
Changes in the Saliva Proteome of Pigs with Diarrhoea Caused by Escherichia coli
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population of Animals
2.2. Saliva Collection and Sample Processing
2.3. SDS PAGE
2.4. Two-Dimensional (2-DE) Gel Electrophoresis
2.5. In-Gel Trypsin Digestion
2.6. Protein Identification through HPLC-MS/MS Analysis
2.7. Statistical Analysis
2.8. Validation
3. Results
3.1. Total Protein Concentration
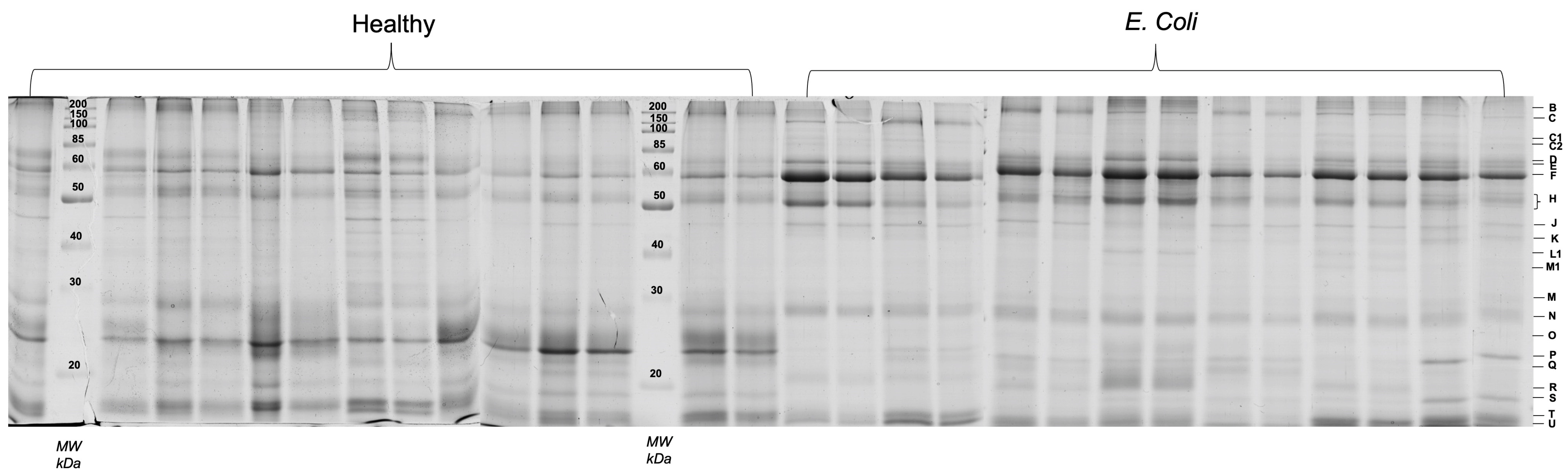
3.2. SDS-PAGE Profile
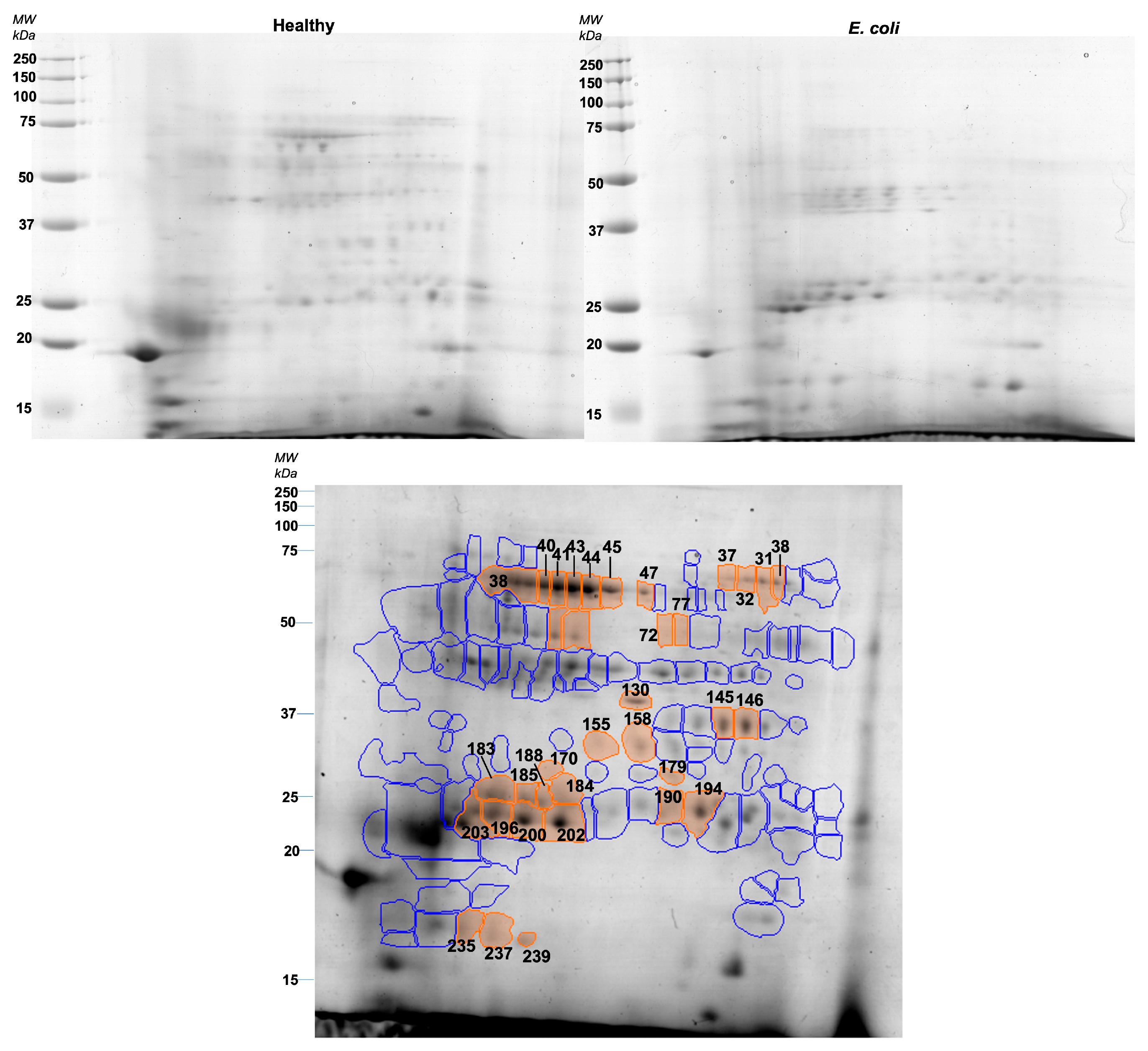
3.3. Two-Dimensional Protein Profile (2-DE)
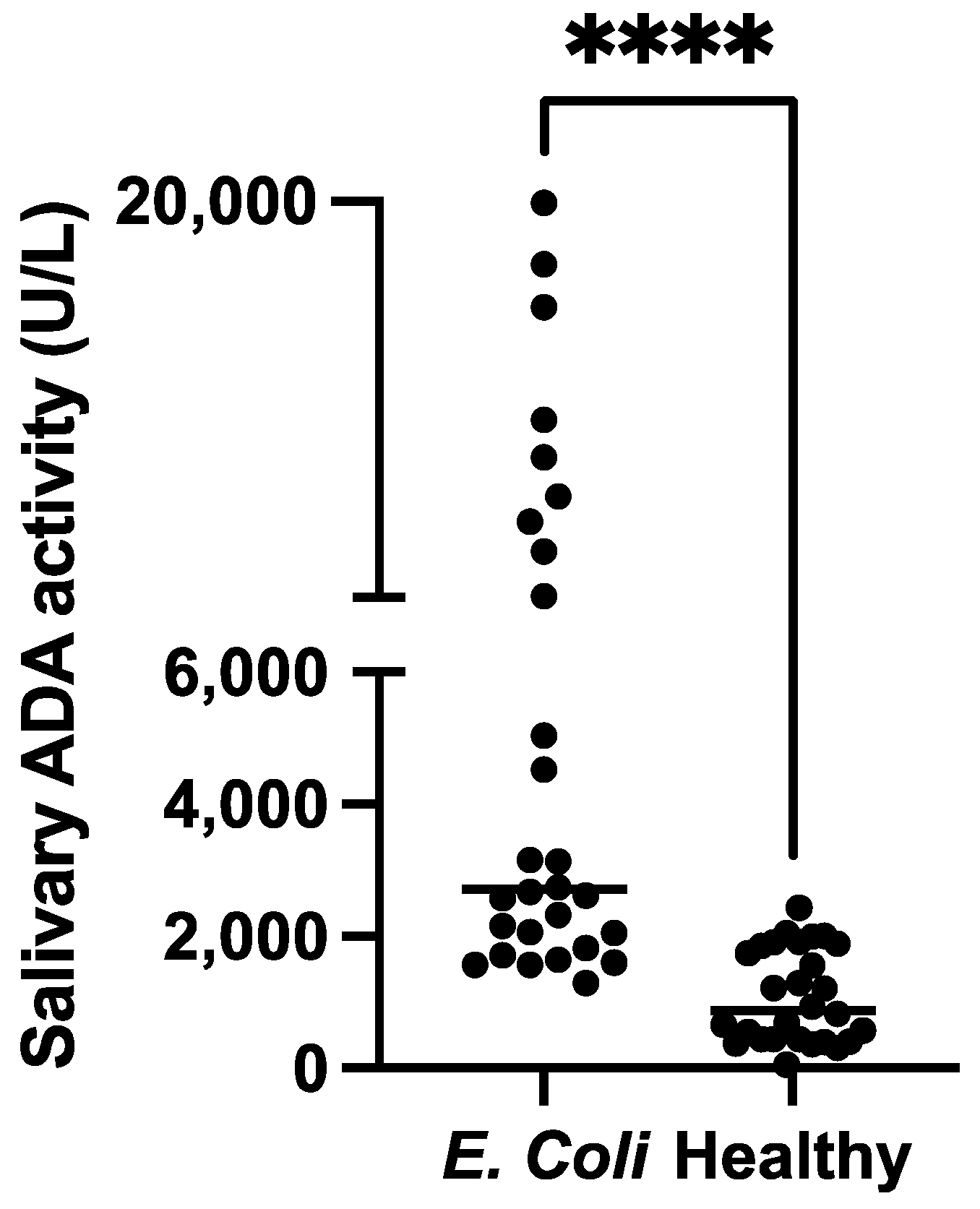
3.4. Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lamy, E.; Mau, M. Saliva proteomics as an emerging, non-invasive tool to study livestock physiology, nutrition and diseases. J. Proteom. 2012, 75, 4251–4258. [Google Scholar] [CrossRef] [PubMed]
- Cerón, J.J.; Contreras-Aguilar, M.D.; Escribano, D.; Martínez-Miró, S.; López-Martínez, M.J.; Ortín-Bustillo, A.; Franco-Martínez, L.; Rubio, C.P.; Muñoz-Prieto, A.; Tvarijonaviciute, A.; et al. Basics for the potential use of saliva to evaluate stress, inflammation, immune system, and redox homeostasis in pigs. BMC Vet. Res. 2022, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- López-Martínez, M.J.; Cerón, J.J.; Ortín-Bustillo, A.; Escribano, D.; Kuleš, J.; Beletić, A.; Rubić, I.; González-Sánchez, J.C.; Mrljak, V.; Martínez-Subiela, S.; et al. A Proteomic Approach to Elucidate the Changes in Saliva and Serum Proteins of Pigs with Septic and Non-Septic Inflammation. Int. J. Mol. Sci. 2022, 23, 6738. [Google Scholar] [CrossRef] [PubMed]
- López-Martínez, M.J.; Beletić, A.; Kuleš, J.; Rešetar-Maslov, D.; Rubić, I.; Mrljak, V.; Manzanilla, E.G.; Goyena, E.; Martínez-Subiela, S.; Cerón, J.J.; et al. Revealing the Changes in Saliva and Serum Proteins of Pigs with Meningitis Caused by Streptococcus Suis: A Proteomic Approach. Int. J. Mol. Sci. 2022, 23, 13700. [Google Scholar] [CrossRef] [PubMed]
