

Proteomics-Based Identification of Dysregulated Proteins and Biomarker Discovery in Invasive Ductal Carcinoma, the Most Common Breast Cancer Subtype

 , ,
, ,
Abstract
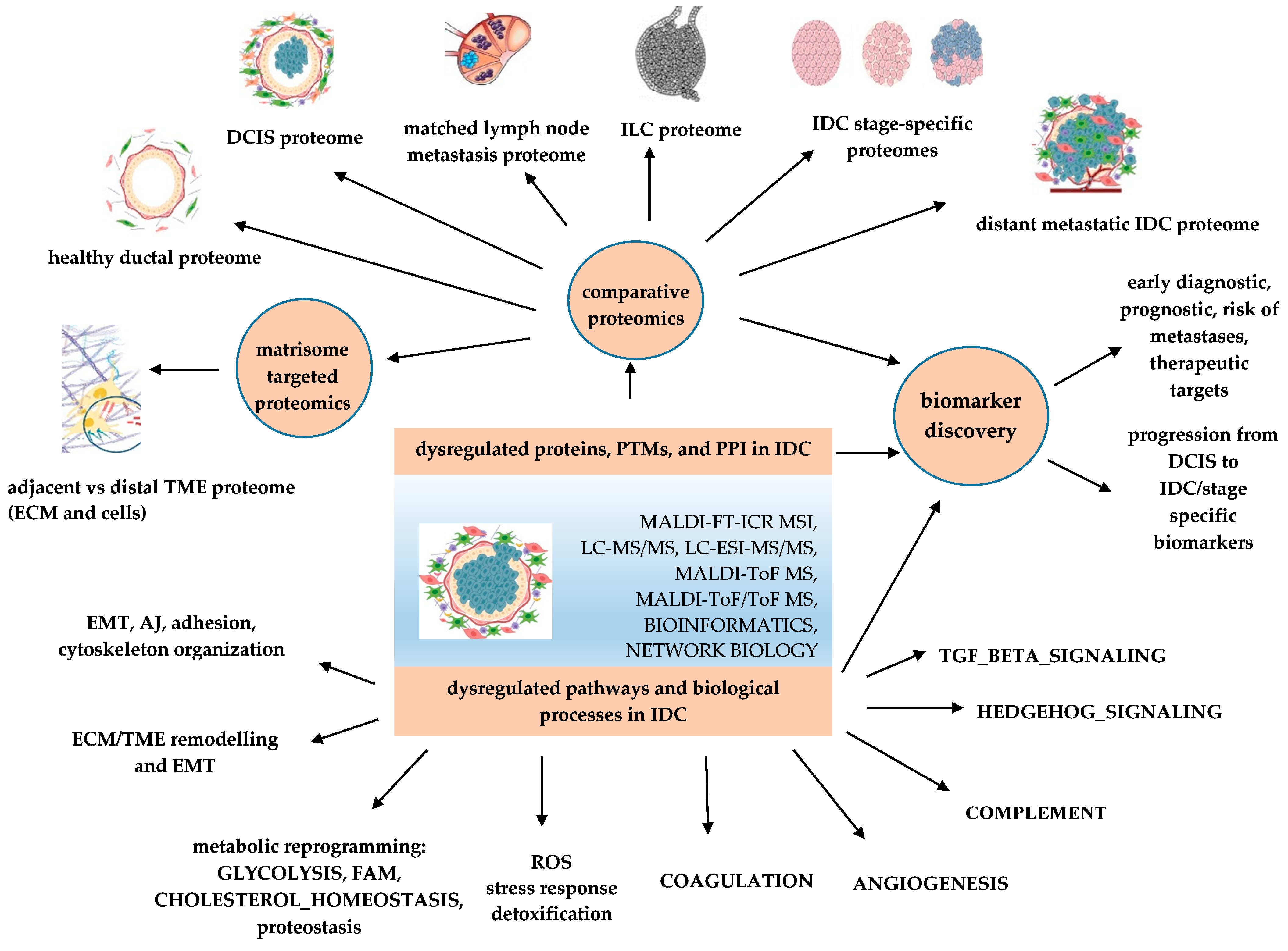
:1. Introduction
2. Differentiating IDC from Other BCs
3. Models of the Malignant Continuum from DCIS to IDC
4. Molecular Biomarker Discovery and Related Technological Advancements
5. Proteomics-Based Investigation of Dysregulated Proteins, Processes, and Pathways in IDC
5.1. Programs of EMT and EMT-Related Pathways Are Deeply Involved in IDC
5.2. Proteomic Remodeling of Tumor Microenvironment (TME) Is One of the Most Important Hallmarks of IDC

{kind=link}
| Dysregulated Proteins | Genes | Proteomics-Based Methods | Functions | Associated Roles in Cancer | References |
|---|---|---|---|---|---|
| Tenascin | TNC | LC–MS/MS | ECM protein | partial EMT marker [130]; cell adhesion, tissue remodeling, transduction of cellular signaling pathways [131] | [87] |
| Collagen isoforms | COL1A1, COL1A2, COL14A1 | LC–MS/MS; MALDI-FT-ICR MSI; HRAM, nanoLC-ESI–MS/MS | TME/ECM protein | cancer fibrosis, EMT [132,133] | [85,94] |
| Fibronectin | FN1 | LC–MS/MS | component of the mammary mesenchymal compartment of breast tumor | cell invasion, metastasis, tumor progression, EMT [134] | [85] |
| Periostin | POSTN/OSF-2 | FFPE, LCM, IHC, RT-PCR; LC–MS/MS | secreted ECM cell adhesion glycoprotein | EMT, proliferation, adhesion, migration [135] | [136] |
| Thrombospondins | THBS1/TSP1, THBS2/TSP2 | LC–MS/MS | ECM proteins | cell adhesion, invasion, migration, proliferation, apoptosis, tumor immunity [137] | [85,87] |
| Decorin | DCN | LC–MS/MS | small leucine-rich ECM proteoglycan | overexpression decreases migration, invasion, stemness and tumor growth and metastasis [138] | [85] |
| Lumican | LUM | LC–MSE, MALDI-MS/MS | small leucine-rich ECM proteoglycan | EMT regulator [139] | [23] |
| Mimecan/osteoglycin | OGN | LC–MS/MS | small leucine-rich ECM proteoglycan | inhibits BC cell proliferation and reverses EMT via repressing PI3K/AKT/mTOR pathway [140] | [85] |
| Matrix metalloproteinases | MMP-2, MMP-9 | 2-DE, MALDI-ToF MS | Zn-dependent endopeptidases | ECM remodeling, tumor initiation, progression, metastasis [141] | [88] |
5.3. Proteomics-Based Investigation of the Breast Cancer Proteomic Continuum Concept (BCPCC) in IDC for Non-Invasive Biomarker Discovery
5.4. Proteomics-Based Investigation of Protein Isoforms in IDC
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, H.; Langerød, A.; Ji, Y.; Nowels, K.W.; Nesland, J.M.; Tibshirani, R.; Bukholm, I.K.; Kåresen, R.; Botstein, D.; Børresen-Dale, A.-L.; et al. Different Gene Expression Patterns in Invasive Lobular and Ductal Carcinomas of the Breast. Mol. Biol. Cell 2004, 15, 2523–2536. [Google Scholar] [CrossRef] [Green Version]
- Kao, Y.; Wu, Y.-J.; Hsu, C.-C.; Lin, H.-J.; Wang, J.-J.; Tian, Y.-F.; Weng, S.-F.; Huang, C.-C. Short- and long-term recurrence of early-stage invasive ductal carcinoma in middle-aged and old women with different treatments. Sci. Rep. 2022, 12, 4422. [Google Scholar] [CrossRef]
- Fredolini, C.; Pathak, K.V.; Paris, L.; Chapple, K.M.; Tsantilas, K.A.; Rosenow, M.; Tegeler, T.J.; Garcia-Mansfield, K.; Tamburro, D.; Zhou, W.; et al. Shotgun proteomics coupled to nanoparticle-based biomarker enrichment reveals a novel panel of extracellular matrix proteins as candidate serum protein biomarkers for early-stage breast cancer detection. Breast Cancer Res. 2020, 22, 135. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Leone, J.P. Male Breast Cancer: An Updated Review of Epidemiology, Clinicopathology, and Treatment. J. Oncol. 2022, 2022, 1734049. [Google Scholar] [CrossRef] [PubMed]
- Makki, J. Diversity of Breast Carcinoma: Histological Subtypes and Clinical Relevance. Clinical medicine insights. Pathology 2015, 8, 23–31. [Google Scholar]
- Thennavan, A.; Beca, F.; Xia, Y.; Garcia-Recio, S.; Allison, K.; Collins, L.C.; Tse, G.M.; Chen, Y.-Y.; Schnitt, S.J.; Hoadley, K.A.; et al. Molecular analysis of TCGA breast cancer histologic types. Cell Genom. 2021, 1, 100067. [Google Scholar] [CrossRef]
- Zavyalova, M.V.; Denisov, E.V.; Tashireva, L.A.; Gerashchenko, T.S.; Litviakov, N.V.; Skryabin, N.A.; Vtorushin, S.V.; Telegina, N.S.; Slonimskaya, E.M.; Cherdyntseva, N.V.; et al. Phenotypic drift as a cause for intratumoral morphological heterogeneity of invasive ductal breast carcinoma not otherwise specified. BioRes. Open Access 2013, 2, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Rebbeck, C.A.; Xian, J.; Bornelöv, S.; Geradts, J.; Hobeika, A.; Geiger, H.; Alvarez, J.F.; Rozhkova, E.; Nicholls, A.; Robine, N.; et al. Gene expression signatures of individual ductal carcinoma in situ lesions identify processes and biomarkers associated with progression towards invasive ductal carcinoma. Nat. Commun. 2022, 13, 3399. [Google Scholar] [CrossRef]
- Dettogni, R.S.; Stur, E.; Laus, A.C.; Vieira, R.A.D.C.; Marques, M.M.C.; Santana, I.V.V.; Pulido, J.Z.; Ribeiro, L.F.; Parmanhani, N.D.J.; Agostini, L.P.; et al. Potential biomarkers of ductal carcinoma in situ progression. BMC Cancer 2020, 20, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Bai, F.; Wang, M.; Zhang, M.; Zhang, P.; Wu, K. The prognostic significance of co-existence ductal carcinoma in situ in invasive ductal breast cancer: A large population-based study and a matched case-control analysis. Ann. Transl. Med. 2019, 7, 484. [Google Scholar] [CrossRef] [PubMed]
- Mannu, G.S.; Wang, Z.; Broggio, J.; Charman, J.; Cheung, S.; Kearins, O.; Dodwell, D.; Darby, S.C. Invasive breast cancer and breast cancer mortality after ductal carcinoma in situ in women attending for breast screening in England, 1988–2014: Population based observational cohort study. BMJ 2020, 369, m1570. [Google Scholar] [CrossRef] [PubMed]
- van Seijen, M.; Lips, E.H.; Thompson, A.M.; Nik-Zainal, S.; Futreal, A.; Hwang, E.S.; Verschuur, E.; Lane, J.; Jonkers, J.; Rea, D.W.; et al. Ductal carcinoma in situ: To treat or not to treat, that is the question. Br. J. Cancer 2019, 121, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meurs, C.J.C.; Van Rosmalen, J.; Menke-Pluijmers, M.B.E.; Ter Braak, B.P.M.; De Munck, L.; Siesling, S.; Westenend, P.J. A prediction model for underestimation of invasive breast cancer after a biopsy diagnosis of ductal carcinoma in situ: Based on 2892 biopsies and 589 invasive cancers. Br. J. Cancer 2018, 119, 1155–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thennavan, A.; Garcia-Recio, S.; Liu, S.; He, X.; Perou, C.M. Molecular signatures of in situ to invasive progression for basal-like breast cancers: An integrated mouse model and human DCIS study. NPJ Breast Cancer 2022, 8, 83. [Google Scholar] [CrossRef]
- Brock, E.J.; Ji, K.; Shah, S.; Mattingly, R.R.; Sloane, B.F. In Vitro Models for Studying Invasive Transitions of Ductal Carcinoma In Situ. J. Mammary Gland. Biol. Neoplasia 2019, 24, 1–15. [Google Scholar] [CrossRef]
