Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production


Abstract
:1. Introduction
2. Plant Expression Platform

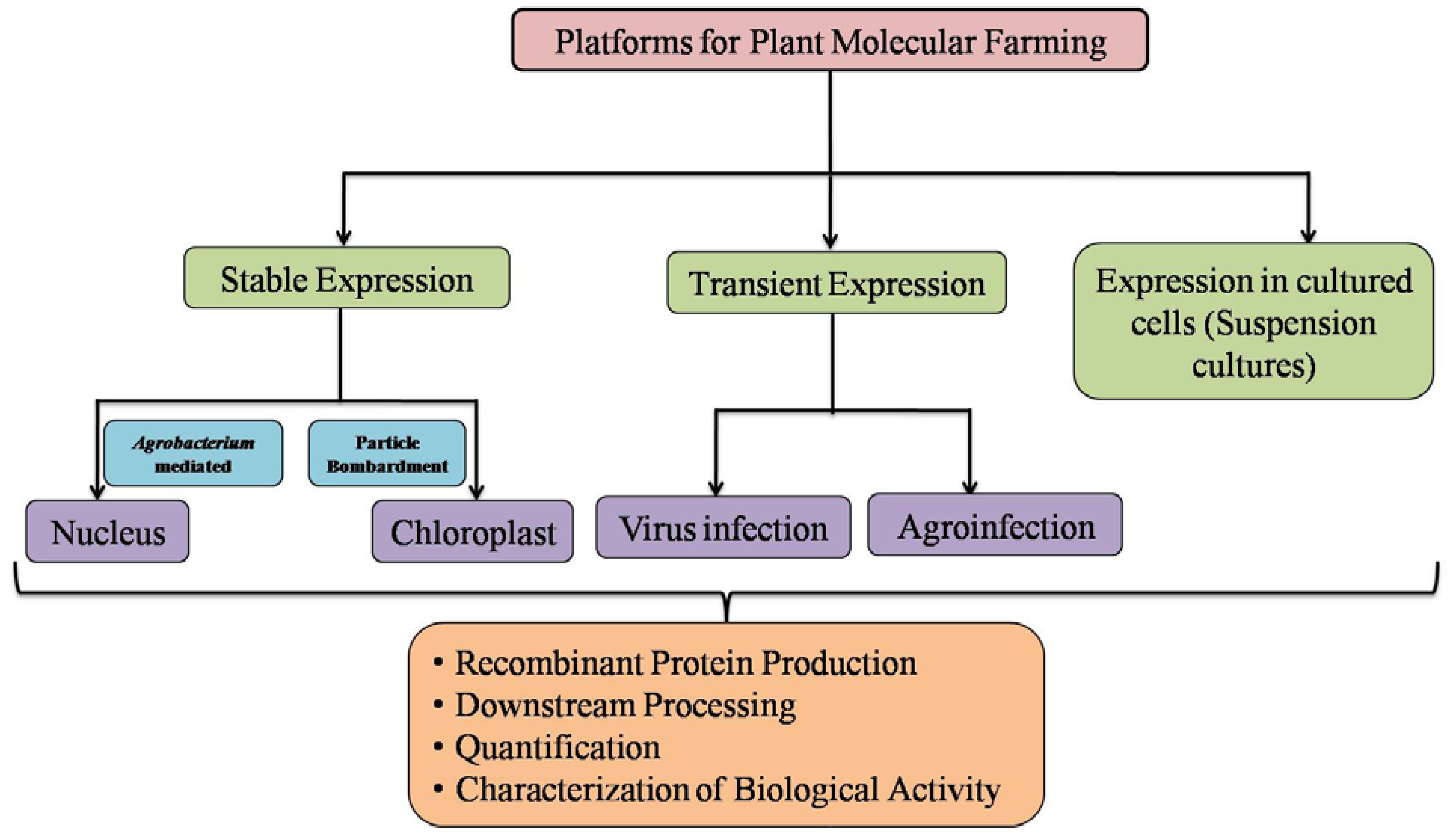{kind=link}
{kind=link}
| Expression System | Advantages | Disadvantages |
|---|---|---|
| Bacteria | Easy to manipulate Low cost High expression Ease of scale up Short turnaround time Established regulatory procedures and approval | Improper folding Lack of post-translational modifications which may affect the protein function Endotoxin accumulation |
| Mammalian Cells | Proper folding and authentic post-translational modifications Existing regulatory approval | High production cost Expensive media and culture condition requirements |
| Yeast | Rapid growth and scalable Easy to manipulate Simple and inexpensive media requirements and culture conditions Post-translational modifications of recombinant proteins | Difficulty in cell disruption due to the thick and hard cell walls Hyperglycosylation of proteins Limited glycosylation capacity |
| Insect cells | High expression levels Ability to produce complex proteins including secreted, membrane, and intracellular proteins Proper folding and post-translational modifications | High cost and time consuming Expensive media and culture condition requirements |
| Plant | Rapid and affordable Optimized growth conditions Free from pathogen and bacterial toxin contaminants Economical Post-translational modification somewhat similar like mammalian system | Regulatory compliance Limited glycosylation capacity |
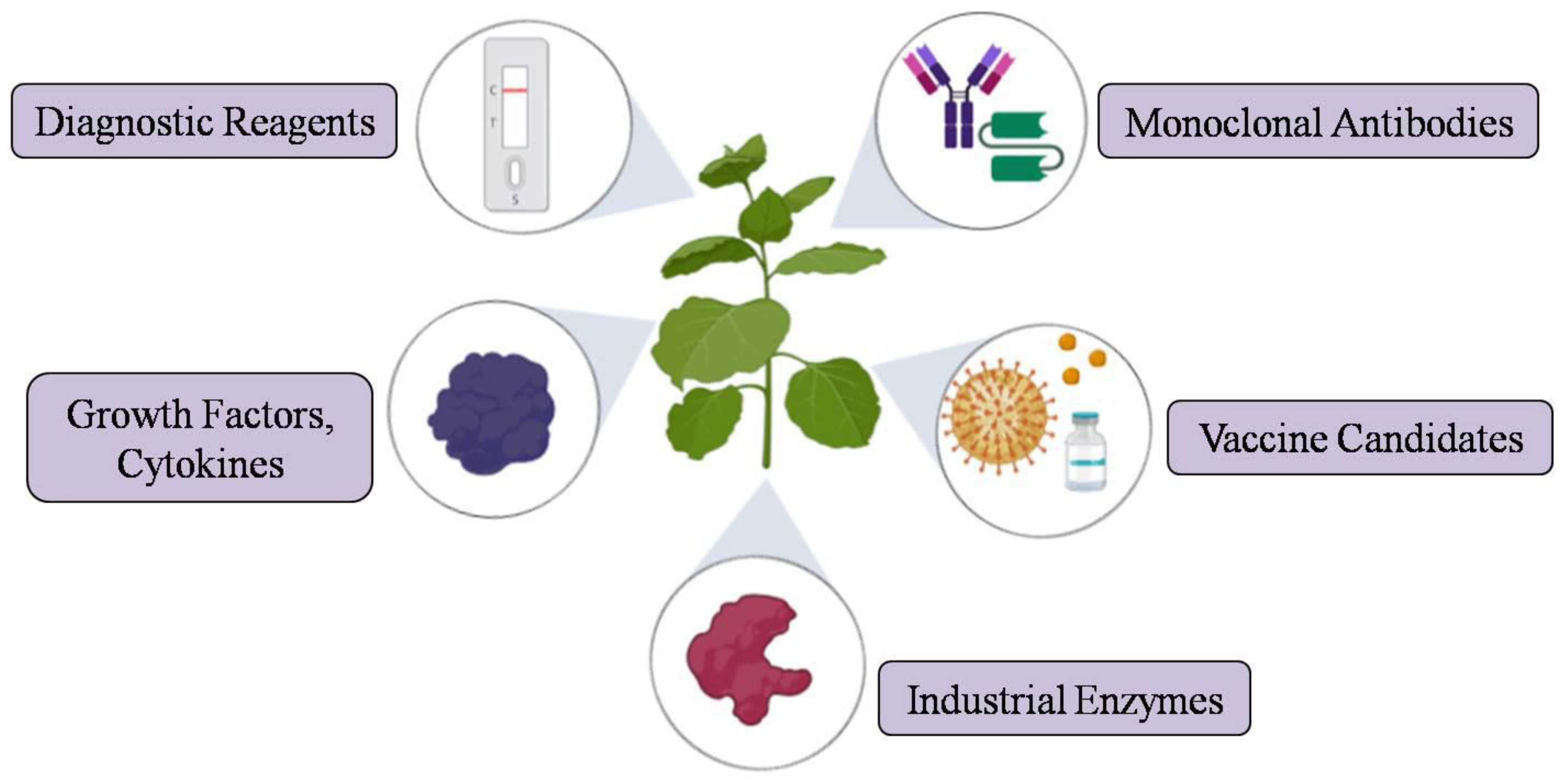
3. Plant-Derived Recombinant Proteins
| Vaccine Candidates | |||||
|---|---|---|---|---|---|
| Recombinant Protein | Pathogen/Disease | Expression System | Transformation Method | Expression Level | Reference |
| Hepatitis B surface antigen | Hepatitis B virus | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 66 ng/mg of soluble protein | [38] |
| Structural protein VP60 | Rabbit hemorrhagic disease virus (RHDV) | Potato (Solanum tuberosum) | Agrobacterium mediated (Stable expression/Nucleus) | 0.3% of total soluble protein | [39] |
| Spike (S) protein of transmissible gastroenteritis virus | Transmissible gastroenteritis virus (TGEV) | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 0.1–0.2% of total soluble protein | [40] |
| Hemagglutinin protein of rinderpest virus | Rinderpest virus (RPV) | Peanut (Arachis hypogea L.) | Agrobacterium mediated (Stable expression/Nucleus) | 0.2–1.3% of total soluble protein | [41] |
| Glycoprotein D (gD) of bovine herpes virus | Bovine herpes virus | Tobacco (Nicotiana benthamiana) | Mechanical inoculation (Stable expression/Nucleus) | 20 μg/g fresh weight (FW) | [42] |
| L1 major capsid protein | Human papillomavirus | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 2–4 µg/kg FW | [43] |
| Spike (S) protein of transmissible gastroenteritis virus | Transmissible gastroenteritis virus (TGEV) | Corn (Zea mays) | Agrobacterium mediated (Stable expression/Nucleus) | 13 mg/kg FW | [44] |
| Spike (S) protein of infectious bronchitis virus | Infectious bronchitis virus (IBV) | Potato (Solanum tuberosum) | Agrobacterium mediated (Stable expression/Nucleus) | 2.39–2.53 µg/g FW | [26] |
| Anthrax protective antigen (PA) | Anthrax | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 14.2% of total soluble protein | [45] |
| Hepatitis B virus surface antigen | Hepatitis B virus (HBV) | Potato (Solanum tuberosum) | Agrobacterium mediated (Stable expression/Nucleus) | 8.5 μg/g FW | [46] |
| Fusion (F) protein of Newcastle disease virus | Newcastle disease virus (NDV) | Corn (Zea mays L.) | Biolistic method (Stable expression/Chloroplast) | 3.0% of total soluble protein | [47] |
| F4 fimbrial adhesin FaeG | Enterotoxigenic E. coli | Alfalfa (Medicago sativa L.) | Agrobacterium mediated (Stable expression/Chloroplast) | 1.0% of total soluble protein | [48] |
| L1 capsid protein gene | Cottontail rabbit papillomavirus | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 0.4–1 mg/kg of total leaf mass | [49] |
| Structural protein VP2 | Infectious bursal disease virus (IBDV) | Rice | Agrobacterium mediated (Stable expression/Nucleus) | 40.21 µg/g FW | [50] |
| Hepatitis B virus surface antigen | Hepatitis B virus (HBV) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 295 µg/g FW | [51] |
| Haemagglutinin (HA) | H5N1 (avian influenza virus) & H1N1 (human) influenza strains | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 50 mg/kg FW | [52] |
| Heat-labile toxin B subunit (LTB) | Enterotoxigenic E. coli | Carrot (Daucus carota) | Agrobacterium mediated (Stable expression/Nucleus) | 0.3% of total soluble protein | [53] |
| Norwalk virus capsid protein | Norwalk virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.8 mg/g FW | [54] |
| Structural protein VP1 | Foot-and-mouth disease virus (FMDV) | Legume (Stylosanthes guianensis) | Agrobacterium mediated (Stable expression/Nucleus) | 0.1–0.5% of total soluble protein | [55] |