- Escribano, D.; Horvatić, A.; Contreras-Aguilar, M.D.; Guillemin, N.; Cerón, J.J.; Tecles, F.; Martinez-Miró, S.; Eckersall, P.D.; Manteca, X.; Mrljak, V. Changes in saliva proteins in two conditions of compromised welfare in pigs: An experimental induced stress by nose snaring and lameness. Res. Vet. Sci. 2019, 125, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Yin, J.; Chen, S.; Duan, J.; Liu, G.; Li, T.; Li, N.; Peng, Y.; Tan, B.; Yin, Y. Proteome analysis for the global proteins in the jejunum tissues of enterotoxigenic Escherichia coli -infected piglets. Sci. Rep. 2016, 6, 25640. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Lv, Y.; Yang, J.; Bai, Y.; Ye, J.; Wang, Z.; Chen, Z.; Jiang, Z. Proteomic alteration of porcine intestinal epithelial cells after pretreatment with Lactobacillus plantarum followed by infection with enterotoxigenic Escherichia coli F4. Vet. Immunol. Immunopathol. 2020, 222, 109943. [Google Scholar] [CrossRef]
- Hartadi, E.B.; Helmi Effendi, M.; Plumeriastuti, H.; Sofiana, E.D.; Wibisono, F.M.; Hidayatullah, A.R.; Helmi, M. A Review of Enterotoxigenic Escherichia Coli Infection in Piglets: Public Health Importance. Syst. Rev. Pharm. 2020, 11, 687–698. [Google Scholar]
- Mesonero-Escuredo, S.; Strutzberg-Minder, K.; Casanovas, C.; Segalés, J. Viral and bacterial investigations on the aetiology of recurrent pig neonatal diarrhoea cases in Spain. Porc. Heal. Manag. 2018, 4, 5. [Google Scholar] [CrossRef]
- Lucena, S.; Coelho, A.V.; Capela-Silva, F.; Tvarijonaviciute, A.; Lamy, E. The Effect of Breed, Gender, and Acid Stimulation in Dog Saliva Proteome. BioMed Res. Int. 2018, 2018, 7456894. [Google Scholar] [CrossRef]
- Contreras-Aguilar, M.D.; Tvarijonaviciute, A.; Monkeviciene, I.; Martín-Cuervo, M.; González-Arostegui, L.G.; Franco-Martínez, L.; Cerón, J.J.; Tecles, F.; Escribano, D. Characterization of total adenosine deaminase activity (ADA) and its isoenzymes in saliva and serum in health and inflammatory conditions in four different species: An analytical and clinical validation pilot study. BMC Vet. Res. 2020, 16, 384. [Google Scholar] [CrossRef]
