the monitoring of an age-structured population dynamics model, further developing and extending the initiative commenced by Gődényné Hajdu and Varga [
14].
In dynamic population models, both the state and the time may be either discrete or continuous. The classical matrix population models are discrete-state–discrete-time models, where, from the present state, the next state can be obtained by a linear transform.
The use of matrix models to describe the dynamics of an age-structured population goes back to Bernadelli [
15], who explained the periodic behavior observed in certain insect populations with a three-dimensional linear dynamic model. The classical discrete-time linear model of an age-specific population growth was introduced by Leslie [
16] and further developed by Leslie [
17].
We note that in matrix population models, both the state vector and the population projection matrix (PPM, Caswell [
18]) describing the linear state transform are non-negative; the dynamics model leaves the non-negative orthant invariant. This property is also characteristic of the linear dynamics models used in economics (see, e.g., Farina and Rinaldi [
19]). Therefore, for the study of the asymptotic behavior of such models (including the mathematical demography of human populations; see, e.g., Pianese et al. [
20]), the Perron– Frobenius theory of non-negative matrices was applied (see, e.g., Gantmacher [
21]), even in the discrete-state—continuous-time case (see, e.g., Varga [
22]). Stochastic variants of matrix population models (with environmental or life-cycle stochasticity), involving Markov chains, are discussed in Caswell’s work [
18], which is a comprehensive monograph on matrix population models.
The classical discrete-time linear Leslie model of an age-structured population growth, introduced by Leslie [
16], would often lead to an unlimited growth, and such behavior is obviously impossible in nature. There are different nonlinearities introduced in the model, implying a more realistic, limited growth. For example, in fisheries, to this end, a (nonlinear) recruitment function was introduced to describe the limited survival of newborn individuals. One of the most popular recruitment functions was due to Beverton and Holt [
23] (for further recruitment functions, see Getz and Haight [
24] and the recent review by Sharma et al. [
25]). We note that a variety of discrete-time density-dependent matrix population models can be obtained by substituting different state dependencies in the different entries of the PPM (see, e.g., Jensen [
26]). Further density-dependent matrix population models based on discrete logistic dynamics have been introduced by Liu and Cohen [
27]. For a recent overview of both density- and frequency-dependent matrix population models, we again refer to Caswell [
18].
In addition to age-specific population models, stage-specific models have also been developed. Since we would like to dedicate another study to the monitoring in stage-structured models (using observer design), in addition to the classical model of Lefkovitch [
28], here we only refer to the very recent surveys by Logofet and Salguero-Gómez [
29] and Logofet and Ulanova [
30] (see also Logofet and Klochkova [
31]). Stage-specific models, however, may display structural properties needing different constructions for observer design.
In this study, we consider a nonlinear Leslie-type model, where both the reproduction and the survival rates decrease as the total population size increases (see, e.g., Svirezhev and Logofet [
32] and Logofet [
33]). In this context, the monitoring problem means that from the observation of the size of certain age class(es) as a function of time, the whole state process (i.e., the time-dependent size of the rest of the age classes) should be recovered (estimated).
The motivation for the present study is the following: In the biological context, it often occurs that only the individuals of certain age class(es) of a population can be observed (i.e., counted). For example, if in
biological pest control in a greenhouse we wanted to estimate a pest population of insects, the imagos may be easier to count than other developmental stages (see, e.g., a collection of papers by Schreiber [
34] and a recent comprehensive review by Lima et al. [
35]). Another practical example occurs
in fisheries, where the fish population is estimated from the catch of only individuals over a threshold size. The
novelty of the present study is twofold: (a) Although Guiro et al. [
36] already applied an observer to estimate the fish stock in a stage-structured population model, their nonlinearity is again based on the above-mentioned Beverton–Holt recruitment function. Our monitoring approach is based, instead, on the nonlinear matrix model of Svirezhev and Logofet [
32] cited above, where all vital rates depend on the total population size. (b) In addition to the unknown state process, we also estimate an unknown environmental change affecting the vital rates of the population, which is a new feature of our work with respect to Guiro et al. [
36].
The paper is organized as follows: In
Section 2, a consistent nonlinear Leslie-type model is defined, with an invariant set as the phase space and conditions for the existence of a unique positive equilibrium of the dynamics.
Section 3 is dedicated to a stability analysis. In
Section 4, the observer design is illustrated. In
Section 5, it is shown how the observer design applies to estimate different (constant and periodic) unknown changes in the environment that affect the population dynamics. A Discussion and Outlook Section closes the main body of the paper.
2. Nonlinear Leslie-Type Model
In this section, we recall some basics of the classical discrete-time, age-structured matrix population model as a starting point of the nonlinear model, for which we propose a monitoring method based on the observer design of MST. Let
be a Leslie matrix with reproduction rates
survival rates
and state vector
Then, the classical Leslie model is
with a given non-negative initial state
(the set of all non-negative vectors of
).
In the following theorem, based on Svirezhev and Logofet [
32], in II. §4 and II. §5, we collected statements from the classical theory of Leslie models, which we needed in the sequel.
Theorem 1. Let the last age class be reproductive () and either let the first age class be reproductive () or have two consecutive reproductive age classes (for some). Then,has a unique positive and simple dominant eigenvalueand a unique associated eigenvectorwith. Furthermore, for any, we havefor some number.
In what follows, the conditions of Theorem 1 will be supposed for matrix .