| HIV-1 Pr55gag Polyprotein | Human immunodeficiency virus type 1 (HIV) | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 312–363 mg/kg FW | [56] |
| Japanese encephalitis virus (JEV) envelope protein (E) | Japanese encephalitis virus | Japonica rice (Nipponbare) | Agrobacterium mediated (Stable expression/Nucleus) | 1.1–1.9 μg/mg of total soluble protein | [57] |
| Hemagglutinin (HA) | Avian influenza (H5N1) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.3 g/kg FW | [58] |
| Haemagglutinin (HA) | Influenza virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 400–1300 mg/kg FW | [59] |
| Haemagglutinin (HA) | Avian influenza A (H7N7) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.2 g/kg FW | [60] |
| Structural protein VP2 | Infectious bursal disease virus (IBDV) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 1.0% of total soluble protein | [61] |
| Structural protein E2 | Bovine viral diarrhea virus (BVDV) | Alfalfa (Medicago sativa L.) | Agrobacterium mediated (Stable expression/Nucleus) | 1 μg/g FW | [62] |
| Bluetongue virus-like particles | Bluetongue virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 70 mg/kg FW | [63] |
| Narita 104 virus virus-like particles | Narita 104 virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.3 mg/g FW | [64] |
| Glycoprotein (GP) of PRRSV | Porcine reproductive and respiratory syndrome virus (PRRSV) | Arabidopsis thaliana | Agrobacterium mediated (Stable expression/Nucleus) | 2.74% of total soluble protein | [65] |
| Matrix protein 2 ectodomain (M2e) | Avian influenza (H5N1) | Duckweed (Lemna minor) | Agrobacterium mediated (Stable expression/Nucleus) | 90–970 mg/kg FW | [66] |
| Matrix protein 2 ectodomain (M2e) | Avian influenza (H5N1) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 125–205 mg/kg FW | [67] |
| Consensus domain III of dengue virus E glycoprotein (cEDIII) | Dengue virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 5.2 mg/g FW | [68] |
| Dengue envelop protein domain III (EDIII) | Dengue virus | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 0.8–1.6% of total soluble protein | [69] |
| PV3 VLPs | Poliovirus (PV) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 60 mg/kg FW | [28] |
| E. maxima gametocyte antigen (Gam82) | Eimeria maxima | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Transient expression) | 20 mg/kg FW | [70] |
| CHIKV E1 and E2 | Chikungunya virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 8–13 mg/kg of fresh leaf weight | [71] |
| ZIKV envelope (E) protein | Zika virus (ZIKV) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 160 μg/g FW | [72] |
| Porcine circovirus type 2 (PCV-2) capsid protein | Porcine circovirus type 2 | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 6.5 mg/kg leaf wet weight | [73] |
| HIV Env gp140 | Human immunodeficiency virus (HIV) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 5–6 mg/kg FW | [74] |
| H6 subtype haemagglutinin (HA) | Influenza A virus (H6N2) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 95 mg/kg FW | [31] |
| Antibodies | |||||
| cT84.66 | Cancer (tumor marker) | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Transient expression) | 1 mg/kg FW | [75] |
| scFvT84.66 | Cancer (tumor marker) | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Transient expression) | 5 mg/kg FW | [75] |
| scFvT84.66 | Cancer (tumor marker) | Rice (Oryza sativa) | Biolistic method (Stable expression/Nucleus) | 3.8 μg/g FW | [76] |
| scFvT84.66 | Cancer (tumor marker) | Cereal crops (wheat and rice) | Biolistic method (Stable expression/Nucleus) | 30 μg/g FW | [77] |
| BR55-2 | Human colorectal cancer | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 30 mg/kg FW | [78] |
| 2F5 | HIV | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Stable expression/Nucleus) | 0.01% of total soluble protein | [79] |
| 2G12 | HIV | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.3 g/kg FW | [80] |
| 2G12 | HIV | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 8 mg/L culture medium | [81] |
| 6D8 | Ebola virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.5 mg/g FW | [82] |
| 6D8 | Ebola virus | Lettuce (L. sativa) | Agrobacterium mediated (Transient expression) | 0.23–0.27 mg/g FW | [83] |
| CO17-1AK | Human colorectal cancer | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 0.25 mg/kg FW | [84] |
| Palivizumab-N | Respiratory syncytial virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 180 mg/kg FW | [85] |
| E559 | Rabies | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 1.8 mg/kg FW (0.04% of total soluble protein) | [86] |
| pE16 | West Nile virus | Tobacco (Nicotiana benthamiana ∆XF) | Agrobacterium mediated (Transient expression) | 0.74 mg/g FW | [87] |
| pE16scFv-CH | West Nile virus | Tobacco (Nicotiana benthamiana ∆XF) | Agrobacterium mediated (Transient expression) | 0.77 mg/g FW | [87] |
| E60 | Dengue virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 120 μg/g FW | [88] |