- Heller, S.L.; Plaunova, A.; Gao, Y. Ductal Carcinoma In Situ and Progression to Invasive Cancer: A Review of the Evidence. J. Breast Imaging 2021, 3, 135–143. [Google Scholar] [CrossRef]
- Casasent, A.K.; Edgerton, M.; Navin, N.E. Genome evolution in ductal carcinoma in situ: Invasion of the clones. J. Pathol. 2017, 241, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Cowell, C.F.; Weigelt, B.; Sakr, R.A.; Ng, C.K.; Hicks, J.; King, T.A.; Reis-Filho, J.S. Progression from ductal carcinoma in situ to invasive breast cancer: Revisited. Mol. Oncol. 2013, 7, 859–869. [Google Scholar] [CrossRef]
- Lesurf, R.; Aure, M.R.; Mørk, H.H.; Vitelli, V.; Lundgren, S.; Børresen-Dale, A.-L.; Kristensen, V.; Wärnberg, F.; Hallett, M.; Sørlie, T.; et al. Molecular Features of Subtype-Specific Progression from Ductal Carcinoma In Situ to Invasive Breast Cancer. Cell Rep. 2016, 16, 1166–1179. [Google Scholar] [CrossRef] [Green Version]
- Trinh, A.; Gil Del Alcazar, C.R.; Shukla, S.A.; Chin, K.; Chang, Y.H.; Thibault, G.; Eng, J.; Jovanović, B.; Aldaz, C.M.; Park, S.Y.; et al. Genomic Alterations during the In Situ to Invasive Ductal Breast Carcinoma Transition Shaped by the Immune System. Mol. Cancer Res. MCR 2021, 19, 623–635. [Google Scholar] [CrossRef]
- Heselmeyer-Haddad, K.; Garcia, L.Y.B.; Bradley, A.; Ortiz-Melendez, C.; Lee, W.-J.; Christensen, R.; Prindiville, S.A.; Calzone, K.A.; Soballe, P.W.; Hu, Y.; et al. Single-Cell Genetic Analysis of Ductal Carcinoma in Situ and Invasive Breast Cancer Reveals Enormous Tumor Heterogeneity yet Conserved Genomic Imbalances and Gain of MYC during Progression. Am. J. Pathol. 2012, 181, 1807–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.C.; Koestler, D.C.; Fleischer, T.; Chen, P.; Jenson, E.G.; Marotti, J.D.; Onega, T.; Kristensen, V.N.; Christensen, B.C. DNA methylation in ductal carcinoma in situ related with future development of invasive breast cancer. Clin. Epigenet. 2015, 7, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korwar, A.M.; Bhonsle, H.S.; Ghole, V.S.; Gawai, K.R.; Koppikar, C.B.; Kulkarni, M.J. Proteomic Profiling and Interactome Analysis of ER-Positive/HER2/ neu Negative Invasive Ductal Carcinoma of the Breast: Towards Proteomics Biomarkers. Omics J. Integr. Biol. 2013, 17, 27–40. [Google Scholar] [CrossRef] [PubMed]
- More, T.H.; RoyChoudhury, S.; Christie, J.; Taunk, K.; Mane, A.; Santra, M.K.; Chaudhury, K.; Rapole, S. Metabolomic alterations in invasive ductal carcinoma of breast: A comprehensive metabolomic study using tissue and serum samples. Oncotarget 2017, 9, 2678–2696. [Google Scholar] [CrossRef] [Green Version]
- Saha, T.; Solomon, J.; Samson, A.; Gil-Henn, H. Invasion and Metastasis as a Central Hallmark of Breast Cancer. J. Clin. Med. 2021, 10, 3498. [Google Scholar] [CrossRef] [PubMed]
- Macklin, A.; Khan, S.; Kislinger, T. Recent advances in mass spectrometry based clinical proteomics: Applications to cancer research. Clin. Proteom. 2020, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Xu, F.; Fang, D.; Chen, Y. Targeted Proteomics Enables Simultaneous Quantification of Folate Receptor Isoforms and Potential Isoform-based Diagnosis in Breast Cancer. Sci. Rep. 2015, 5, 16733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beňačka, R.; Szabóová, D.; Guľašová, Z.; Hertelyová, Z.; Radoňák, J. Classic and New Markers in Diagnostics and Classification of Breast Cancer. Cancers 2022, 14, 5444. [Google Scholar] [CrossRef]
- Barron, M.; Asaad, A.; Idaewor, P.; Rasheed, N.; Al-Zawi, A.S.A. Breast Apocrine Carcinoma Detected Incidentally as Axillary Lymphadenopathy in a CT Scan. Cureus 2021, 13, e18523. [Google Scholar] [CrossRef]
- Lee, M.; Jara-Lazaro, A.R.; Cheok, P.Y.; Thike, A.A. Medullary breast carcinoma: A pathogenic review and immunohistochemical study using tissue microarray. Singap. Med. J. 2022, 63, 394–401. [Google Scholar]
- Yim, H.E.; Kim, J.-H.; Ahn, M.S.; Jung, Y.; Roh, J.; Park, S.H.; Kim, T.-G.; Choi, J.-H.; Kang, S.Y. Clinicopathological and Molecular Analysis of 45 Cases of Pure Mucinous Breast Cancer. Front. Oncol. 2021, 10, 558760. [Google Scholar] [CrossRef]
- Nascimento, R.; Otoni, K. Histological and molecular classification of breast cancer: What do we know? Mastology 2020, 30. [Google Scholar] [CrossRef]
- Romaniuk, A.; Lyndin, M.; Sikora, V.; Lyndina, Y.; Panasovska, K. Histological and immunohistochemical features of medullary breast cancer. Folia Med. Crac. 2015, 2, 41–48. [Google Scholar]
- Wilson, G.M.; Dinh, P.; Pathmanathan, N.; Graham, J.D. Ductal Carcinoma in Situ: Molecular Changes Accompanying Disease Progression. J. Mammary Gland. Biol. Neoplasia 2022, 27, 101–131. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Céspedes, R.; Suárez-Bonnet, A.; Millán, Y.; Guil-Luna, S.; Reymundo, C.; Herráez, P.; Monteros, A.E.D.L.; Mulas, J.M.D.L. Use of CD10 as a marker of canine mammary myoepithelial cells. Vet. J. 2013, 195, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Russell, T.D.; Jindal, S.; Agunbiade, S.; Gao, D.; Troxell, M.; Borges, V.F.; Schedin, P. Myoepithelial Cell Differentiation Markers in Ductal Carcinoma in Situ Progression. Am. J. Pathol. 2015, 185, 3076–3089. [Google Scholar] [CrossRef] [Green Version]
- Moelans, C.B.; de Wegers, R.A.; Monsuurs, H.N.; Maess, A.H.J.; van Diest, P.J. Molecular differences between ductal carcinoma in situ and adjacent invasive breast carcinoma: A multiplex ligation-dependent probe amplification study. Cell. Oncol. 2011, 34, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Jung, S.H.; Kim, M.S.; Baek, I.-P.; Lee, S.H.; Kim, T.-M.; Chung, Y.-J.; Lee, S.H. Genomic differences between pure ductal carcinoma in situ and synchronous ductal carcinoma in situ with invasive breast cancer. Oncotarget 2015, 6, 7597–7607. [Google Scholar] [CrossRef] [Green Version]
- Arps, D.P.; Healy, P.; Zhao, L.; Kleer, C.G.; Pang, J.C. Invasive ductal carcinoma with lobular features: A comparison study to invasive ductal and invasive lobular carcinomas of the breast. Breast Cancer Res. Treat. 2013, 138, 719–726. [Google Scholar] [CrossRef] [Green Version]
- Barroso-Sousa, R.; Metzger-Filho, O. Differences between invasive lobular and invasive ductal carcinoma of the breast: Results and therapeutic implications. Ther. Adv. Med. Oncol. 2016, 8, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Adachi, Y.; Ishiguro, J.; Kotani, H.; Hisada, T.; Ichikawa, M.; Gondo, N.; Yoshimura, A.; Kondo, N.; Hattori, M.; Sawaki, M.; et al. Comparison of clinical outcomes between luminal invasive ductal carcinoma and luminal invasive lobular carcinoma. BMC Cancer 2016, 16, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shee, K.; Muller, K.E.; Marotti, J.; Miller, T.W.; Wells, W.A.; Tsongalis, G.J. Ductal Carcinoma in Situ Biomarkers in a Precision Medicine Era: Current and Future Molecular-Based Testing. Am. J. Pathol. 2019, 189, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Casasent, A.K.; Schalck, A.; Gao, R.; Sei, E.; Long, A.; Pangburn, W.; Casasent, T.; Meric-Bernstam, F.; Edgerton, M.E.; Navin, N.E. Multiclonal Invasion in Breast Tumors Identified by Topographic Single Cell Sequencing. Cell 2018, 172, 205–217.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pareja, F.; Brown, D.N.; Lee, J.Y.; Paula, A.D.C.; Selenica, P.; Bi, R.; Geyer, F.C.; Gazzo, A.; da Silva, E.M.; Vahdatinia, M.; et al. Whole-Exome Sequencing Analysis of the Progression from Non-Low-Grade Ductal Carcinoma In Situ to Invasive Ductal Carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 3682–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, H.; Breitling, R.; Francavilla, C.; Schwartz, J.-M. Data integration and mechanistic modelling for breast cancer biology: Current state and future directions. Curr. Opin. Endocr. Metab. Res. 2022, 24, 100350. [Google Scholar] [CrossRef]