- Wang, L.; Xie, W.; Li, G.; Hu, B.; Wu, W.; Zhan, L.; Zou, H. Lipocalin 10 as a New Prognostic Biomarker in Sepsis-Induced Myocardial Dysfunction and Mortality: A Pilot Study. Mediat. Inflamm. 2021, 2021, 6616270. [Google Scholar] [CrossRef]
- Abella, V.; Scotece, M.; Conde, J.; Gómez, R.; Lois, A.; Pino, J.; Gómez-Reino, J.J.; Lago, F.; Mobasheri, A.; Gualillo, O. The potential of lipocalin-2/NGAL as biomarker for inflammatory and metabolic diseases. Biomarkers 2015, 20, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Stallhofer, J.; Friedrich, M.; Konrad-Zerna, A.; Wetzke, M.; Lohse, P.; Glas, J.; Tillack-Schreiber, C.; Schnitzler, F.; Beigel, F.; Brand, S. Lipocalin-2 Is a Disease Activity Marker in Inflammatory Bowel Disease Regulated by IL-17A, IL-22, and TNF-α and Modulated by IL23R Genotype Status. Inflamm. Bowel Dis. 2015, 21, 2327–2340. [Google Scholar] [CrossRef]
- Oikonomou, K.A.; Kapsoritakis, A.N.; Theodoridou, C.; Karangelis, D.; Germenis, A.; Stefanidis, I.; Potamianos, S.P. Neutrophil gelatinase-associated lipocalin (NGAL) in inflammatory bowel disease: Association with pathophysiology of inflammation, established markers, and disease activity. J. Gastroenterol. 2011, 47, 519–530. [Google Scholar] [CrossRef]
- Moschen, A.R.; Adolph, T.E.; Gerner, R.R.; Wieser, V.; Tilg, H. Lipocalin-2: A Master Mediator of Intestinal and Metabolic Inflammation. Trends Endocrinol. Metab. 2017, 28, 388–397. [Google Scholar] [CrossRef]
- Mitchell, G.B.; Clark, M.E.; Lu, R.; Caswell, J.L. Localization and Functional Characterization of Pulmonary Bovine Odorant-Binding Protein. Vet. Pathol. 2010, 48, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Yousuf, M.; Ali, A.; Khan, P.; Anjum, F.; Elasbali, A.M.; Islam, A.; Yadav, D.K.; Shafie, A.; Haque, Q.M.R.; Hassan, I. Insights into the Antibacterial Activity of Prolactin-Inducible Protein against the Standard and Environmental MDR Bacterial Strains. Microorganisms 2022, 10, 597. [Google Scholar] [CrossRef]