Remark 1. The first condition,, impliesto be irreducible (Section II.4 inSvirezhev and Logofet [32]), while the second one is sufficient but not necessary forto be primitive (see Theorem A2.2 in Logofet [33]). It is known that
is a kind of long-term growth rate of the population: for
< 1, the population dies out (
), and for
> 1 the classical Leslie growth is of the Malthus type,
; in both cases, for
t >> 1 we have
. However, in order to avoid unlimited growth, the following modified, nonlinear model was introduced (see, e.g., Logofet [
33]).
2.1. Nonlinear Model
Obviously, in the modified, state-dependent Leslie matrix
, both the reproduction and the survival rates decrease as the total population size increases. The modified, nonlinear Leslie-type model then was
with
given.
2.2. Equilibrium of the Model
Let us find a nontrivial equilibrium of this dynamics, i.e., a nonzero fixed point of function
. First, we noted that, with a particular choice of
c, the
from Theorem 1 would be a fixed point of
. Indeed,
hence,
Here, of course, for should have held. In a more general setting, we had the following:
Theorem 2. Let the Theorem 1 conditions hold,
, andbe arbitrary. Then,is the unique positive equilibrium of dynamics (1).
Proof of Theorem 2. Substituting
, we have
Furthermore, let
be an equilibrium for dynamics (1), i.e.,
hence,
implying that
is a positive eigenvalue of
, associated with eigenvector
. Now, using Theorem 1,
is the unique positive eigenvalue of
, thus, we have
, and, hence,
. Furthermore, due to the simplicity of eigenvalue
, eigenvector
is a multiple of
. Hence, we obtain
, implying that
. □
2.3. Invariant Set
Now, we define a phase space (an invariant set) for dynamics (1).
Theorem 3. For any, setis invariant under.
Remark 2. Sinceis convex and compact, andis continuous, the Brouwer fixed-point theorem would imply thathas a fixed point in. Nevertheless,is always a trivial fixed point. Of course, we are interested in a nontrivial equilibrium of dynamics (1).
Now, in terms of the demographic parameters
and
, we find a necessary and sufficient condition for equilibrium
to belong to the invariant set
. It is known (Logofet [
33]) that the normed characteristic equation of
is
Hence, it is easy to see that a
is an eigenvalue of
if, and only if,
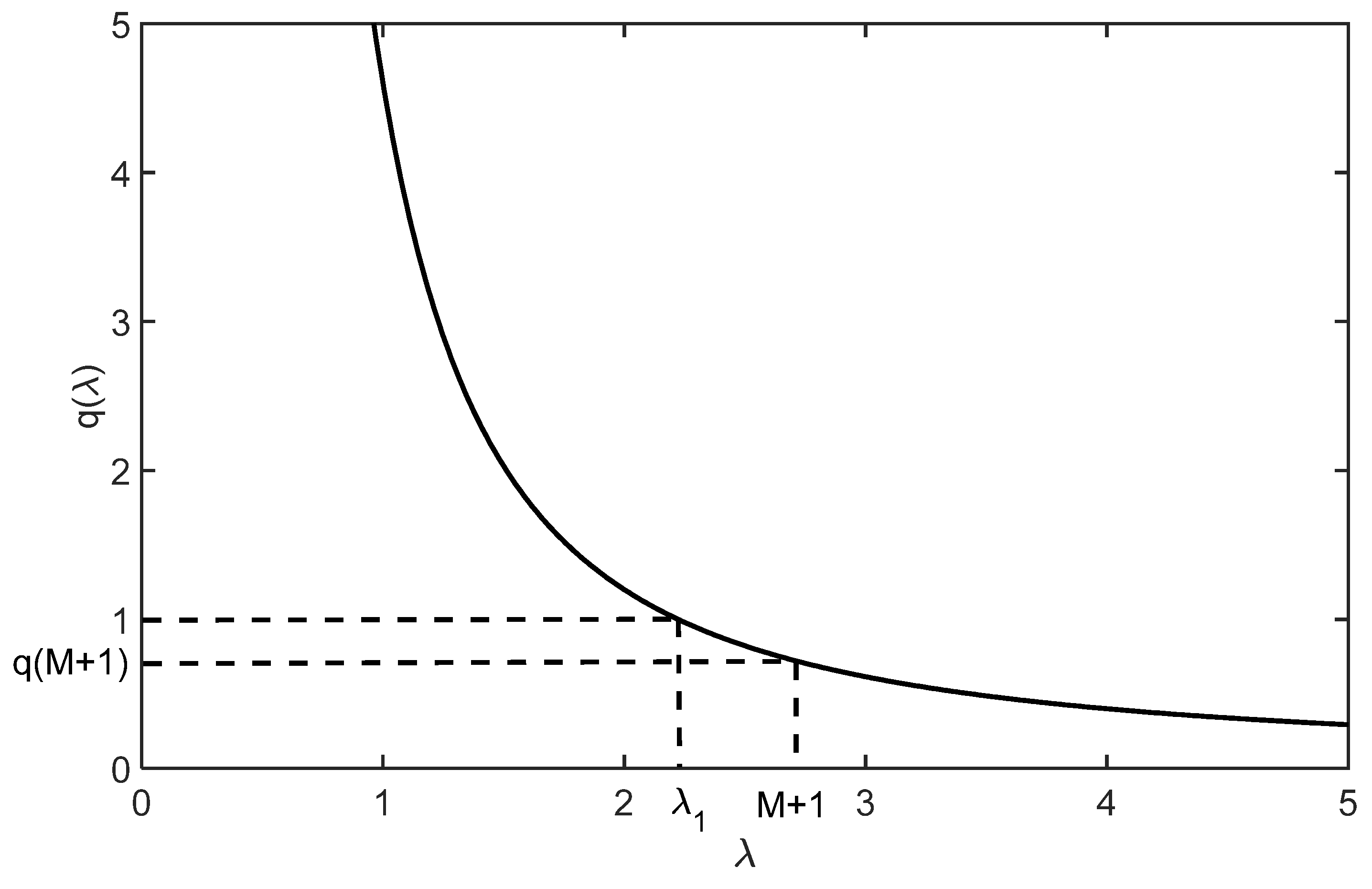
Furthermore, since function
is strictly decreasing, with
and
, condition
holds if, and only if,
(see
Figure 1).