| 2G12 | HIV | Rice (Oryza sativa) | Biolistic method (Stable expression/Nucleus) | 46.4 μg/g dry seed weight | [89] |
| 8B10 and 5F10 | Chikungunya virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 20–30 mg/kg FW | [71] |
| SO57 | Rabies virus | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Transient expression) | 0.014–0.019% of total soluble protein | [90] |
| cD5 | Enterovirus 71 | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 50 μg/g FW | [91] |
| PD1 | Cancer | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 140 μg/g FW | [92] |
| c2A10G6 | Zika virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 1.47 mg/g FW | [93] |
| 6D8 | Ebola | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 1.21 mg/g FW | [93] |
| HSV8 | Herpes simplex virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 1.42 mg/g FW | [93] |
| CHKV mab | Chikungunya virus | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 100 μg/g FW | [94] |
| Recombinant Proteins | Expression System | Transformation Method | Expression Level | Reference |
|---|---|---|---|---|
| Human serum albumin | Potato (Solanum tuberosum) | Agrobacterium mediated (Stable expression/Nucleus) | 0.25 μg/mg (0.02% of total soluble protein) | [95] |
| Erythropoietin | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 26 pg/mg total protein | [96] |
| α1-antitrypsin | Rice (Japonica rice) | Biolistic method (Stable expression/Nucleus) | 4.6–5.7 mg/g dry cell | [97] |
| Aprotinin | Corn | Biolistic method (Stable expression/Nucleus) | 0.069% of total extractable seed protein | [98] |
| Human-secreted alkaline phosphatase | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 1.1 μg/g FW (3% of total soluble protein) | [99] |
| Collagen | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 0.03 g/kg powdered plants | [100] |
| Human somatotropin (hST) | Tobacco | Biolistic method (Stable expression/Chloroplast) | >7% of total soluble protein | [101] |
| Bacillus thuringiensis (Bt) cry2Aa2 | Tobacco | Biolistic method (Stable expression/Chloroplast) | 5 mg/g FW (45.3–46.1% of total soluble protein) | [102] |
| Human serum albumin | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 11.1% of total protein | [103] |
| Human epidermal growth factor (hEGF) | Tobacco (Nicotiana tabacum) | Agrobacterium mediated (Stable expression/Nucleus) | 34.2 µg/g FW | [104] |
| Human basic fibroblast growth factor (bFGF) | Soybean (Glycine max) | Cotyledonary node explant method (Stable expression/Nucleus) | 2.3% of total soluble protein | [105] |
| Type I interferon (IFNα2b) | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 3 mg/g FW (20% of total soluble protein) | [106] |
| Human growth hormone (hGH) | Rice (Oryza sativa) | Biolistic method (Stable expression/Nucleus) | 57 mg/L culture medium | [107] |
| PlyGBS lysin | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | >70% of the total soluble protein | [108] |
| Human growth hormone (hGH) | Tobacco BY-2 cells | Agrobacterium mediated (Stable expression/Nucleus) | 35 mg/L culture medium (2-4% of total soluble protein) | [109] |
| Human basic fibroblast growth factor (bFGF) | Rice (Oryza sativa) | Agrobacterium mediated (Stable expression/Nucleus) | 185.66 mg/kg | [110] |
| Lumbrokinase | Sunflower (Helianthus annuus L.) | Agrobacterium mediated (Stable expression/Nucleus) | 5.1 g/kg seeds | [111] |
| Human acidic fibroblast growth factor 1 (FGF-1) | Salvia miltiorrhiza | Agrobacterium mediated (Stable expression/Nucleus) | 272 ng/g FW | [112] |
| Glucocerebrosidase (GCase) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Stable expression/Nucleus) | 68 μg/g FW (1.45% of total soluble protein) | [113] |
| Human acid alpha glucosidase | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 6.38 μg/g FW | [114] |
| Human basic fibroblast growth factor (bFGF) | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 0.1% of total soluble protein | [115] |
| Endo-β-1,4-xylanase | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | 35.7% of total soluble protein | [116] |
| β-Glucosidase | Tobacco (Nicotiana tabacum) | Biolistic method (Stable expression/Chloroplast) | >75% of total soluble protein | [116] |
| Osteopontin (OPN) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 100 ng/g FW | [117] |
| Dentin matrix protein-1 (DMP1) | Tobacco (Nicotiana benthamiana) | Agrobacterium mediated (Transient expression) | 0.3 µg/g FW | [118] |
4. Strategies Used for Recombinant Protein Production in Plants
5. Perspectives
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| cGMP | Current good manufacturing practice |
| COVID-19 | Coronavirus disease |
| FW | Fresh weight |
| mAb | Monoclonal antibody |
| MERS-CoV | Middle East respiratory syndrome coronavirus |
| PMF | Plant molecular farming |
| PTM | Post-translational modification |
| SARS-CoV | Severe acute respiratory syndrome coronavirus |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
References