- Schultz, S.; Bartsch, H.; Sotlar, K.; Petat-Dutter, K.; Bonin, M.; Kahlert, S.; Harbeck, N.; Vogel, U.; Seeger, H.; Fehm, T.; et al. Progression-specific genes identified in microdissected formalin-fixed and paraffin-embedded tissue containing matched ductal carcinoma in situ and invasive ductal breast cancers. BMC Med. Genom. 2018, 11, 80. [Google Scholar] [CrossRef]
- Whitworth, P.W.; Beitsch, P.D.; Murray, M.K.; Richards, P.D.; Mislowsky, A.; Dul, C.L.; Pellicane, J.V.; Baron, P.L.; Rahman, R.L.; Lee, L.A.; et al. Genomic Classification of HER2-Positive Patients With 80-Gene and 70-Gene Signatures Identifies Diversity in Clinical Outcomes With HER2-Targeted Neoadjuvant Therapy. JCO Precis. Oncol. 2022, 6, e2200197. [Google Scholar] [CrossRef]
- Asleh, K.; Negri, G.L.; Miko, S.E.S.; Colborne, S.; Hughes, C.S.; Wang, X.Q.; Gao, D.; Gilks, C.B.; Chia, S.K.L.; Nielsen, T.O.; et al. Proteomic analysis of archival breast cancer clinical specimens identifies biological subtypes with distinct survival outcomes. Nat. Commun. 2022, 13, 896. [Google Scholar] [CrossRef]
- Bouchal, P.; Schubert, O.T.; Faktor, J.; Capkova, L.; Imrichova, H.; Zoufalova, K.; Paralova, V.; Hrstka, R.; Liu, Y.; Ebhardt, H.A.; et al. Breast Cancer Classification Based on Proteotypes Obtained by SWATH Mass Spectrometry. Cell Rep. 2019, 28, 832–843.e7. [Google Scholar] [CrossRef]
- Ntai, I.; LeDuc, R.D.; Fellers, R.T.; Erdmann-Gilmore, P.; Davies, S.R.; Rumsey, J.; Early, B.P.; Thomas, P.M.; Li, S.; Compton, P.D.; et al. Integrated Bottom-Up and Top-Down Proteomics of Patient-Derived Breast Tumor Xenografts*. Mol. Cell. Proteom. 2016, 15, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Cristobal, A.; Marino, F.; Post, H.; Toorn, H.W.P.V.D.; Mohammed, S.; Heck, A.J.R. Toward an Optimized Workflow for Middle-Down Proteomics. Anal. Chem. 2017, 89, 3318–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neagu, A.-N.; Jayathirtha, M.; Whitham, D.; Mutsengi, P.; Sullivan, I.; Petre, B.A.; Darie, C.C. Proteomics-Based Identification of Dysregulated Proteins in Breast Cancer. Proteomes 2022, 10, 35. [Google Scholar] [CrossRef]
- Mosavi, Z.; Fakhar, H.B.Z.; Rezaei-Tavirani, M.; Akbari, M.E.; Rostami, F. Proteome profiling of ductal carcinoma in situ. Breast Dis. 2023, 41, 513–520. [Google Scholar] [CrossRef]
- Braga, F.H.G.; Gómez-Mendoza, D.P.; Lemos, R.P.; Rodrigues-Ribeiro, L.; Raíssa-Oliveira, B.; Rodrigues, A.L.P.; Gorshkov, V.; Kjeldsen, F.; Cruz, J.S.; Verano-Braga, T. Proteomic analysis reveals stage-specific reprogramed metabolism for the primary breast cancer cell lines MGSO-3 and MACL-1. Proteomics 2022, 22, 2200095. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-T.; Ding, J.-N.; Wang, J.-L.; Li, Z.-S.; Ding, Y.-L.; Ma, R. Differences in pathologic characteristics between ductal carcinoma in situ (DCIS), DCIS with microinvasion and DCIS with invasive ductal carcinoma. Int. J. Clin. Exp. Pathol. 2020, 13, 1066–1072. [Google Scholar] [PubMed]
- Smith, Y.E.; Wang, G.; Flynn, C.L.; Madden, S.F.; MacEneaney, O.; Cruz, R.G.B.; Richards, C.E.; Jahns, H.; Brennan, M.; Cremona, M.; et al. Functional Antagonism of Junctional Adhesion Molecule-A (JAM-A), Overexpressed in Breast Ductal Carcinoma In Situ (DCIS), Reduces HER2-Positive Tumor Progression. Cancers 2022, 14, 1303. [Google Scholar] [CrossRef] [PubMed]
- Tamburro, D.; Fredolini, C.; Espina, V.; Douglas, T.A.; Ranganathan, A.; Ilag, L.; Zhou, W.; Russo, P.; Espina, B.H.; Muto, G.; et al. Multifunctional Core–Shell Nanoparticles: Discovery of Previously Invisible Biomarkers. J. Am. Chem. Soc. 2011, 133, 19178–19188. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhu, J.; He, K.; Zhao, L.-Y.; Liu, L.-Y.; Song, T.-S.; Huang, C. Proteomic Profiling of Invasive Ductal Carcinoma (IDC) using Magnetic Beads-based Serum Fractionation and MALDI-TOF MS. J. Clin. Lab. Anal. 2015, 29, 321–327. [Google Scholar] [CrossRef]
- Garza, K.Y.; Zhang, J.; Lin, J.Q.; Carter, S.; Suliburk, J.; Nagi, C.; Eberlin, L.S. Abstract P1-20-04: Advanced development of the MasSpec Pen technology to aid in breast cancer surgical margin evaluation and diagnosis during surgery. Cancer Res. 2020, 80 (Suppl. 4), P1-20-04. [Google Scholar] [CrossRef]
- Woolman, M.; Gribble, A.; Bluemke, E.; Zou, J.; Ventura, M.; Bernards, N.; Wu, M.; Ginsberg, H.J.; Das, S.; Vitkin, A.; et al. Optimized Mass Spectrometry Analysis Workflow with Polarimetric Guidance for ex vivo and in situ Sampling of Biological Tissues. Sci. Rep. 2017, 7, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänel, L.; Kwiatkowski, M.; Heikaus, L.; Schlüter, H.; Zhou, M.; Veenstra, T.D.; Lederman, L.; Lewis, J.K.; Krone, J.R.; Nelson, R.W.; et al. Mass spectrometry-based intraoperative tumor diagnostics. Future Sci. OA 2019, 5, FSO373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carcoforo, P.; Ura, B.; Mischiati, C.; Squerzanti, M.; Lanzara, V.; Cervellati, C.; Calza, R.; De Laureto, P.P.; Frare, E.; Portinari, M.; et al. Comparative proteomic analysis of ductal breast carcinoma demonstrates an altered expression of chaperonins and cytoskeletal proteins. Mol. Med. Rep. 2013, 7, 1700–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabbage, M.; Chahed, K.; Hamrita, B.; Guillier, C.L.; Trimeche, M.; Remadi, S.; Hoebeke, J.; Chouchane, L. Protein alterations in infiltrating ductal carcinomas of the breast as detected by nonequilibrium pH gradient electrophoresis and mass spectrometry. J. Biomed. Biotechnol. 2008, 2008, 564127. [Google Scholar] [CrossRef] [Green Version]
- Röwer, C.; Koy, C.; Hecker, M.; Reimer, T.; Gerber, B.; Thiesen, H.-J.; Glocker, M.O. Mass Spectrometric Characterization of Protein Structure Details Refines the Proteome Signature for Invasive Ductal Breast Carcinoma. J. Am. Soc. Mass Spectrom. 2011, 22, 440–456. [Google Scholar] [CrossRef] [Green Version]