- Hassan, I.; Waheed, A.; Yadav, S.; Singh, T.P.; Ahmad, F. Prolactin inducible protein in cancer, fertility and immunoregulation: Structure, function and its clinical implications. Cell. Mol. Life Sci. 2008, 66, 447–459. [Google Scholar] [CrossRef]
- Kaiser, M.; Jacobsen, S.; Andersen, P.H.; Bækbo, P.; Cerón, J.J.; Dahl, J.; Escribano, D.; Theil, P.K.; Jacobson, M. Hormonal and metabolic indicators before and after farrowing in sows affected with postpartum dysgalactia syndrome. BMC Vet. Res. 2018, 14, 334. [Google Scholar] [CrossRef] [Green Version]
- Elmasry, R.; Negm, F.; Soliman, D.; Ahmed, E. Assessment of serum zinc, selenium, and prolactin concentrations in critically ill children. Pediatr. Health Med. Ther. 2016, 7, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tecles, F.; Rubio, C.P.; Contreras-Aguilar, M.D.; López-Arjona, M.; Miro, S.M.; Martinez-Subiela, S.; Cerón, J.J. Adenosine deaminase activity in pig saliva: Analytical validation of two spectrophotometric assays. J. Vet. Diagn. Investig. 2017, 30, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int. J. Mol. Sci. 2020, 21, 9254. [Google Scholar] [CrossRef] [PubMed]
- Fonti, R.; Latella, G.; Caprilli, R.; Frieri, G.; Marcheggiano, A.; Sambuy, M.Y.; Giovanni, R.F.; Renzo, L.; Giuseppe, C.; Adriana, F. Carbonic Anhydrase I Reduction in Colonic Mucosa of Patients with Active Ulcerative Colitis. Dig. Dis. Sci. 1998, 43, 2086–2092. [Google Scholar] [CrossRef]
- Contreras-Aguilar, M.D.; Escribano, D.; Martínez-Miró, S.; López-Arjona, M.; Rubio, C.P.; Martínez-Subiela, S.; Cerón, J.J.; Tecles, F. Application of a score for evaluation of pain, distress and discomfort in pigs with lameness and prolapses: Correlation with saliva biomarkers and severity of the disease. Res. Vet. Sci. 2019, 126, 155–163. [Google Scholar] [CrossRef]
- Contreras-Aguilar, M.D.; Escribano, D.; Martínez-Subiela, S.; Martínez-Miró, S.; Rubio, M.; Tvarijonaviciute, A.; Tecles, F.; Cerón, J.J. Influence of the way of reporting alpha-Amylase values in saliva in different naturalistic situations: A pilot study. PLoS ONE 2017, 12, e0180100. [Google Scholar] [CrossRef] [Green Version]