- Burnett, M.J.B.; Burnett, A.C. Therapeutic recombinant protein production in plants: Challenges and opportunities. Plants People Planet 2020, 2, 121–132. [Google Scholar] [CrossRef]
- Schillberg, S.; Raven, N.; Spiegel, H.; Rasche, S.; Buntru, M. Critical analysis of the commercial potential of plants for the production of recombinant proteins. Front. Plant Sci. 2019, 10, 720. [Google Scholar] [CrossRef] [PubMed]
- Vieira Gomes, A.M.; Souza Carmo, T.; Silva Carvalho, L.; Mendonca Bahia, F.; Parachin, N.S. Comparison of yeasts as hosts for recombinant protein production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Lalonde, M.E.; Durocher, Y. Therapeutic glycoprotein production in mammalian cells. J. Biotechnol. 2017, 251, 128–140. [Google Scholar] [CrossRef]
- Sainsbury, F.; Lomonossoff, G.P. Transient expressions of synthetic biology in plants. Curr. Opin. Plant Biol. 2014, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Shanmugaraj, B.; Ramalingam, S. Plant expression platform for the production of recombinant pharmaceutical proteins. Austin J. Biotechnol. Bioeng. 2014, 1, 4. [Google Scholar]
- Hiatt, A.; Cafferkey, R.; Bowdish, K. Production of antibodies in transgenic plants. Nature 1989, 342, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.L. First plant-made biologic approved. Nat. Biotechnol. 2012, 30, 472. [Google Scholar] [CrossRef]
- Tschofen, M.; Knopp, D.; Hood, E.; Stöger, E. Plant molecular farming: Much more than medicines. Annu. Rev. Anal. Chem. 2016, 9, 271–294. [Google Scholar] [CrossRef]
- Daniell, H.; Singh, N.D.; Mason, H.; Streatfield, S.J. Plant-made vaccine antigens and biopharmaceuticals. Trends Plant Sci. 2009, 14, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Lomonossoff, G.P.; D’Aoust, M.A. Plant-produced biopharmaceuticals: A case of technical developments driving clinical deployment. Science 2016, 353, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Park, K.Y.; Wi, S.J. Potential of plants to produce recombinant protein products. J. Plant Biol. 2016, 59, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Topp, E.; Irwin, R.; McAllister, T.; Lessard, M.; Joensuu, J.J.; Kolotilin, I.; Conrad, U.; Stoger, E.; Mor, T.; Warzecha, H.; et al. The case for plant-made veterinary immunotherapeutics. Biotechnol. Adv. 2016, 34, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Sohrab, S.S.; Suhail, M.; Kamal, M.A.; Husen, A.; Azhar, E.I. Recent development and future prospects of plant-based vaccines. Curr. Drug Metab. 2017, 18, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Wong-Arce, A.; Gonzalez-Ortega, O.; Rosales-Mendoza, S. Plant-made vaccines in the fight against cancer. Trends Biotechnol. 2017, 35, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Taunt, H.N.; Stoffels, L.; Purton, S. Green biologics: The algal chloroplast as a platform for making biopharmaceuticals. Bioengineered 2018, 9, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Donini, M.; Marusic, C. Current state-of-the-art in plant-based antibody production systems. Biotechnol. Lett. 2019, 41, 335–346. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant molecular farming of virus-like nanoparticles as vaccines and reagents. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2020, 12, e1587. [Google Scholar] [CrossRef]
- Chen, Q.; Davis, K.R. The potential of plants as a system for the development and production of human biologics. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Sahdev, S.; Khattar, S.K.; Saini, K.S. Production of active eukaryotic proteins through bacterial expression systems: A review of the existing biotechnology strategies. Mol. Cell. Biochem. 2008, 307, 249–264. [Google Scholar] [CrossRef]
- Strasser, R.; Stadlmann, J.; Schahs, M.; Stiegler, G.; Quendler, H.; Mach, L.; Glossl, J.; Weterings, K.; Pabst, M.; Steinkellner, H. Generation of glyco-engineered Nicotiana benthamiana for the production of monoclonal antibodies with a homogeneous human-like N-glycan structure. Plant Biotechnol. J. 2008, 6, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Montero-Morales, L.; Steinkellner, H. Advanced plant-based glycan engineering. Front. Bioeng. Biotechnol. 2018, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Ma, J.K. Plant-made pharmaceuticals: Leading products and production platforms. Biotechnol. Appl. Biochem. 2011, 58, 58–67. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Dickey, A.; Wang, K.Y. Plants as factories for human pharmaceuticals: Applications and challenges. Int. J. Mol. Sci. 2015, 16, 28549–28565. [Google Scholar] [CrossRef] [PubMed]
- Shanmugaraj, B.; Malla, A.; Phoolcharoen, W. Emergence of novel coronavirus 2019-nCoV: Need for rapid vaccine and biologics development. Pathogens 2020, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.Y.; Cheng, L.Q.; Zheng, X.J.; Wu, J.X.; Shang, S.B.; Wang, J.Y.; Chen, J.G. Generation of the transgenic potato expressing full-length spike protein of infectious bronchitis virus. J. Biotechnol. 2004, 111, 121–130. [Google Scholar] [CrossRef]