- Raso, C.; Cosentino, C.; Gaspari, M.; Malara, N.; Han, X.; McClatchy, D.; Park, S.K.; Renne, M.; Vadalà, N.; Prati, U.; et al. Characterization of Breast Cancer Interstitial Fluids by TmT Labeling, LTQ-Orbitrap Velos Mass Spectrometry, and Pathway Analysis. J. Proteome Res. 2012, 11, 3199–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.L.; Xiao, C.; Fu, G.; Wang, X.; Li, L. Identification of potential serum biomarkers for breast cancer using a functional proteomics technology. Biomark. Res. 2017, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamrita, B.; Chahed, K.; Trimeche, M.; Guillier, C.L.; Hammann, P.; Chaïeb, A.; Korbi, S.; Chouchane, L. Proteomics-based identification of α1-antitrypsin and haptoglobin precursors as novel serum markers in infiltrating ductal breast carcinomas. Clin. Chim. Acta 2009, 404, 111–118. [Google Scholar] [CrossRef]
- Daily, A.; Ravishankar, P.; Harms, S.; Klimberg, V.S. Using tears as a non-invasive source for early detection of breast cancer. PLoS ONE 2022, 17, e0267676. [Google Scholar] [CrossRef]
- Lebrecht, A.; Boehm, D.; Schmidt, M.; Koelbl, H.; Schwirz, R.L.; Grus, F.H. Diagnosis of breast cancer by tear proteomic pattern. Cancer Genom. Proteom. 2009, 6, 177–182. [Google Scholar]
- Brunoro, G.V.F.; Carvalho, P.C.; Barbosa, V.C.; Pagnoncelli, D.; De Moura Gallo, C.V.; Perales, J.; Zahedi, R.P.; Valente, R.H.; De Costa Neves-Ferreira, A.G. Differential proteomic comparison of breast cancer secretome using a quantitative paired analysis workflow. BMC Cancer 2019, 19, 365. [Google Scholar] [CrossRef] [Green Version]
- Brunoro, G.V.F.; Carvalho, P.C.; de Silva Ferreira, A.T.; Perales, J.; Valente, R.H.; de Moura Gallo, C.V.; Pagnoncelli, D.; De Costa Neves-Ferreira, A.G. Proteomic profiling of nipple aspirate fluid (NAF): Exploring the complementarity of different peptide fractionation strategies. J. Proteom. 2015, 117, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Kuerer, H.M.; Goldknopf, I.L.; Fritsche, H.; Krishnamurthy, M.D.S.K.; Sheta, E.A.; Hunt, K.K. Identification of distinct protein expression patterns in bilateral matched pair breast ductal fluid specimens from women with unilateral invasive breast carcinoma. Cancer 2002, 95, 2276–2282. [Google Scholar] [CrossRef]
- Li, J.; Zhao, J.; Yu, X.; Lange, J.; Kuerer, H.; Krishnamurthy, S.; Schilling, E.; Khan, S.A.; Sukumar, S.; Chan, D.W. Identification of Biomarkers for Breast Cancer in Nipple Aspiration and Ductal Lavage Fluid. Clin. Cancer Res. 2005, 11, 8312–8320. [Google Scholar] [CrossRef] [Green Version]
- Beretov, J.; Wasinger, V.; Millar, E.; Schwartz, P.; Graham, P.H.; Li, Y. Proteomic Analysis of Urine to Identify Breast Cancer Biomarker Candidates Using a Label-Free LC-MS/MS Approach. PLoS ONE 2015, 10, e0141876. [Google Scholar] [CrossRef]
- Giri, K.; Mehta, A.; Ambatipudi, K. In search of the altering salivary proteome in metastatic breast and ovarian cancers. FASEB BioAdvances 2019, 1, 191–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslebagh, R.; Whitham, D.; Channaveerappa, D.; Mutsengi, P.; Pentecost, B.T.; Arcaro, K.F.; Darie, C.C. Mass Spectrometry-Based Proteomics of Human Milk to Identify Differentially Expressed Proteins in Women with Breast Cancer versus Controls. Proteomes 2022, 10, 36. [Google Scholar] [CrossRef]
- Aslebagh, R.; Channaveerappa, D.; Arcaro, K.F.; Darie, C.C. Proteomics analysis of human breast milk to assess breast cancer risk. Electrophoresis 2018, 39, 653–665. [Google Scholar] [CrossRef]
- Aslebagh, R.; Channaveerappa, D.; Arcaro, K.F.; Darie, C.C. Comparative two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) of human milk to identify dysregulated proteins in breast cancer. Electrophoresis 2018, 39, 1723–1734. [Google Scholar] [CrossRef] [PubMed]
- Aslebagh, R.; Channaveerappa, D.; Pentecost, B.T.; Arcaro, K.F.; Darie, C.C. Combinatorial Electrophoresis and Mass Spectrometry-Based Proteomics in Breast Milk for Breast Cancer Biomarker Discovery. In Advancements of Mass Spectrometry in Biomedical Research; Woods, A.G., Darie, C.C., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 451–467. [Google Scholar]
- Milioli, H.H.; Sousa, K.S.; Kaviski, R.; Oliveira, N.C.D.S.; Urban, C.D.A.; De Lima, R.S.; Cavalli, I.J.; Ribeiro, E.M.D.S.F. Comparative Proteomics of Primary Breast Carcinomas and Lymph Node Metastases Outlining Markers of Tumor Invasion. Cancer Genom. Proteom. 2015, 12, 89–101. [Google Scholar]
- Pozniak, Y.; Balint-Lahat, N.; Rudolph, J.D.; Lindskog, C.; Katzir, R.; Avivi, C.; Pontén, F.; Ruppin, E.; Barshack, I.; Geiger, T. System-wide Clinical Proteomics of Breast Cancer Reveals Global Remodeling of Tissue Homeostasis. Cell Syst. 2016, 2, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Zhang, Y.; Zhang, A.; Li, Y.; Zhu, Z.; Shao, Z.; Zeng, R.; Xu, L.X. Comparative serum proteome analysis of human lymph node negative/positive invasive ductal carcinoma of the breast and benign breast disease controls via label-free semiquantitative shotgun technology. Omics J. Integr. Biol. 2009, 13, 291–300. [Google Scholar] [CrossRef]
- Al-Wajeeh, A.S.; Salhimi, S.M.; Al-Mansoub, M.A.; Khalid, I.A.; Harvey, T.M.; Latiff, A.; Ismail, M.N. Comparative proteomic analysis of different stages of breast cancer tissues using ultra high performance liquid chromatography tandem mass spectrometer. PLoS ONE 2020, 15, e0227404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, N.; Gomig, T.; Milioli, H.; Cordeiro, F.; Costa, G.; Urban, C.; Lima, R.; Cavalli, I.; Ribeiro, E. Comparative proteomic analysis of ductal and lobular invasive breast carcinoma. Genet. Mol. Res. GMR 2016, 15, 15027701. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.A.; Mikesh, L.; Moskulak, C.; Harvey, J.; Sherman, N.; Zigrino, P.; Mauch, C.; Fox, J.W. Host Response to Human Breast Invasive Ductal Carcinoma (IDC) as Observed by Changes in the Stromal Proteome. J. Proteome Res. 2014, 13, 4739–4751. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Jung, W.H.; Koo, J.S. Expression of cancer-associated fibroblast-related proteins differs between invasive lobular carcinoma and invasive ductal carcinoma. Breast Cancer Res. Treat. 2016, 159, 55–69. [Google Scholar] [CrossRef]
- Tomko, L.A.; Hill, R.C.; Barrett, A.; Szulczewski, J.M.; Conklin, M.W.; Eliceiri, K.W.; Keely, P.J.; Hansen, K.C.; Ponik, S.M. Targeted matrisome analysis identifies thrombospondin-2 and tenascin-C in aligned collagen stroma from invasive breast carcinoma. Sci. Rep. 2018, 8, 12941. [Google Scholar] [CrossRef] [Green Version]
- Di Cara, G.; Marabeti, M.R.; Musso, R.; Riili, I.; Cancemi, P.; Minafra, I.P. New Insights into the Occurrence of Matrix Metalloproteases -2 and -9 in a Cohort of Breast Cancer Patients and Proteomic Correlations. Cells 2018, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Du, J.; Wang, Y.; Jia, H. A Coagulation-Related Gene-Based Prognostic Model for Invasive Ductal Carcinoma. Front. Genet. 2021, 12, 722992. [Google Scholar] [CrossRef]
- Yoneten, K.K.; Kasap, M.; Akpinar, G.; Gunes, A.; Gurel, B.; Utkan, N.Z. Comparative Proteome Analysis of Breast Cancer Tissues Highlights the Importance of Glycerol-3-phosphate Dehydrogenase 1 and Monoacylglycerol Lipase in Breast Cancer Metabolism. Cancer Genom. Proteom. 2019, 16, 377–397. [Google Scholar] [CrossRef] [Green Version]