Hence, we have the following
Theorem 4. Under the conditions of Theorem 1,if, and only if, for function q defined in (3), we have Example 1. Let us consider the simplified case, when all demographic parameters are the same:,. Then, Thus, inequality (5) was satisfied, implying that equilibriumbelongs to the interior of the phase space. This example also implies an infinite family of examples. Indeed, by the continuity of function, the above strict inequality also holds for all models having different demographic parametersand, sufficiently near the particular valuesandof the above example.
Remark 3. Since, in the above reasoning, inequality (5) is strictly satisfied, and, we can also conclude that in Example 1,also belongs to the interior of the invariant set. Through continuity reasoning, it also follows that for any parameter set forandclose to that of Example 1,also belongs to the interior of.
3. Stability of the Positive Equilibrium
For the sake of simplicity, we consider the case of
. Now, with a given
, dynamics (1) is considered with
In order to satisfy the conditions of Theorem 1, we suppose that
and at least one of the inequalities
or
, holds. Assuming that
, with Theorem 2 we obtain that
is the unique positive equilibrium of dynamics (1). Therefore, the Jacobian matrix at this positive equilibrium is
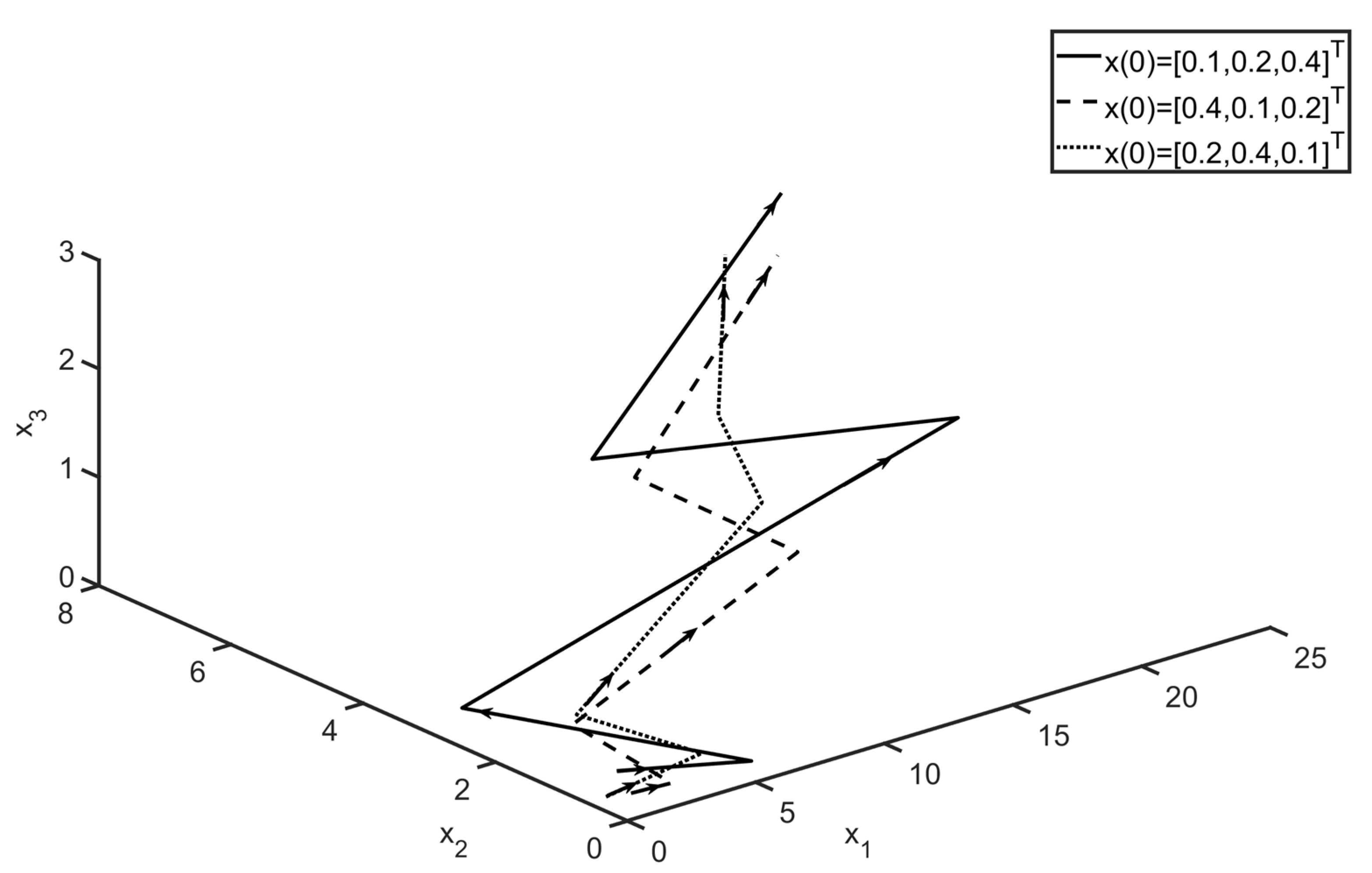
The following example illustrates the stability of the population age structure.