- Daniell, H.; Rai, V.; Xiao, Y. Cold chain and virus-free oral polio booster vaccine made in lettuce chloroplasts confers protection against all three poliovirus serotypes. Plant Biotechnol. J. 2019, 17, 1357–1368. [Google Scholar] [CrossRef]
- Marsian, J.; Fox, H.; Bahar, M.W.; Kotecha, A.; Fry, E.E.; Stuart, D.I.; Macadam, A.J.; Rowlands, D.J.; Lomonossoff, G.P. Plant-made polio type 3 stabilized VLPs-a candidate synthetic polio vaccine. Nat. Commun. 2017, 8, 245. [Google Scholar] [CrossRef]
- Diamos, A.G.; Mason, H.S. High-level expression and enrichment of norovirus virus-like particles in plants using modified geminiviral vectors. Protein Expr. Purif. 2018, 151, 86–92. [Google Scholar] [CrossRef]
- Veerapen, V.P.; van Zyl, A.R.; Wigdorovitz, A.; Rybicki, E.P.; Meyers, A.E. Novel expression of immunogenic foot-and-mouth disease virus-like particles in Nicotiana benthamiana. Virus Res. 2018, 244, 213–217. [Google Scholar] [CrossRef]
- Smith, T.; O’Kennedy, M.M.; Wandrag, D.B.R.; Adeyemi, M.; Abolnik, C. Efficacy of a plant-produced virus-like particle vaccine in chickens challenged with Influenza A H6N2 virus. Plant Biotechnol. J. 2020, 18, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Scotti, N.; Rybicki, E.P. Virus-like particles produced in plants as potential vaccines. Expert Rev. Vaccines 2013, 12, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P. Plant-based vaccines against viruses. Virol. J. 2014, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Hood, E.E.; Witcher, D.R.; Maddock, S.; Meyer, T.; Baszczynski, C.; Bailey, M.; Flynn, P.; Register, J.; Marshall, L.; Bond, D.; et al. Commercial production of avidin from transgenic maize: Characterization of transformant, production, processing, extraction and purification. Mol. Breed. 1997, 3, 291–306. [Google Scholar] [CrossRef]
- Witcher, D.R.; Hood, E.E.; Peterson, D.; Bailey, M.; Bond, D.; Kusnadi, A.; Evangelista, R.; Nikolov, Z.; Wooge, C.; Mehigh, R.; et al. Commercial production of β-glucuronidase (GUS): A model system for the production of proteins in plants. Mol. Breed. 1998, 4, 301–312. [Google Scholar] [CrossRef]
- Woodard, S.L.; Mayor, J.M.; Bailey, M.R.; Barker, D.K.; Love, R.T.; Lane, J.R.; Delaney, D.E.; McComas-Wagner, J.M.; Mallubhotla, H.D.; Hood, E.E.; et al. Maize (Zea mays)-derived bovine trypsin: Characterization of the first large-scale, commercial protein product from transgenic plants. Biotechnol. Appl. Biochem. 2003, 38, 123–130. [Google Scholar] [CrossRef]
- Ma, J.K.-C.; Drossard, J.; Lewis, D.; Altmann, F.; Boyle, J.; Christou, P.; Cole, T.; Dale, P.; van Dolleweerd, C.J.; Isitt, V.; et al. Regulatory approval and a first-in-human phase I clinical trial of a monoclonal antibody produced in transgenic tobacco plants. Plant Biotechnol. J. 2015, 13, 1106–1120. [Google Scholar] [CrossRef]
- Mason, H.S.; Lam, D.M.; Arntzen, C.J. Expression of hepatitis B surface antigen in transgenic plants. Proc. Natl. Acad. Sci. USA 1992, 89, 11745. [Google Scholar] [CrossRef] [Green Version]
- Castanon, S.; Marin, M.S.; Martin-Alonso, J.M.; Boga, J.A.; Casais, R.; Humara, J.M.; Ordas, R.J.; Parra, F. Immunization with potato plants expressing VP60 protein protects against rabbit hemorrhagic disease virus. J. Virol. 1999, 73, 4452–4455. [Google Scholar] [CrossRef] [Green Version]
- Tuboly, T.; Yu, W.; Bailey, A.; Degrandis, S.; Du, S.; Erickson, L.; Nagy, E. Immunogenicity of porcine transmissible gastroenteritis virus spike protein expressed in plants. Vaccine 2000, 18, 2023–2028. [Google Scholar] [CrossRef]
- Khandelwal, A.; Lakshmi Sita, G.; Shaila, M.S. Oral immunization of cattle with hemagglutinin protein of rinderpest virus expressed in transgenic peanut induces specific immune responses. Vaccine 2003, 21, 3282–3289. [Google Scholar] [CrossRef]
- Pérez Filgueira, D.M.; Zamorano, P.I.; Domínguez, M.G.; Taboga, O.; Del Médico Zajac, M.P.; Puntel, M.; Romera, S.A.; Morris, T.J.; Borca, M.V.; Sadir, A.M. Bovine herpes virus gD protein produced in plants using a recombinant tobacco mosaic virus (TMV) vector possesses authentic antigenicity. Vaccine 2003, 21, 4201–4209. [Google Scholar] [CrossRef] [Green Version]
- Varsani, A.; Williamson, A.L.; Rose, R.C.; Jaffer, M.; Rybicki, E.P. Expression of Human papillomavirus type 16 major capsid protein in transgenic Nicotiana tabacum cv. Xanthi. Arch. Virol. 2003, 148, 1771–1786. [Google Scholar] [CrossRef]
- Lamphear, B.J.; Jilka, J.M.; Kesl, L.; Welter, M.; Howard, J.A.; Streatfield, S.J. A corn-based delivery system for animal vaccines: An oral transmissible gastroenteritis virus vaccine boosts lactogenic immunity in swine. Vaccine 2004, 22, 2420–2424. [Google Scholar] [CrossRef]