- Böhm, D.; Keller, K.; Pieter, J.; Boehm, N.; Wolters, D.; Siggelkow, W.; Lebrecht, A.; Schmidt, M.; Kölbl, H.; Pfeiffer, N.; et al. Comparison of tear protein levels in breast cancer patients and healthy controls using a de novo proteomic approach. Oncol. Rep. 2012, 28, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sun, L.; Li, H.; Ai, L.; Ma, Q.; Qiao, X.; Yang, J.; Zhang, H.; Ou, X.; Wang, Y.; et al. Binding blockade between TLN1 and integrin β1 represses triple-negative breast cancer. eLife 2022, 11, e68481. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, K.; Minashi, K.; Igaki, H.; Tachimori, Y.; Nishimura, T.; Hokamura, N.; Ashida, A.; Daiko, H.; Ochiai, A.; Muto, M.; et al. Artificially Induced Epithelial-Mesenchymal Transition in Surgical Subjects: Its Implications in Clinical and Basic Cancer Research. PLoS ONE 2011, 6, e18196. [Google Scholar] [CrossRef] [PubMed]
- Angel, P.M.; Schwamborn, K.; Comte-Walters, S.; Clift, C.L.; Ball, L.E.; Mehta, A.S.; Drake, R.R. Extracellular Matrix Imaging of Breast Tissue Pathologies by MALDI–Imaging Mass Spectrometry. Proteom.—Clin. Appl. 2019, 13, e1700152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, E.S.; Ertel, A.; Davicioni, E.; Kline, J.; Schwartz, G.F.; Witkiewicz, A.K. Progression of ductal carcinoma in situ to invasive breast cancer is associated with gene expression programs of EMT and myoepithelia. Breast Cancer Res. Treat. 2012, 133, 1009–1024. [Google Scholar] [CrossRef]
- Gawin, M.; Kurczyk, A.; Niemiec, J.; Stanek-Widera, A.; Grela-Wojewoda, A.; Adamczyk, A.; Biskup-Frużyńska, M.; Polańska, J.; Widłak, P. Intra-Tumor Heterogeneity Revealed by Mass Spectrometry Imaging Is Associated with the Prognosis of Breast Cancer. Cancers 2021, 13, 4349. [Google Scholar] [CrossRef]
- Martinez, J.; Smith, P. The Dynamic Interaction between Extracellular Matrix Remodeling and Breast Tumor Progression. Cells 2021, 10, 1046. [Google Scholar] [CrossRef]
- Gu, Y.; Tang, S.; Wang, Z.; Cai, L.; Lian, H.; Shen, Y.; Zhou, Y. A pan-cancer analysis of the prognostic and immunological role of β-actin (ACTB) in human cancers. Bioengineered 2021, 12, 6166–6185. [Google Scholar] [CrossRef]
- Gu, Y.; Tang, S.; Wang, Z.; Cai, L.; Lian, H.; Shen, Y.; Zhou, Y. Role of cellular cytoskeleton in epithelial-mesenchymal transition process during cancer progression. Biomed. Rep. 2015, 3, 603–610. [Google Scholar]
- Lopes, D.; Maiato, H. The Tubulin Code in Mitosis and Cancer. Cells 2020, 9, 2356. [Google Scholar] [CrossRef]
- Saha, S.K.; Kim, K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. Cytokeratin 19 (KRT19) has a Role in the Reprogramming of Cancer Stem Cell-Like Cells to Less Aggressive and More Drug-Sensitive Cells. Int. J. Mol. Sci. 2018, 19, 1423. [Google Scholar] [CrossRef] [Green Version]
- Polioudaki, H.; Agelaki, S.; Chiotaki, R.; Politaki, E.; Mavroudis, D.; Matikas, A.; Georgoulias, V.; Theodoropoulos, P.A. Variable expression levels of keratin and vimentin reveal differential EMT status of circulating tumor cells and correlation with clinical characteristics and outcome of patients with metastatic breast cancer. BMC Cancer 2015, 15, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khandeparkar, S.G.S.; Khillare, C.D.; Joshi, A.R.; Kulkarni, M.M.; Gogate, B.P.; Battin, S. Immunohistochemical Expression of Vimentin in Invasive Breast Carcinoma and Its Correlation with Clinicopathological Parameters. Niger. Med. J. 2019, 60, 17. [Google Scholar] [CrossRef]
- Wieczorek, K.; Wiktorska, M.; Sacewicz-Hofman, I.; Boncela, J.; Lewiński, A.; Kowalska, M.A.; Niewiarowska, J. Filamin A upregulation correlates with Snail-induced epithelial to mesenchymal transition (EMT) and cell adhesion but its inhibition increases the migration of colon adenocarcinoma HT29 cells. Exp. Cell Res. 2017, 359, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Zografos, E.; Anagnostopoulos, A.K.; Papadopoulou, A.; Legaki, E.; Zagouri, F.; Marinos, E.; Tsangaris, G.T.; Gazouli, M. Serum Proteomic Signatures of Male Breast Cancer. Cancer Genom. Proteom. 2019, 16, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yang, Y.; Du, B. Clinical Characterization and Prognostic Value of TPM4 and Its Correlation with Epithelial-Mesenchymal Transition in Glioma. Brain Sci. 2022, 12, 1120. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Du, B. Profilin 1, negatively regulated by microRNA-19a-3p, serves as a tumor suppressor in human hepatocellular carcinoma. Pathol.—Res. Pract. 2019, 215, 499–505. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Du, B. Gelsolin Promotes Cancer Progression by Regulating Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma and Correlates with a Poor Prognosis. J. Oncol. 2020, 2020, 1980368. [Google Scholar]
- Xu, J.; Huang, Y.; Zhao, J.; Wu, L.; Qi, Q.; Liu, Y.; Li, G.; Li, J.; Liu, H.; Wu, H. Cofilin: A Promising Protein Implicated in Cancer Metastasis and Apoptosis. Front. Cell Dev. Biol. 2021, 9, 599065. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, Q.; Wan, C.; Sun, S.; Zhu, L.; Zhao, Z.; Wang, B. Transgelin Inhibits the Malignant Progression of Esophageal Squamous Cell Carcinomas by Regulating Epithelial–Mesenchymal Transition. Front. Oncol. 2021, 11, 3313. [Google Scholar]
- Chen, M.-J.; Gao, X.-J.; Xu, L.-N.; Liu, T.-F.; Liu, X.-H.; Liu, L.-X. Ezrin is required for epithelial-mesenchymal transition induced by TGF-β1 in A549 cells. Int. J. Oncol. 2014, 45, 1515–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Wan, F.; Zhang, H.; Shi, G.; Ye, D. ITGA2B and ITGA8 are predictive of prognosis in clear cell renal cell carcinoma patients. Tumor Biol. 2016, 37, 253–262. [Google Scholar] [CrossRef]
- Azizi, L.; Cowell, A.R.; Mykuliak, V.V.; Goult, B.T.; Turkki, P.; Hytönen, V.P. Cancer associated talin point mutations disorganise cell adhesion and migration. Sci. Rep. 2021, 11, 347. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Duan, H.; Wang, S.; Quan, Y.-J.; Huang, J.-H.; Guo, Z.-C. Talin1 Induces Epithelial-Mesenchymal Transition to Facilitate Endometrial Cell Migration and Invasion in Adenomyosis Under the Regulation of microRNA-145-5p. Reprod. Sci. 2021, 28, 1523–1539. [Google Scholar] [CrossRef]
- Fedele, M.; Sgarra, R.; Battista, S.; Cerchia, L.; Manfioletti, G. The Epithelial & ndash; Mesenchymal Transition at the Crossroads between Metabolism and Tumor Progression. Int. J. Mol. Sci. 2022, 23, 800. [Google Scholar]
- Sciacovelli, M.; Frezza, C. Metabolic reprogramming and epithelial-to-mesenchymal transition in cancer. FEBS J. 2017, 284, 3132–3144. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, T.; Verdone, J.E.; Huang, J.; Kahlert, U.D.; Hernandez, J.R.; Torga, G.; Zarif, J.C.; Epstein, T.; Gatenby, R.; McCartney, A.; et al. Glycolysis is the primary bioenergetic pathway for cell motility and cytoskeletal remodeling in human prostate and breast cancer cells. Oncotarget 2014, 6, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Bourcy, M.; Suarez-Carmona, M.; Lambert, J.; Francart, M.-E.; Schroeder, H.; Delierneux, C.; Skrypek, N.; Thompson, E.W.; Jérusalem, G.; Berx, G.; et al. Tissue Factor Induced by Epithelial–Mesenchymal Transition Triggers a Procoagulant State That Drives Metastasis of Circulating Tumor Cells. Cancer Res. 2016, 76, 4270–4282. [Google Scholar] [CrossRef] [Green Version]