Example 2. Consider a population with the following demographic parameters:
, and set c = 0.05.
Conditions of Theorem 1 were fulfilled, so matrix has a unique positive and simple dominant eigenvalue and the unique normed eigenvector associated with it is (with ).
The unique positive equilibrium of dynamics (1) is
. Moreover, since
, we have
. Therefore,
. The eigenvalues of the Jacobian at equilibrium,
, are:
, 0.37. Therefore, A has a spectral radius of less than one (i.e., only has eigenvalues with modulus of less than one) and, hence, equilibrium
is asymptotically stable. In
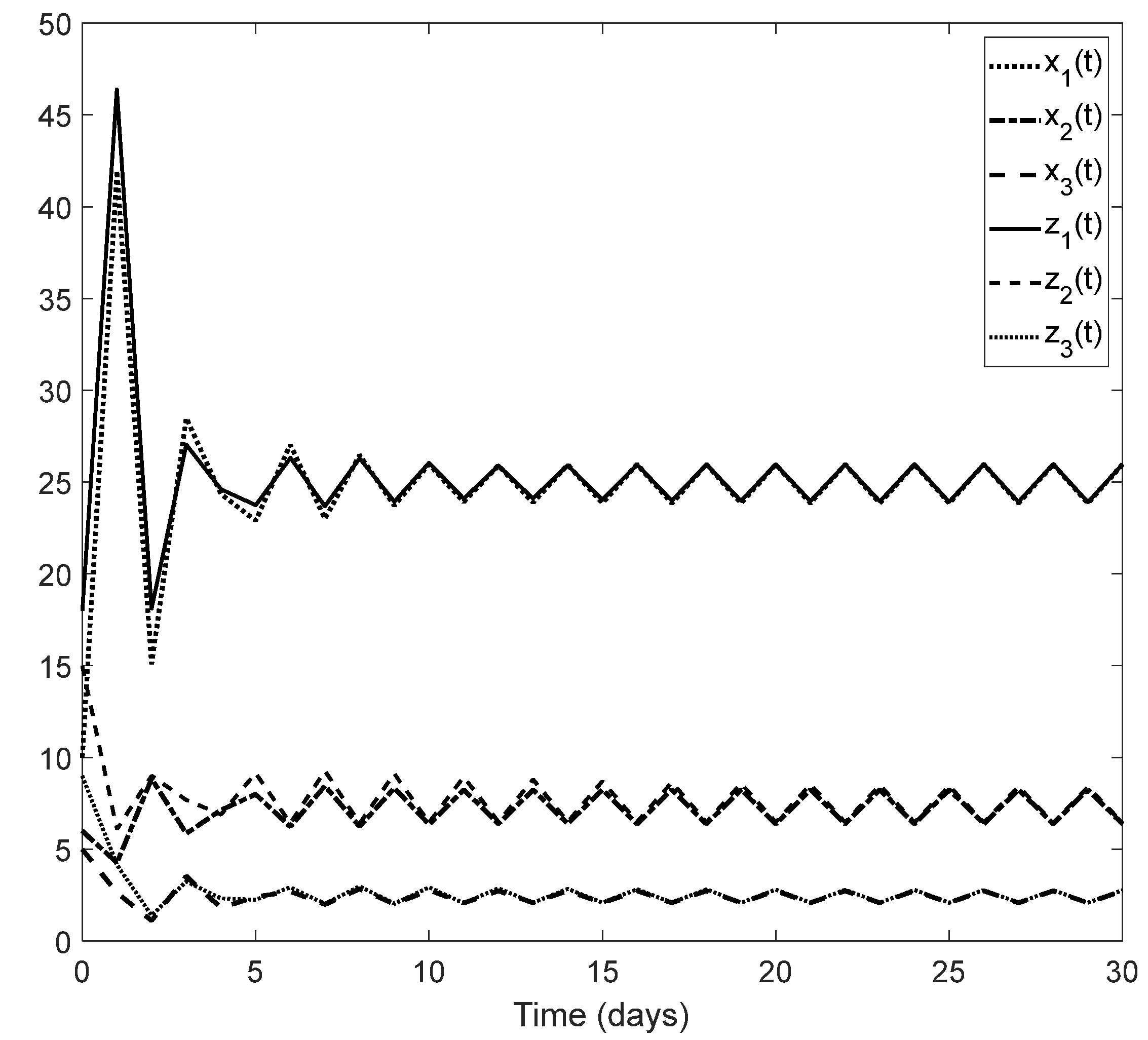
Figure 2, we can see how the solution of system (1) converge to
for different initial conditions.