- Donadio, E.; Piccolomini, F.; Dimuccio, V.; Felicioli, A.; Balestreri, E.; Cianti, R.; Armini, A.; Bini, L.; Felicioli, R.; Donadio, C. Serum albumin fragmentation in end-stage renal disease patients—A pilot study. Clin. Chem. Lab. Med. 2009, 47, 1373–1379. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Band | Healthy | E. coli | p-Value | UNIPROT Protein Accession Number | Protein (Entry Name) | Seq Coverage (%) | ID Score | Theoretical MW (kDa) | Apparent MW (kDa) |
|---|---|---|---|---|---|---|---|---|---|
| B | 1.62 ± 0.80 | 5.36 ± 3.06 | 0.001 | 018758 | Submaxillary apomucin | 1.3 | 238.4 | 1184.1 | >200 kDa |
| C1 | - | ni | 120 | ||||||
| H | 3.74 ± 0.59 | 9.77 ± 2.91 | 0.0005 | A0A287B626 | IgA constant region | 39.3 | 209.6 | 44.2 | 54 |
| M # | 1.35 ± 1.09 | 2.94 ± 0.75 | 0.015 | A0A0A0MY58 and F1SN92 | Immunoglobulin heavy constant mu and Salivary lipocalin | 28.5 and 25.1 | 75.1 and 43.5 | 32.7 and 21.6 | 28.5 |
| N | 6.88 ± 2.44 | 10.20 ± 1.43 | 0.009 | F1SN92 | Salivary lipocalin | 54.9 | 152.5 | 21.6 | 26 |
| P | 17.51 ± 4.27 | 3.40 ± 2.10 | 0.0005 | P81245 | Odorant-binding protein | 75.1 | 199.5 | 17.7 | 18 |
| R | 1.22 ± 1.63 | 4.00 ± 2.47 | 0.033 | A0A4X1TU02 | Salivary lipocalin | 57.5 | 143.4 | 21.6 | 16.5 |
| T # | 14.15 ± 4.91 | 8.33 ± 4.70 | 0.043 | A0A286ZRW6 and A0A287ASS4 | Double-headed protease inhibitor, submandibular gland-like and Prolactin inducible protein | 29.4 and 36 | 58.31 and 56.35 | 13.3 and 12.4 | 13 |
| Spot Number | Fold Change | Group with Higher Level | p-Value | Protein (Entry Name) | UNIPROT Protein Accession Number | Seq Coverage (%) | ID Score | Theoretical MW (kDa) | Apparent MW (kDa) |
|---|---|---|---|---|---|---|---|---|---|
| 237 | 4.24 | E. coli | 5.24 × 10–5 | Adenosine deaminase and salivary lipocalin | A0A0B8RW47 and A0A4X1TU02 | 22.5 and 15.7 | 39.5 and 23.8 | 40.9 and 21.6 | 17.5 |