- Dongre, A.; Ortiz-Cuaran, S.; Korkaya, H. Editorial: The Role of the EMT Program in Regulating the Immune Response in Carcinoma. Front. Immunol. 2022, 13, 940164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, Q.; Li, T.; Liao, Q.; Zhao, Y. Role of the complement system in the tumor microenvironment. Cancer Cell Int. 2019, 19, 300. [Google Scholar] [CrossRef]
- Zarychta, E.; Ruszkowska-Ciastek, B. Cooperation between Angiogenesis, Vasculogenesis, Chemotaxis, and Coagulation in Breast Cancer Metastases Development: Pathophysiological Point of View. Biomedicines 2022, 10, 300. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ju, M.K.; Jeon, H.M.; Lee, Y.J.; Kim, C.H.; Park, H.G.; Kang, H.S. Reactive oxygen species induce epithelial-mesenchymal transition, glycolytic switch, and mitochondrial repression through the Dlx-2/Snail signaling pathways in MCF-7 cells. Mol. Med. Rep. 2019, 20, 2339–2346. [Google Scholar] [CrossRef]
- Felipe Lima, J.; Nofech-Mozes, S.; Bayani, J.; Bartlett, J.M. EMT Breast Carcinoma-A Review. J. Clin. Med. 2016, 5, 65. [Google Scholar]
- Lee, S.; Stewart, S.; Nagtegaal, I.; Luo, J.; Wu, Y.; Colditz, G.; Medina, D.; Allred, D.C. Differentially expressed genes regulating the progression of ductal carcinoma in situ to invasive breast cancer. Cancer Res. 2012, 72, 4574–4586. [Google Scholar] [CrossRef] [Green Version]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.T.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byron, A.; Humphries, J.; Humphries, M. Defining the extracellular matrix using proteomics. Int. J. Exp. Pathol. 2013, 94, 75–92. [Google Scholar] [CrossRef]
- Santi, A.; Kugeratski, F.G.; Zanivan, S. Cancer Associated Fibroblasts: The Architects of Stroma Remodeling. Proteomics 2018, 18, e1700167. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Hoersch, S.; Liu, H.; Carr, S.A.; Hynes, R.O. The Matrisome: In Silico Definition and In Vivo Characterization by Proteomics of Normal and Tumor Extracellular Matrices*. Mol. Cell. Proteom. 2012, 11, M111.014647. [Google Scholar] [CrossRef] [Green Version]
- Crake, R.L.I.; Phillips, E.; Kleffmann, T.; Currie, M.J. Co-culture With Human Breast Adipocytes Differentially Regulates Protein Abundance in Breast Cancer Cells. Cancer Genom.—Proteom. 2019, 16, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Haerinck, J.; Berx, G. Partial EMT takes the lead in cancer metastasis. Dev. Cell 2021, 56, 3174–3176. [Google Scholar] [CrossRef]
- Cheng, X.; Li, F.; Tao, Z. Tenascin-C promotes epithelial-to-mesenchymal transition and the mTOR signaling pathway in nasopharyngeal carcinoma. Oncol. Lett. 2021, 22, 570. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Xu, H.; Wang, W.; Li, S.; Li, H.; Li, T.; Zhang, W.; Yu, X.; Liu, L. The role of collagen in cancer: From bench to bedside. J. Transl. Med. 2019, 17, 309. [Google Scholar] [CrossRef] [Green Version]
- Koorman, T.; Jansen, K.A.; Khalil, A.; Haughton, P.D.; Visser, D.; Rätze, M.A.K.; Haakma, W.E.; Sakalauskaitè, G.; van Diest, P.J.; de Rooij, J.; et al. Spatial collagen stiffening promotes collective breast cancer cell invasion by reinforcing extracellular matrix alignment. Oncogene 2022, 41, 2458–2469. [Google Scholar] [CrossRef]
- Li, C.-L.; Yang, D.; Cao, X.; Wang, F.; Hong, D.-Y.; Wang, J.; Shen, X.-C.; Chen, Y. Fibronectin induces epithelial-mesenchymal transition in human breast cancer MCF-7 cells via activation of calpain. Oncol. Lett. 2017, 13, 3889–3895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, Y.; Taniyama, Y.; Sanada, F.; Morishita, R.; Nakamori, S.; Morimoto, K.; Yeung, K.T.; Yang, J. Periostin blockade overcomes chemoresistance via restricting the expansion of mesenchymal tumor subpopulations in breast cancer. Sci. Rep. 2018, 8, 4013. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak-Wielgomas, K.; Grzegrzolka, J.; Piotrowska, A.; Gomulkiewicz, A.; Witkiewicz, W.; Dziegiel, P. Periostin expression in cancer-associated fibroblasts of invasive ductal breast carcinoma. Oncol. Rep. 2016, 36, 2745–2754. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Sun, L.; Yuan, X.; Qiu, H. Thrombospondin-1 is a multifaceted player in tumor progression. Oncotarget 2017, 8, 84546–84558. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Villodre, E.S.; Larson, R.; Rahal, O.M.; Wang, X.; Gong, Y.; Debeb, B.G. Decorin, a novel negative modulator of E-cadherin in inflammatory breast cancer. bioRxiv 2020, bioRxiv:2020.07.07.190496. [Google Scholar]
- Karamanou, K.; Franchi, M.; Vynios, D.; Brézillon, S. Epithelial-to-mesenchymal transition and invadopodia markers in breast cancer: Lumican a key regulator. Semin. Cancer Biol. 2020, 62, 125–133. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, R.; Dong, M.; Zhang, Z.; Li, H.; Zhan, C.; Li, X. Osteoglycin (OGN) Inhibits Cell Proliferation and Invasiveness in Breast Cancer via PI3K/Akt/mTOR Signaling Pathway. OncoTargets Ther. 2019, 12, 10639–10650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshkian, Z.; Nobili, S.; Peyravian, N.; Shojaee, B.; Nazari, H.; Soleimani, H.; Asadzadeh-Aghdaei, H.; Bonab, M.A.; Nazemalhosseini-Mojarad, E.; Mini, E. Insights into the Role of Matrix Metalloproteinases in Precancerous Conditions and in Colorectal Cancer. Cancers 2021, 13, 6226. [Google Scholar] [CrossRef] [PubMed]
- Neagu, A.N.; Jayathirtha, M.; Whitham, D.; Mutsengi, P.; Sullivan, I.; Petre, B.A.; Darie, C.C. Proteomics and its applications in breast cancer. Am. J. Cancer Res. 2021, 11, 4006–4049. [Google Scholar]
- Li, J.; Guan, X.; Fan, Z.; Ching, L.-M.; Li, Y.; Wang, X.; Cao, W.-M.; Liu, D.-X. Non-Invasive Biomarkers for Early Detection of Breast Cancer. Cancers 2020, 12, 2767. [Google Scholar] [CrossRef]
- Veyssière, H.; Bidet, Y.; Penault-Llorca, F.; Radosevic-Robin, N.; Durando, X. Circulating proteins as predictive and prognostic biomarkers in breast cancer. Clin. Proteom. 2022, 19, 25. [Google Scholar] [CrossRef]
- Capone, E.; Iacobelli, S.; Sala, G. Role of galectin 3 binding protein in cancer progression: A potential novel therapeutic target. J. Transl. Med. 2021, 19, 405. [Google Scholar] [CrossRef]
- Lin, T.-W.; Chang, H.-T.; Chen, C.-H.; Lin, S.-W.; Hsu, T.-L.; Wong, C.-H. Galectin-3 Binding Protein and Galectin-1 Interaction in Breast Cancer Cell Aggregation and Metastasis. J. Am. Chem. Soc. 2015, 137, 9685–9693. [Google Scholar] [CrossRef] [PubMed]
- White, M.J.V.; Roife, D.; Gomer, R.H. Galectin-3 Binding Protein Secreted by Breast Cancer Cells Inhibits Monocyte-Derived Fibrocyte Differentiation. J. Immunol. 2015, 195, 1858–1867. [Google Scholar] [CrossRef] [Green Version]