- Koya, V.; Moayeri, M.; Leppla, S.H.; Daniell, H. Plant-based vaccine: Mice immunized with chloroplast-derived anthrax protective antigen survive anthrax lethal toxin challenge. Infect. Immun. 2005, 73, 8266–8274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanavala, Y.; Mahoney, M.; Pal, S.; Scott, A.; Richter, L.; Natarajan, N.; Goodwin, P.; Arntzen, C.J.; Mason, H.S. Immunogenicity in humans of an edible vaccine for hepatitis B. Proc. Natl. Acad. Sci. USA 2005, 102, 3378–3382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Andrade, O.; Loza-Rubio, E.; Olivera-Flores, T.; Fehervari-Bone, T.; Gomez-Lim, M.A. Expression of the Newcastle disease virus fusion protein in transgenic maize and immunological studies. Transgenic Res. 2006, 15, 455–463. [Google Scholar] [CrossRef]
- Joensuu, J.J.; Verdonck, F.; Ehrstrom, A.; Peltola, M.; Siljander-Rasi, H.; Nuutila, A.M.; Oksman-Caldentey, K.M.; Teeri, T.H.; Cox, E.; Goddeeris, B.M.; et al. F4 (K88) fimbrial adhesin FaeG expressed in alfalfa reduces F4+ enterotoxigenic Escherichia coli excretion in weaned piglets. Vaccine 2006, 24, 2387–2394. [Google Scholar] [CrossRef]
- Kohl, T.; Hitzeroth, I.I.; Stewart, D.; Varsani, A.; Govan, V.A.; Christensen, N.D.; Williamson, A.L.; Rybicki, E.P. Plant-produced cottontail rabbit papillomavirus L1 protein protects against tumor challenge: A proof-of-concept study. Clin. Vaccine Immunol. 2006, 13, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Yu, L.; Li, L.; Hu, J.; Zhou, J.; Zhou, X. Oral immunization with transgenic rice seeds expressing VP2 protein of infectious bursal disease virus induces protective immune responses in chickens. Plant Biotechnol. J. 2007, 5, 570–578. [Google Scholar] [CrossRef]
- Huang, Z.; LePore, K.; Elkin, G.; Thanavala, Y.; Mason, H.S. High-yield rapid production of hepatitis B surface antigen in plant leaf by a viral expression system. Plant Biotechnol. J. 2008, 6, 202–209. [Google Scholar] [CrossRef]
- D’Aoust, M.-A.; Lavoie, P.-O.; Couture, M.M.-J.; Trépanier, S.; Guay, J.-M.; Dargis, M.; Mongrand, S.; Landry, N.; Ward, B.J.; Vézina, L.-P. Influenza virus-like particles produced by transient expression in Nicotiana benthamiana induce a protective immune response against a lethal viral challenge in mice. Plant Biotechnol. J. 2008, 6, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; Soria-Guerra, R.E.; Lopez-Revilla, R.; Moreno-Fierros, L.; Alpuche-Solis, A.G. Ingestion of transgenic carrots expressing the Escherichia coli heat-labile enterotoxin B subunit protects mice against cholera toxin challenge. Plant Cell Rep. 2008, 27, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Santi, L.; Batchelor, L.; Huang, Z.; Hjelm, B.; Kilbourne, J.; Arntzen, C.J.; Chen, Q.; Mason, H.S. An efficient plant viral expression system generating orally immunogenic Norwalk virus-like particles. Vaccine 2008, 26, 1846–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.M.; Zhu, J.B.; Peng, M.; Zhou, P. Induction of a protective antibody response to FMDV in mice following oral immunization with transgenic Stylosanthes spp. as a feedstuff additive. Transgenic Res. 2008, 17, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Scotti, N.; Alagna, F.; Ferraiolo, E.; Formisano, G.; Sannino, L.; Buonaguro, L.; De Stradis, A.; Vitale, A.; Monti, L.; Grillo, S.; et al. High-level expression of the HIV-1 Pr55gag polyprotein in transgenic tobacco chloroplasts. Planta 2009, 229, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, H.; Zhang, X.; Xiao, H.; Jiang, Y.; Song, Y.; Fang, L.; Xiao, S.; Zhen, Y.; Chen, H. Generation and immunogenicity of Japanese encephalitis virus envelope protein expressed in transgenic rice. Biochem. Biophys. Res. Commun. 2009, 380, 292–297. [Google Scholar] [CrossRef]
- Kalthoff, D.; Giritch, A.; Geisler, K.; Bettmann, U.; Klimyuk, V.; Hehnen, H.R.; Gleba, Y.; Beer, M. Immunization with plant-expressed hemagglutinin protects chickens from lethal highly pathogenic avian influenza virus H5N1 challenge infection. J. Virol. 2010, 84, 12002–12010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, Y.; Farrance, C.E.; Bautista, J.; Bi, H.; Musiychuk, K.; Horsey, A.; Park, H.; Jaje, J.; Green, B.J.; Shamloul, M.; et al. A plant-based system for rapid production of influenza vaccine antigens. Influenza Other Respir. Viruses 2012, 6, 204–210. [Google Scholar] [CrossRef]
- Kanagarajan, S.; Tolf, C.; Lundgren, A.; Waldenstrom, J.; Brodelius, P.E. Transient expression of hemagglutinin antigen from low pathogenic avian influenza A (H7N7) in Nicotiana benthamiana. PLoS ONE 2012, 7, e33010. [Google Scholar] [CrossRef] [Green Version]
- Gómez, E.; Lucero, M.S.; Chimeno Zoth, S.; Carballeda, J.M.; Gravisaco, M.J.; Berinstein, A. Transient expression of VP2 in Nicotiana benthamiana and its use as a plant-based vaccine against Infectious Bursal Disease Virus. Vaccine 2013, 31, 2623–2627. [Google Scholar] [CrossRef] [PubMed]