| 33 | 1.72 | Healthy control | 0.000222 | n.i. | |||||
| 185 | 2.30 | E. coli | 0.00063 | IgA constant region | A0A287B626 | 3.8 | 23.6 | 44.2 | 27.5 |
| 188 | 2.41 | E. coli | 0.000733 | IgA constant region | A0A287B626 | 2.6 | 20.0 | 44.2 | 27.5 |
| 41 | 3.29 | Healthy control | 0.000763 | Albumin (whole) | A0A286ZT13 | 41.1 | 327.8 | 68.2 | 74.5 |
| 145 | 1.56 | Healthy control | 0.000794 | Carbonate dehydratase VI | A0A4X1W7S7 | 15.1 | 39.5 | 34.7 | 36.0 |
| 40 | 2.72 | Healthy control | 0.000871 | Albumin (whole) | A0A286ZT13 | 41.1 | 327.8 | 68.2 | 74.5 |
| 202 | 2.28 | E. coli | 0.000887 | Ig-like domain-containing protein | A0A287A4Y3 | 15.4 | 41.4 | 24.7 | 26.0 |
| 44 | 3.03 | Healthy control | 0.001118 | Lactoperoxidase | A0A480RK36 | 6.6 | 48.7 | 80.3 | 74.5 |
| 196 | 2.97 | E. coli | 0.001675 | Albumin (fragment) and salivary lipocalin | A0A286ZT13 and A0A4X1TU02 | 13.6 and 23.5 | 100.5 and 31.5 | 68.2 and 21.6 | 26.0 |
| 200 | 2.34 | E. coli | 0.002233 | Albumin (fragment) and salivary lipocalin | A0A286ZT13 and A0A4X1TU02 | 13.6 and 23.5 | 100.5 and 31.5 | 68.2 and 21.6 | 26.0 |
| 45 | 2.10 | Healthy control | 0.002679 | Lactoperoxidase | A0A480RK36 | 6.7 | 48.8 | 80.3 | 74.5 |
| 43 | 3.30 | Healthy control | 0.003706 | Lactoperoxidase and polymeric immunoglobulin receptor | A0A480RK36 and A0A0E3M2Q4 | 7.5 and 6.5 | 45.5 and 37.3 | 80.3 and 67.3 | 74.5 |
| 194 | 1.71 | E. coli | 0.004477 | Albumin (fragment) | A0A286ZT13 | 7.8 | 64.3 | 68.2 | 26.5 |
| 31 | 2.29 | Healthy control | 0.005324 | n.i. | |||||
| 47 | 1.66 | Healthy control | 0.005441 | Lactoperoxidase and polymeric immunoglobulin receptor and | A0A0E3M2Q4 and A0A480RK36 | 12.3 and 4.9 | 86.1 and 34.7 | 67.3 and 80.3 | 74.0 |
| 184 | 1.86 | E. coli | 0.007066 | n.i. | |||||
| 203 | 2.94 | E. coli | 0.007897 | Ig-like domain-containing protein | A0A287A4Y3 | 18.5 | 34.0 | 24.7 | 26.0 |
| 38 | 2.12 | Healthy control | 0.009251 | Albumin (whole) | A0A286ZT13 | 41.1 | 327.8 | 68.2 | 101.0 |
| 32 | 1.50 | Healthy control | 0.012577 | n.i. | |||||