- Piccolo, E.; Tinari, N.; Semeraro, D.; Traini, S.; Fichera, I.; Cumashi, A.; La Sorda, R.; Spinella, F.; Bagnato, A.; Lattanzio, R.; et al. LGALS3BP, lectin galactoside-binding soluble 3 binding protein, induces vascular endothelial growth factor in human breast cancer cells and promotes angiogenesis. J. Mol. Med. 2013, 91, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Noma, N.; Simizu, S.; Kambayashi, Y.; Kabe, Y.; Suematsu, M.; Umezawa, K. Involvement of NF-κB-mediated expression of galectin-3-binding protein in TNF-α-induced breast cancer cell adhesion. Oncol. Rep. 2012, 27, 2080–2084. [Google Scholar]
- Koths, K.; Taylor, E.; Halenbeck, R.; Casipit, C.; Wang, A. Cloning and characterization of a human Mac-2-binding protein, a new member of the superfamily defined by the macrophage scavenger receptor cysteine-rich domain. J. Biol. Chem. 1993, 268, 14245–14249. [Google Scholar] [CrossRef] [PubMed]
- Whelan, S.A.; He, J.; Lu, M.; Souda, P.; Saxton, R.E.; Faull, K.F.; Whitelegge, J.P.; Chang, H.R. Mass spectrometry (LC-MS/MS) identified proteomic biosignatures of breast cancer in proximal fluid. J. Proteome Res. 2012, 11, 5034–5045. [Google Scholar] [CrossRef] [Green Version]
- Mbeunkui, F.; Metge, B.J.; Shevde, L.A.; Pannell, L.K. Identification of differentially secreted biomarkers using LC-MS/MS in isogenic cell lines representing a progression of breast cancer. J. Proteome Res. 2007, 6, 2993–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwedar, F.I.; Khalil, G.I.; Nayer, S.A.; Farouk, A. Aberrant Vimentin Methylation Is Characteristic of Breast Cancer. Adv. Breast Cancer Res. 2016, 5, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Ulirsch, J.; Fan, C.; Knafl, G.; Wu, M.J.; Coleman, B.; Perou, C.M.; Swift-Scanlan, T. Vimentin DNA methylation predicts survival in breast cancer. Breast Cancer Res. Treat. 2013, 137, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Wang, W.; Wang, Q.; Zhang, Y.; Zahid, K.R.; Raza, U.; Gong, Y. Alpha-1-antichymotrypsin as a novel biomarker for diagnosis, prognosis, and therapy prediction in human diseases. Cancer Cell Int. 2022, 22, 156. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, J.Y. A method for identifying discriminative isoform-specific peptides for clinical proteomics application. BMC Genom. 2016, 17 (Suppl. 7), 522. [Google Scholar] [CrossRef] [Green Version]
- Pateetin, P.; Pisitkun, T.; McGowan, E.; Boonyaratanakornkit, V. Differential quantitative proteomics reveals key proteins related to phenotypic changes of breast cancer cells expressing progesterone receptor A. J. Steroid Biochem. Mol. Biol. 2020, 198, 105560. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, M.; Michael, T.; Drabier, R. Novel alternative splicing isoform biomarkers identification from high-throughput plasma proteomics profiling of breast cancer. BMC Syst. Biol. 2013, 7, S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, K.; Jaehnig, E.J.; Satpathy, S.; Blumenberg, L.; Karpova, A.; Anurag, M.; Miles, G.; Mertins, P.; Geffen, Y.; Tang, L.C.; et al. Proteogenomic Landscape of Breast Cancer Tumorigenesis and Targeted Therapy. Cell 2020, 183, 1436–1456.e31. [Google Scholar] [CrossRef]
- Pani, T.; Rajput, K.; Kar, A.; Sharma, H.; Basak, R.; Medatwal, N.; Saha, S.; Dev, G.; Kumar, S.; Gupta, S.; et al. Alternative splicing of ceramide synthase 2 alters levels of specific ceramides and modulates cancer cell proliferation and migration in Luminal B breast cancer subtype. Cell Death Dis. 2021, 12, 171. [Google Scholar] [CrossRef]

| Relevance | Biological Samples | Other Conventional Analytical and Coupled Methods | MS-Based Proteomics | Results | Dysregulated Pathways and BP | References |
|---|---|---|---|---|---|---|
| Comparison of IDC vs. healthy counterparts; comparison of IDC vs. cytosarcoma phyllodes | interstitial fluid, primary cell culture | FC, IF, WB | TmT, HPLC–MS/MS + MudPIT | DEPs, quantitative proteomic profile | EMT; IFγ pathway; cell invasion, motility, survival, adhesion, cell cycle/proliferation, Wnt signaling, proteasome, and apoptosis | [65] |
| FF | 2-DE | MALDI-ToF MS | activity levels of MMP-2 and MMP-9 much higher in IDC | ECM remodeling | [88] | |
| FF | 2-DE, DIGE | MALDI-ToF/ToF | proteome profiling | triacylglyceride (TAG) metabolism | [90] | |
| Comparison of IDC vs. ILC | IDC and ILC tissue samples | 2-DE | MALDI-ToF/ToF MS | proteome profiling | AJ; EMT; GLYCOLYSIS | [84] |
| FFPE ILC and IDC-NST tissue samples | TMA, H&E, IHC | - | comparison of CAFs-related proteins | cell growth, angiogenesis, macrophage recruitment, ECM remodeling | [86] | |
| Comparison of IDC vs. other cancers (ovarian, lung, prostate, colon cancers, and melanoma) vs. healthy controls | saliva | WB, IHC | nLC–MS/MS | DEPs; PPI | cell-motility related proteins, cytoskeletal organization, ECM remodeling | [75] |
| serum | nanoparticle-based protein enrichment technology | LC–MS/MS | IDC-specific protein signatures | candidate biomarkers | [3] | |
| Comparison of IDC vs. healthy/benign controls | tissue samples | 2D SDS-PAGE; IHC | LC–MS/MS; MALDI-ToF-MS | DEPs and candidate biomarkers | deregulated chaperonins, stress-related proteins, cytoskeletal proteins involved in motility mechanisms, metabolic enzymes, immunologic responses | [62,63,64] |
| serum | magnetic bead-based serum fractionation | MALDI-ToF MS | serum protein profiling | metabolic enzymes and protease activity as biomarkers for diagnosis and drug development; protein isoforms detection | [58,66,67] | |
| nanoparticle-based protein enrichment technology | LC–MS/MS | LMW and protein fragments; IDC-specific protein signatures; early-stage IDC ECM biomarkers | EMT/migration, cell proliferation, adhesion and metastasis | [3] | ||
| milk | 1D-SDS-PAGE, 2D-PAGE | nLC–MS/MS | protein profiling, DEPs | putative biomarkers | [76,77,78,79] | |
| tear fluid | 1D-SDS-PAGE, ELISA | SELDI-ToF MS; MALDI-ToF/ToF; LC–MS/MS | biomarkers for early detection | metabolic reprogramming; immune response | [68,69,91] | |
| Comparison of bilateral matched pair NAF/DLF proteomes in unilateral IDC | NAF/DLF | 2D-PAGE, SRM, ELISA | nLC–MS/MS, SELDI-ToF | secretome analysis; abundant DEPs; biomarker discovery and validation | GLYCOLYSIS; COMPLEMENT; cell-stroma communication | [70,71,72,73] |
| Comparison of adjacent healthy/benign breast disease vs. lymph node ± IDC vs. matched LNM | FFPE (ER+/HER2- negative and matched LNM) | pulsed-SILAC assay, IHC | UHPLC-EASY spray ionization source-MS/MS | BC progression; proteomic profiles of LNM similar to those of primary tumors | proteostasis alteration: downregulation of DNA repair proteins; upregulation of ribosomal, lysosomal and proteasomal proteins; elevated rate of protein translation; deregulation of protein folding machinery; increased amounts of unfolded proteins; metabolic reprogramming: OXPHOS and GLYCOLYSIS; ROS upregulation; reduced biosynthesis and increased breakdown of fatty acids, decrease in cholesterol biosynthesis, increase in peroxisomal β-oxidation | [81] |