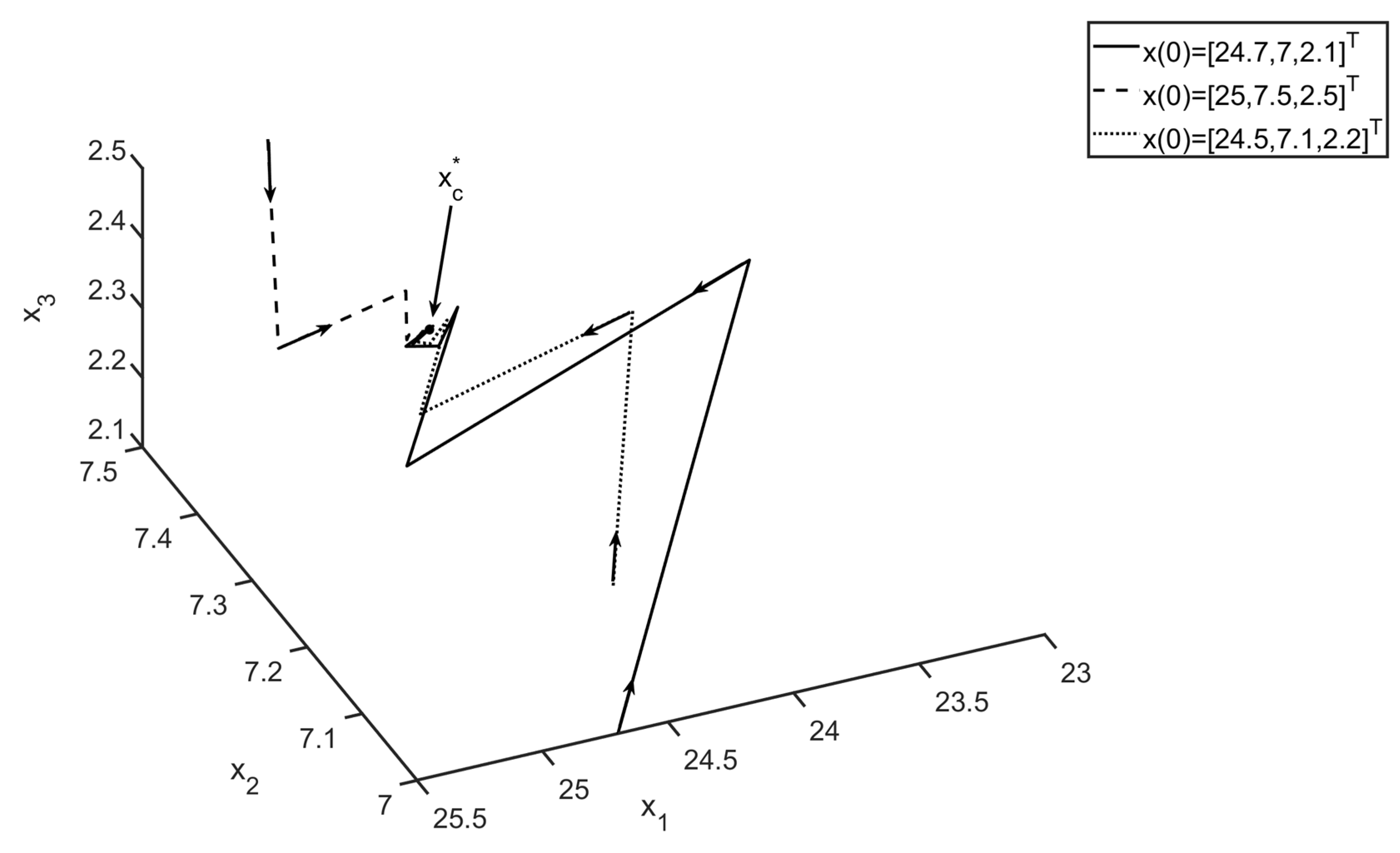
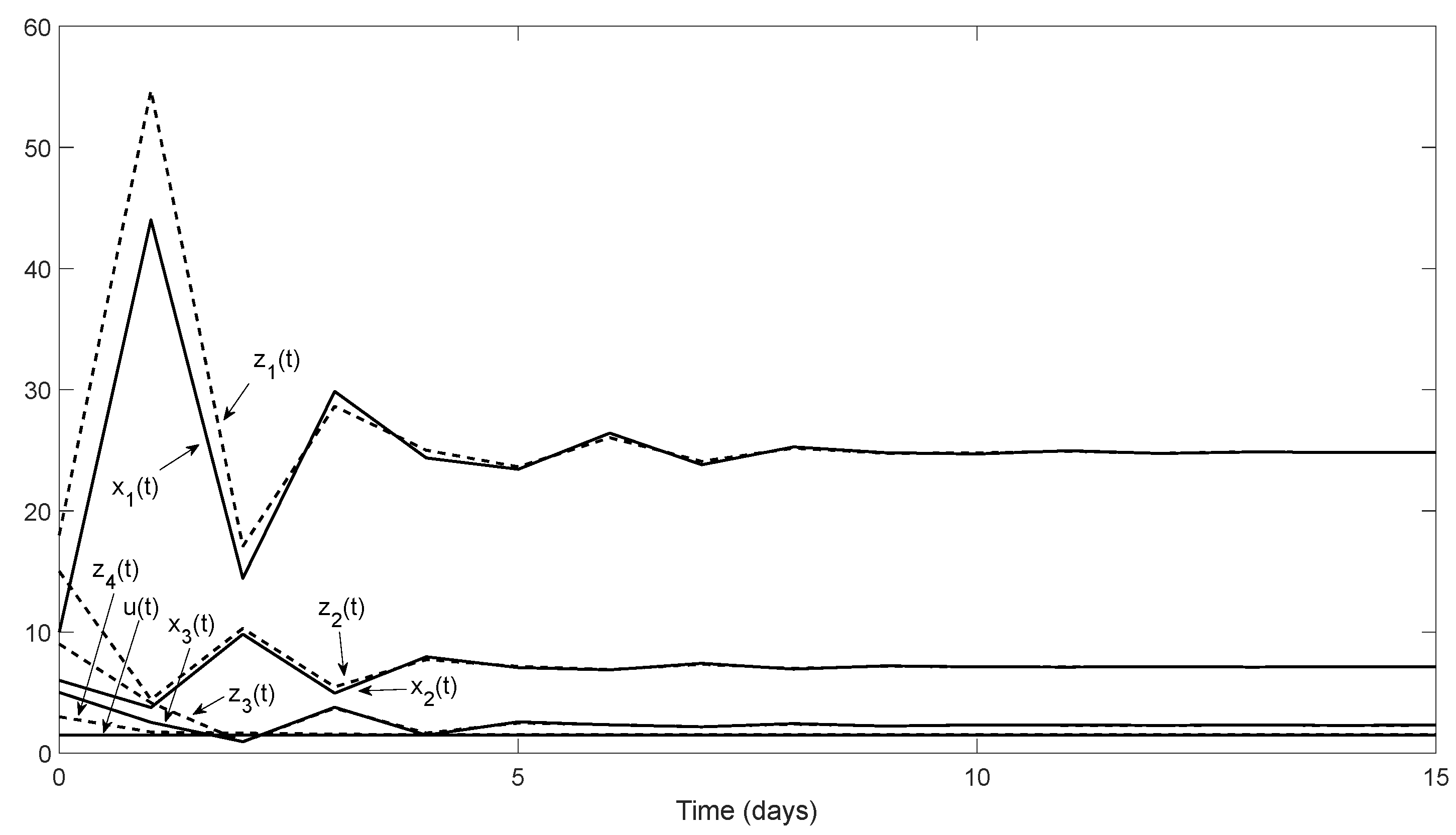
Remark 4. For the zero equilibrium, we have, and since, in our case,, the zero equilibrium is unstable. For our Example 1, inFigure 3, it is shown how the solutions of system (1) initially near zero diverge from zero (In fact, the eigenvalues ofare). 6. Discussion and Outlook
First of all, we note that Gámez et al. [
6] already proposed the observer approach to the deterministic stock estimation of a single fish population with reserve area, which obeyed the continuous-time logistic dynamics. The discrete-time observer approach to monitoring, proposed in the present paper (also including the case of a changing environment), is different from that considered by Ngom et al. [
38]. Indeed, in the latter, the first age class was governed by a Beverton–Holt recruitment function and the time-dependent survival rates (including a time-dependent fishing effort) were supposed, while there is a feedback of the total population size, both in the reproduction and the survival rates in our model.