- Perez Aguirreburualde, M.S.; Gomez, M.C.; Ostachuk, A.; Wolman, F.; Albanesi, G.; Pecora, A.; Odeon, A.; Ardila, F.; Escribano, J.M.; Dus Santos, M.J.; et al. Efficacy of a BVDV subunit vaccine produced in alfalfa transgenic plants. Vet. Immunol. Immunopathol. 2013, 151, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Thuenemann, E.C.; Meyers, A.E.; Verwey, J.; Rybicki, E.P.; Lomonossoff, G.P. A method for rapid production of heteromultimeric protein complexes in plants: Assembly of protective bluetongue virus-like particles. Plant Biotechnol. J. 2013, 11, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.G.; Herbst-Kralovetz, M.M.; Mason, H.S. Norovirus Narita 104 virus-like particles expressed in Nicotiana benthamiana induce serum and mucosal immune responses. Biomed. Res. Int. 2014, 2014, 807539. [Google Scholar] [CrossRef]
- Piron, R.; De Koker, S.; De Paepe, A.; Goossens, J.; Grooten, J.; Nauwynck, H.; Depicker, A. Boosting in planta production of antigens derived from the porcine reproductive and respiratory syndrome virus (PRRSV) and subsequent evaluation of their immunogenicity. PLoS ONE 2014, 9, e91386. [Google Scholar] [CrossRef]
- Firsov, A.; Tarasenko, I.; Mitiouchkina, T.; Ismailova, N.; Shaloiko, L.; Vainstein, A.; Dolgov, S. High-yield expression of M2e peptide of Avian Influenza virus H5N1 in transgenic duckweed plants. Mol. Biotechnol. 2015, 57, 653–661. [Google Scholar] [CrossRef]
- Mbewana, S.; Mortimer, E.; Pera, F.F.; Hitzeroth, I.I.; Rybicki, E.P. Production of H5N1 influenza virus matrix protein 2 ectodomain protein bodies in tobacco plants and in insect cells as a candidate universal influenza vaccine. Front. Bioeng. Biotechnol. 2015, 3, 197. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Jang, Y.-S.; Yang, M.-S.; Kim, T.-G. High expression of consensus dengue virus envelope glycoprotein domain III using a viral expression system in tobacco. Plant Cell Tissue Organ Cult. 2015, 122, 445–451. [Google Scholar] [CrossRef]
- Gottschamel, J.; Lössl, A.; Ruf, S.; Wang, Y.; Skaugen, M.; Bock, R.; Clarke, J.L. Production of dengue virus envelope protein domain III-based antigens in tobacco chloroplasts using inducible and constitutive expression systems. Plant Mol. Biol. 2016, 91, 497–512. [Google Scholar] [CrossRef] [Green Version]
- Kota, S.; Subramanian, M.; Shanmugaraj, B.; Challa, H.; Ponanna, N.M. Subunit vaccine based on plant expressed recombinant Eimeria gametocyte antigen Gam82 elicit protective immune response against chicken coccidiosis. J. Vaccines Vaccin 2017, 8. [Google Scholar] [CrossRef]
- Iyappan, G.; Shanmugaraj, B.M.; Inchakalody, V.; Ma, J.K.C.; Ramalingam, S. Potential of plant biologics to tackle the epidemic like situations—Case studies involving viral and bacterial candidates. Int. J. Infect. Dis. 2018, 73, 363. [Google Scholar] [CrossRef]
- Yang, M.; Sun, H.; Lai, H.; Hurtado, J.; Chen, Q. Plant-produced Zika virus envelope protein elicits neutralizing immune responses that correlate with protective immunity against Zika virus in mice. Plant Biotechnol. J. 2018, 16, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Gunter, C.J.; Regnard, G.L.; Rybicki, E.P.; Hitzeroth, I.I. Immunogenicity of plant-produced porcine circovirus-like particles in mice. Plant Biotechnol. J. 2019, 17, 1751–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolin, E.; Chapman, R.; Meyers, A.E.; van Diepen, M.T.; Ximba, P.; Hermanus, T.; Crowther, C.; Weber, B.; Morris, L.; Williamson, A.L.; et al. Production and immunogenicity of soluble plant-produced HIV-1 subtype C envelope gp140 immunogens. Front. Plant Sci. 2019, 10, 1378. [Google Scholar] [CrossRef]
- Vaquero, C.; Sack, M.; Chandler, J.; Drossard, J.; Schuster, F.; Monecke, M.; Schillberg, S.; Fischer, R. Transient expression of a tumor-specific single-chain fragment and a chimeric antibody in tobacco leaves. Proc. Natl. Acad. Sci. USA 1999, 96, 11128–11133. [Google Scholar] [CrossRef] [Green Version]
- Torres, E.; Vaquero, C.; Nicholson, L.; Sack, M.; Stöger, E.; Drossard, J.; Christou, P.; Fischer, R.; Perrin, Y. Rice cell culture as an alternative production system for functional diagnostic and therapeutic antibodies. Transgenic Res. 1999, 8, 441–449. [Google Scholar] [CrossRef]
- Stöger, E.; Vaquero, C.; Torres, E.; Sack, M.; Nicholson, L.; Drossard, J.; Williams, S.; Keen, D.; Perrin, Y.; Christou, P.; et al. Cereal crops as viable production and storage systems for pharmaceutical scFv antibodies. Plant Mol. Biol. 2000, 42, 583–590. [Google Scholar] [CrossRef]
- Brodzik, R.; Glogowska, M.; Bandurska, K.; Okulicz, M.; Deka, D.; Ko, K.; van der Linden, J.; Leusen, J.H.; Pogrebnyak, N.; Golovkin, M.; et al. Plant-derived anti-Lewis Y mAb exhibits biological activities for efficient immunotherapy against human cancer cells. Proc. Natl. Acad. Sci. USA 2006, 103, 8804–8809. [Google Scholar] [CrossRef] [Green Version]
- Floss, D.M.; Sack, M.; Stadlmann, J.; Rademacher, T.; Scheller, J.; Stöger, E.; Fischer, R.; Conrad, U. Biochemical and functional characterization of anti-HIV antibody–ELP fusion proteins from transgenic plants. Plant Biotechnol. J. 2008, 6, 379–391. [Google Scholar] [CrossRef]
- Sainsbury, F.; Lomonossoff, G.P. Extremely high-level and rapid transient protein production in plants without the use of viral replication. Plant Physiol. 2008, 148, 1212–1218. [Google Scholar] [CrossRef] [Green Version]