| IDC PBT and matched LNM | 2-DE | MALDI-ToF/ToF MS | overexpressed proteins in PBT | cytoskeleton reorganization, cell growth and proliferation, ECM remodeling, proteolysis regulation, metabolic reprogramming, detoxification, stress-related mechanisms, membrane-associated proteins | [80] | |
| serum benign, LNM+IDC and LNM-IDC | 2-DE, ELISA | LC–MS/MS | DEPs during IDC progression | putative biomarkers for early metastasis detection | [82] | |
| Comparison of IDC-stages’ specific protein signatures | serum | hydrogel nanoparticles for protein enrichment technology | LC–MS/MS | IDC early-stage proteins; LMW proteins and protein fragments as IDC candidate biomarkers | EMT [92] | [3] |
| IDC PBT (mastectomy) | SDS-PAGE | LC–MS/MS | specific proteins (stage 2 and 3); identification of putative IDC stage-specific biomarkers | stage 2: proliferation, invasion, migration, stress pathways stage 3: invasion, stress, DNA repair, tumor suppression, inflammation, invasion, glycolysis, metastasis | [83] | |
| IDC-subtype specific protein signature and biomarkers discovery | FF | 2-DE, WB | LC–MSE, MALDI-MS/MS | IDC-specific signature of ER+/HER2/neu negative IDC, PPI networks | EMT; cytoskeleton organization; ROS and stress response; Calcium-binding proteins involved in signaling pathways | [23] |
| Comparison of IDC tumor-adjacent stroma vs. tumor-distal stroma | cell lines; FFPE | LCM, IHC | LC–MS/MS | proteomics of breast cell line-stimulated fibroblast ECM vs proteomics of invasive/metastatic stromal tissue | EMT | [85] |
| IDC matrisome-targeted proteomics | FF FFPE | TPM, SHG, IF | LC–MS/MS | ECM proteomic profile as early diagnosis and risk of metastases biomarker and therapeutic target | ECM remodeling: collagen fiber reorganization/alignment | [87] |
| FFPE | TMA, microscopy | MALDI-FT-ICR MSI; HRAM nanoLC-ESI–MS/MS | alteration of multiple collagen patterns in TME | EMT-related biomarkers [93] | [94] | |
| Discovery of IDC candidate biomarkers | urine, cell lines, FFPE | WB, IHC | LC–MS/MS | DEPs, biomarker candidates for early detection | acute phase response signaling, production of NO and ROS in macrophages, IL-12 signaling and production in macrophages, intrinsic prothrombin activation pathway, clathrin-mediated endocytosis signaling, communication between innate and adaptive immune cells | [74] |
| Pathway analysis and biomarker discovery | bioinformatics approach | proteins expression profile, prognostic significance | COAGULATION; EMT; ANGIOGENESIS; UV_RESPONSE_DN; TGF_BETA_SIGNALING; HEDGEHOG_SIGNALING | [89] | ||
| Dysregulated Proteins | Genes | Proteomics-Based Methods | Functions | Associated Roles in Cancer | References |
|---|---|---|---|---|---|
| Actin isoforms | ACTB, ACTG | LC–MSE, MALDI-MS/MS | cytoskeleton structural protein | cell growth, migration, invasion, metastasis [98], EMT [99] | [23] |
| Tubulin isoforms | TUBB, TUBA1A, TUBA1B | MALDI-ToF/ToF MS | constituents of microtubules | chromosome segregation during mitosis [100] | [84] |
| Keratins | KRT19, KRT8 | MALDI-ToF/ToF MS | cytoplasmic intermediate filament proteins | tumorigenic transformation of cells, stemness, cell proliferation, migration [101]; EMT [102] | [80] |
| Vimentin | VIM | IHC; TmT, HPLC–MS/MS + MudPIT; LC–MSE, MALDI-MS/MS; MALDI-ToF/ToF MS | cytoplasmic intermediate filament protein | EMT [103] | [23,65] |
| Filamins | FLNA | nanoHPLC–MS/MS; HNs coupled with LC–MS/MS | actin-binding protein | cancer progression, cell motility, EMT [104] | [3,105] |
| Tropomyosin family | TPM3, TPM4 | salivary LC–MS/MS; MALDI-ToF/ToF MS | actin-binding protein | cell migration, invasion, motility, metastasis, EMT [106] | [75,84] |
| Profilin family | PFN1 | salivary LC–MS/MS; LC–MSE; MALDI-MS/MS | actin-binding protein | cell proliferation, motility, EMT [107] | [23,75] |
| Gelsolin | GSN | salivary LC–MS/MS | actin-binding protein | cell motility, EMT [108] | [75] |
| Cofilin | CFL1 | serum HNs coupled with LC–MS/MS | actin-binding protein | cytoskeletal reorganization, lamellipodium formation, EMT [109] | [3] |
| Transgelin | TAGLN | LC–MSE; MALDI-MS/MS | actin-binding protein | cell growth, ECM degradation, invasion, metastasis, proliferation, EMT [110] | [23,85] |
| Ezrin | EZR | salivary LC–MS/MS | membrane-cytoskeleton linker | cytoskeleton remodeling, EMT [111] | [75] |
| Integrins | ITGA2B | serum HNs coupled with LC–MS/MS | membrane adhesion receptors | adhesion, recognition, immune response, cell growth, metastasis [112] | [3] |
| Talin | TLN1 | serum HNs coupled with LC–MS/MS | component of adhesion complexes | cell migration, adhesion, integrin signaling [113]; EMT [114] | [3] |
| Protein Isoforms | Biological Samples | Other Conventional Analytical and Coupled Methods | MS-Based Proteomics | Results | Functions | References |
|---|---|---|---|---|---|---|
| Folate receptor isoforms (FRα, FRβ); potential isoform-based diagnosis in BC | BC cells lines and IDC tissue | WB, IHC | LC-ESI–MS/MS | simultaneous and accurate quantification of FR isoforms: FRα is overexpressed in BC cells and tissue samples, FRβ is abundant in TAMs | uni-directional folate transport into cells | [27] |
| Progesterone receptor isoforms A and B; PRA/PRB ratios during BC progression | BC cell line model | SDS-PAGE | HPLC–MS/MS | isoform-specific changes in BC proteome; high PRA/PRB ratios in BC associated with resistance to chemotherapy and poor prognosis | cell metabolism, cell cycle, apoptosis | [157] |
| Haptoglobin and α1-AT precursor isoforms | serum | 2-DE; FFPE tissue sections-IHC | MALDI-MS | DEPs; identification of novel serum biomarkers in IDC patients compared with healthy women | possible role in tumor growth | [67] |
| Alternative splicing of ceramide synthase 2 (AS CERS2) | BC cell lines, IDC and adjacent normal tissue | RT-PCR, WB, SDS-PAGE, IHC | LC–MS/MS | higher expression of AS CERS2 in luminal B IDC | dysregulation of sphingolipid pathway, cancer initiation, proliferation and migration, cell survival, apoptosis | [160] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neagu, A.-N.; Whitham, D.; Seymour, L.; Haaker, N.; Pelkey, I.; Darie, C.C. Proteomics-Based Identification of Dysregulated Proteins and Biomarker Discovery in Invasive Ductal Carcinoma, the Most Common Breast Cancer Subtype. Proteomes 2023, 11, 13. https://doi.org/10.3390/proteomes11020013
Neagu A-N, Whitham D, Seymour L, Haaker N, Pelkey I, Darie CC. Proteomics-Based Identification of Dysregulated Proteins and Biomarker Discovery in Invasive Ductal Carcinoma, the Most Common Breast Cancer Subtype. Proteomes. 2023; 11(2):13. https://doi.org/10.3390/proteomes11020013
Chicago/Turabian StyleNeagu, Anca-Narcisa, Danielle Whitham, Logan Seymour, Norman Haaker, Isabella Pelkey, and Costel C. Darie. 2023. "Proteomics-Based Identification of Dysregulated Proteins and Biomarker Discovery in Invasive Ductal Carcinoma, the Most Common Breast Cancer Subtype" Proteomes 11, no. 2: 13. https://doi.org/10.3390/proteomes11020013