Instead of an age-classified population, it is also natural to divide the individuals of the considered population into groups, according to their developmental stages. For such stage-specific population models applied in biological pest control, see, e.g., Garay et al. [
39]. Therefore, for both the theory and the possible applications, it is a further promising challenge to extend the monitoring methodology developed in the present paper, to
stage-structured population models. While in the age-specific model all surviving individuals pass to the next age class in unite time, a part of the surviving individuals may have remained in the same stage in the stage-specific model, resulting in a different dynamic model. Guiro et al. [
36] already applied an observer to estimate fish stock in a stage-structured population model, but the nonlinearity was again based on the Beverton–Holt recruitment function.
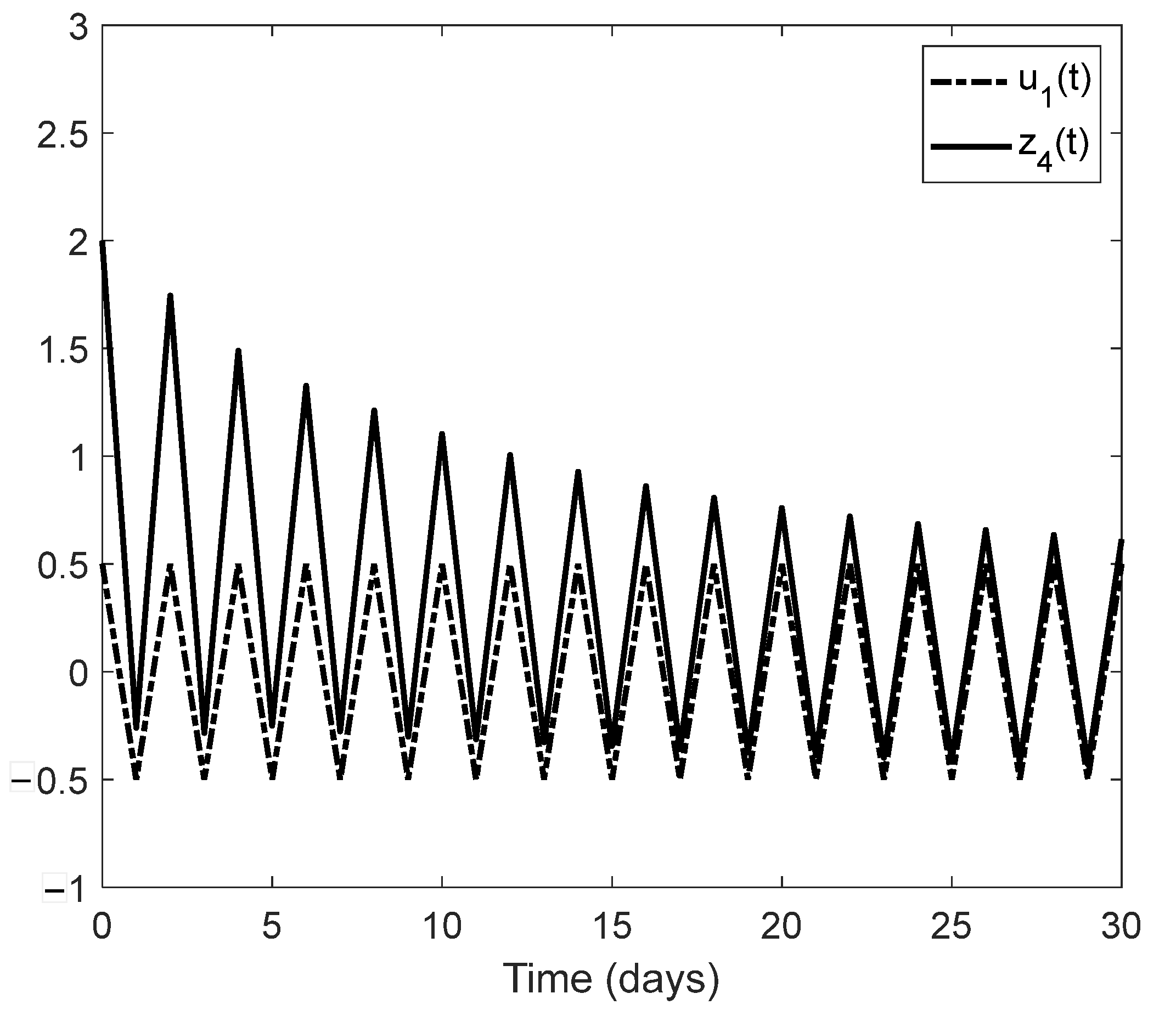
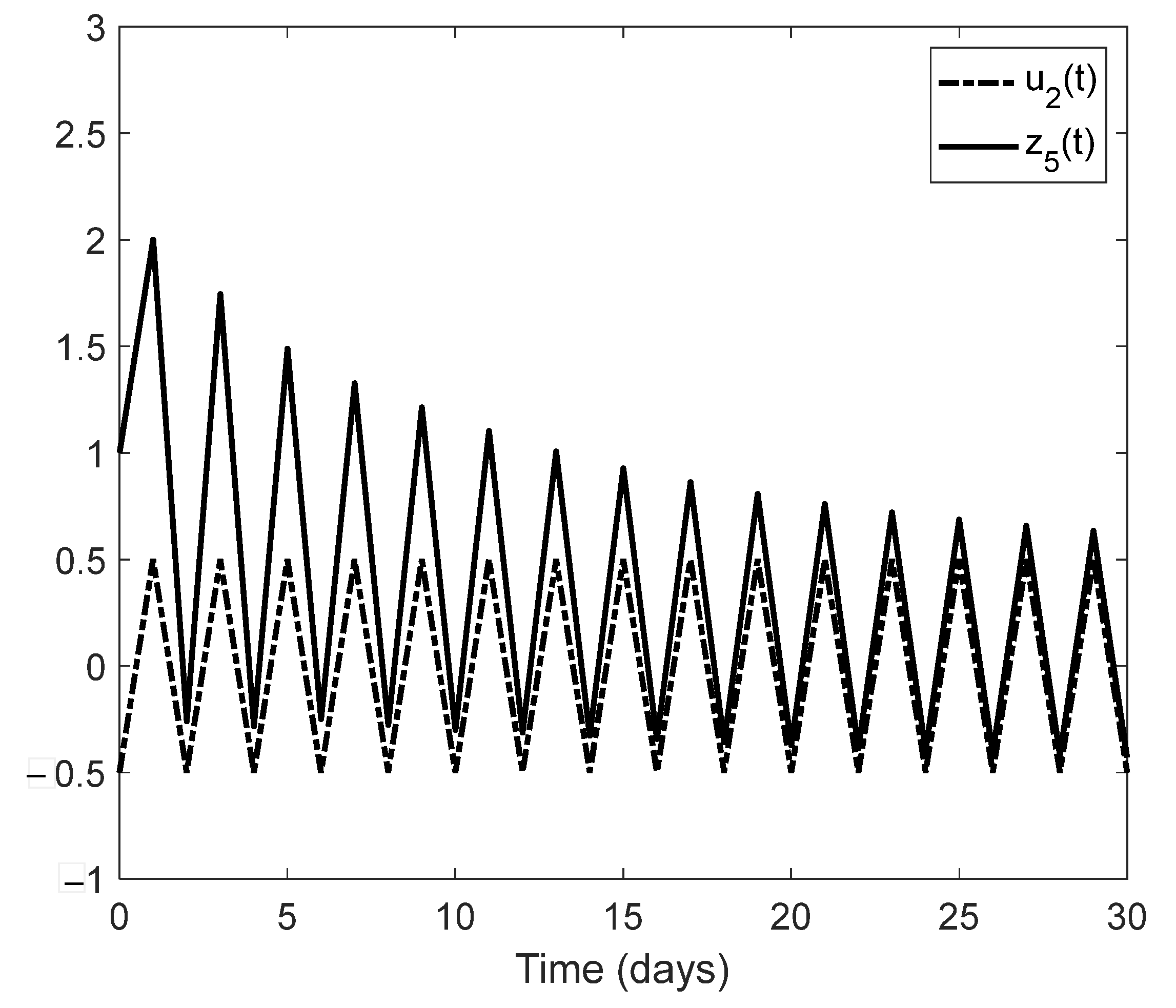
We showed examples of when an unknown change in the environment can be estimated together with the state process of an age-structured population. These examples could be starting points of a new line of research concerning a general monitoring system for environmental contamination based on the observation of easily observable age classes of a given indicator species. Another line of research that would go beyond the framework of the present paper might be an extension of the monitoring problem to the case of several interacting, age-structured populations. In this model, we would have to cope with two types of nonlinearity: one leading to the saturation according to the carrying capacity of the environment, and another one describing interspecific interactions such as predation.