| 37 | 2.03 | Healthy control | 0.01381 | n.i. | |||||
| 155 | 1.51 | Healthy control | 0.015799 | Carbonic anhydrase | A0A4X1W9S1 | 11.0 | 27.7 | 36.3 | 36.0 |
| 179 | 1.96 | Healthy control | 0.020918 | Carbonate dehydratase VI | A0A4X1W7S7 | 11.5 | 47.2 | 34.7 | 27.5 |
| 73 | 1.78 | Healthy control | 0.021757 | Alpha-amylase | F1S573 | 30.1 | 146.0 | 55.8 | 58.0 |
| 74 | 1.39 | Healthy control | 0.026339 | Alpha-amylase | F1S573 | 30.9 | 123.4 | 55.8 | 58.0 |
| 235 | 2.49 | E. coli | 0.030702 | Adenosine deaminase and salivary lipocalin | A0A0B8RW47 and A0A4X1TU02 | 22.5 and 15.7 | 39.5 and 23.8 | 40.86 and 21.61 | 18.0 |
| 130 | 2.26 | E. coli | 0.033046 | n.i. | |||||
| 146 | 1.38 | Healthy control | 0.037883 | Carbonate dehydratase VI | A0A4X1W7S7 | 9.8 | 25.6 | 34.7 | 36.0 |
| 77 | 1.67 | Healthy control | 0.039092 | n.i. | |||||
| 190 | 1.75 | E. coli | 0.040238 | Albumin (fragment) | A0A286ZT13 | 9.2 | 61.0 | 68.2 | 26.0 |
| 239 | 2.94 | E. coli | 0.042094 | Salivary lipocalin | F1SN92 | 4.4 | 24.51 | 21.6 | 17.5 |
| 170 | 1.36 | E. coli | 0.046073 | n.i. | |||||
| 72 | 2.01 | Healthy control | 0.046326 | n.i. | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, M.; López-Martinez, M.J.; Ortin-Bustillo, A.; Cerón, J.J.; Martinez-Subiela, S.; Muñoz-Prieto, A.; Lamy, E. Changes in the Saliva Proteome of Pigs with Diarrhoea Caused by Escherichia coli. Proteomes 2023, 11, 14. https://doi.org/10.3390/proteomes11020014
Rodrigues M, López-Martinez MJ, Ortin-Bustillo A, Cerón JJ, Martinez-Subiela S, Muñoz-Prieto A, Lamy E. Changes in the Saliva Proteome of Pigs with Diarrhoea Caused by Escherichia coli. Proteomes. 2023; 11(2):14. https://doi.org/10.3390/proteomes11020014
Chicago/Turabian StyleRodrigues, Miguel, Maria José López-Martinez, Alba Ortin-Bustillo, Jose Joaquin Cerón, Silvia Martinez-Subiela, Alberto Muñoz-Prieto, and Elsa Lamy. 2023. "Changes in the Saliva Proteome of Pigs with Diarrhoea Caused by Escherichia coli" Proteomes 11, no. 2: 14. https://doi.org/10.3390/proteomes11020014