Finally, for an outlook, we also mention that, while in the observer design, the system dynamics were known, and from the observation, an unknown state process was estimated. In the case of a structural identification problem, the structure of the system dynamics is known, and we want to recover its unknown parameters. On the structural identifiability of continuous-time nonlinear biological systems, a recent review was written by Villaverde [
40]. In the case of discrete-time systems, Anstett et al. [
41] may be a useful reference. Since model parameters can be considered as constant state variables, the structural identification problem can be considered as a particular case of the observation problem. In fact, in
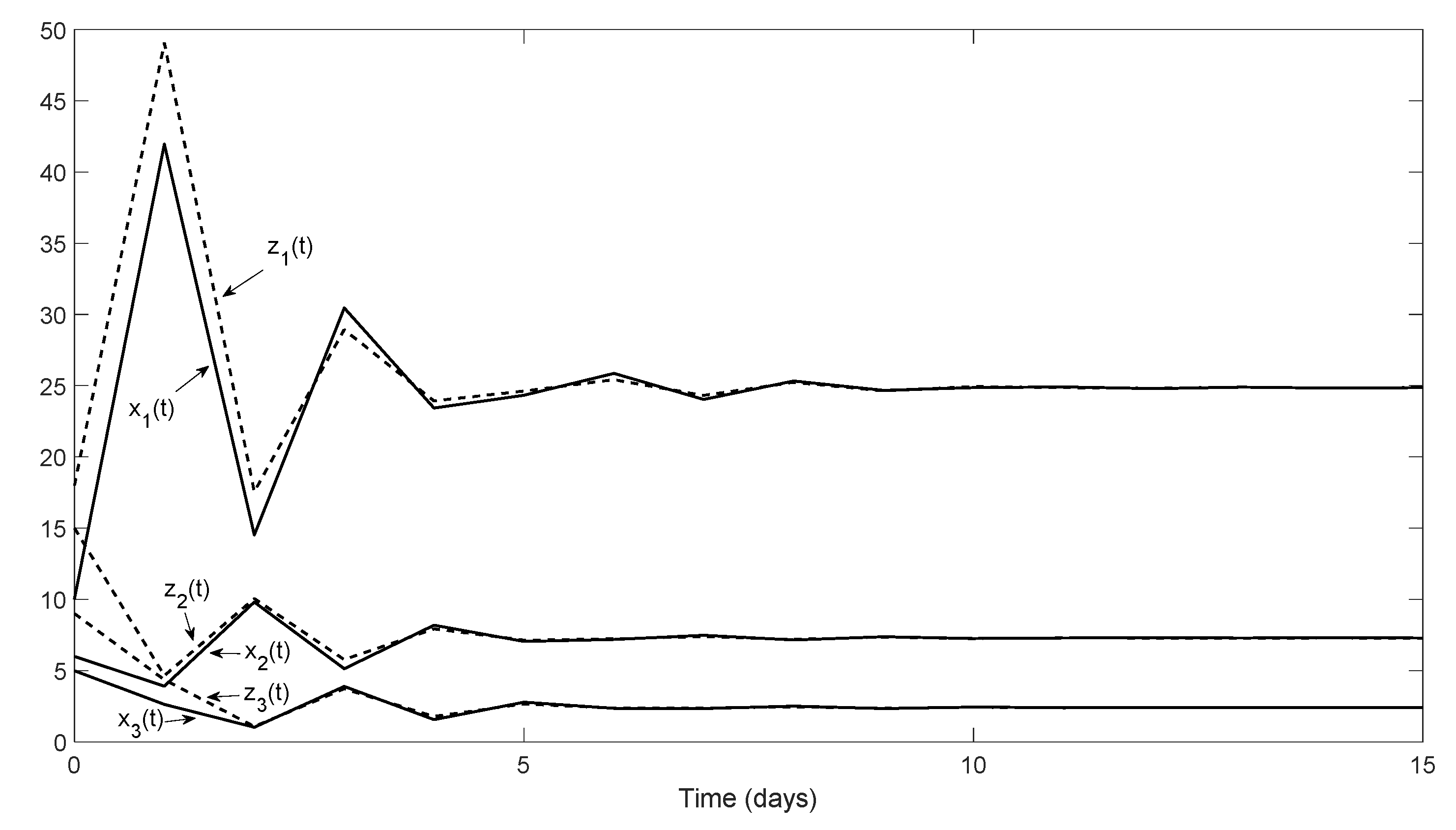
Section 5 of the present paper, observer design methodology was used to numerically estimate certain changes in population parameters due to abiotic environmental effects. This can be considered a particular case of structural identification. The generalization of this approach to different ecological situations, including several interacting species, could open a new line of research.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