- Holland, T.; Sack, M.; Rademacher, T.; Schmale, K.; Altmann, F.; Stadlmann, J.; Fischer, R.; Hellwig, S. Optimal nitrogen supply as a key to increased and sustained production of a monoclonal full-size antibody in BY-2 suspension culture. Biotechnol. Bioeng. 2010, 107, 278–289. [Google Scholar] [CrossRef]
- Huang, Z.; Phoolcharoen, W.; Lai, H.; Piensook, K.; Cardineau, G.; Zeitlin, L.; Whaley, K.J.; Arntzen, C.J.; Mason, H.S.; Chen, Q. High-level rapid production of full-size monoclonal antibodies in plants by a single-vector DNA replicon system. Biotechnol. Bioeng. 2010, 106, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, H.; He, J.; Engle, M.; Diamond, M.S.; Chen, Q. Robust production of virus-like particles and monoclonal antibodies with geminiviral replicon vectors in lettuce. Plant Biotechnol. J. 2012, 10, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, Y.; Lee, K.-J.; Kim, D.-S.; Lee, J.-H.; Oh, D.-B.; Hwang, K.-A.; Ko, K.; Choo, Y.-K.; Ko, K. Glycomodification and characterization of anti-colorectal cancer immunotherapeutic monoclonal antibodies in transgenic tobacco. Plant Cell Tissue Organ Cult. 2013, 113, 41–49. [Google Scholar] [CrossRef]
- Zeitlin, L.; Bohorov, O.; Bohorova, N.; Hiatt, A.; Kim, D.H.; Pauly, M.H.; Velasco, J.; Whaley, K.J.; Barnard, D.L.; Bates, J.T.; et al. Prophylactic and therapeutic testing of Nicotiana-derived RSV-neutralizing human monoclonal antibodies in the cotton rat model. MAbs 2013, 5, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dolleweerd, C.J.; Teh, A.Y.; Banyard, A.C.; Both, L.; Lotter-Stark, H.C.; Tsekoa, T.; Phahladira, B.; Shumba, W.; Chakauya, E.; Sabeta, C.T.; et al. Engineering, expression in transgenic plants and characterisation of E559, a rabies virus-neutralising monoclonal antibody. J. Infect. Dis. 2014, 210, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Lai, H.; He, J.; Hurtado, J.; Stahnke, J.; Fuchs, A.; Mehlhop, E.; Gorlatov, S.; Loos, A.; Diamond, M.S.; Chen, Q. Structural and functional characterization of an anti-West Nile virus monoclonal antibody and its single-chain variant produced in glycoengineered plants. Plant Biotechnol. J. 2014, 12, 1098–1107. [Google Scholar] [CrossRef] [Green Version]
- Dent, M.; Hurtado, J.; Paul, A.M.; Sun, H.; Lai, H.; Yang, M.; Esqueda, A.; Bai, F.; Steinkellner, H.; Chen, Q. Plant-produced anti-dengue virus monoclonal antibodies exhibit reduced antibody-dependent enhancement of infection activity. J. Gen. Virol. 2016, 97, 3280–3290. [Google Scholar] [CrossRef] [Green Version]
- Vamvaka, E.; Twyman, R.M.; Murad, A.M.; Melnik, S.; Teh, A.Y.-H.; Arcalis, E.; Altmann, F.; Stoger, E.; Rech, E.; Ma, J.K.C.; et al. Rice endosperm produces an underglycosylated and potent form of the HIV-neutralizing monoclonal antibody 2G12. Plant Biotechnol. J. 2016, 14, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Shafaghi, M.; Maktoobian, S.; Rasouli, R.; Howaizi, N.; Ofoghi, H.; Ehsani, P. Transient expression of biologically active anti-rabies virus monoclonal antibody in tobacco leaves. Iran. J. Biotechnol. 2018, 16, e1774. [Google Scholar] [CrossRef]
- Rattanapisit, K.; Chao, Z.; Siriwattananon, K.; Huang, Z.; Phoolcharoen, W. Plant-produced anti-enterovirus 71 (EV71) monoclonal antibody efficiently protects mice against EV71 infection. Plants 2019, 8, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattanapisit, K.; Phakham, T.; Buranapraditkun, S.; Siriwattananon, K.; Boonkrai, C.; Pisitkun, T.; Hirankarn, N.; Strasser, R.; Abe, Y.; Phoolcharoen, W. Structural and in vitro functional analyses of novel plant-produced anti-human PD1 antibody. Sci. Rep. 2019, 9, 15205. [Google Scholar] [CrossRef] [PubMed]
- Diamos, A.G.; Hunter, J.G.L.; Pardhe, M.D.; Rosenthal, S.H.; Sun, H.; Foster, B.C.; DiPalma, M.P.; Chen, Q.; Mason, H.S. High level production of monoclonal antibodies using an optimized plant expression system. Front. Bioeng. Biotechnol. 2019, 7, 472. [Google Scholar] [CrossRef]
- Hurtado, J.; Acharya, D.; Lai, H.; Sun, H.; Kallolimath, S.; Steinkellner, H.; Bai, F.; Chen, Q. In vitro and in vivo efficacy of anti-chikungunya virus monoclonal antibodies produced in wild-type and glycoengineered Nicotiana benthamiana plants. Plant Biotechnol. J. 2020, 18, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Sijmons, P.C.; Dekker, B.M.; Schrammeijer, B.; Verwoerd, T.C.; van den Elzen, P.J.; Hoekema, A. Production of correctly processed human serum albumin in transgenic plants. Biotechnology 1990, 8, 217–221. [Google Scholar] [CrossRef]
- Matsumoto, S.; Ikura, K.; Ueda, M.; Sasaki, R. Characterization of a human glycoprotein (erythropoietin) produced in cultured tobacco cells. Plant Mol. Biol. 1995, 27, 1163–1172. [Google Scholar] [CrossRef]
- Terashima, M.; Murai, Y.; Kawamura, M.; Nakanishi, S.; Stoltz, T.; Chen, L.; Drohan, W.; Rodriguez, R.L.; Katoh, S. Production of functional human α1-antitrypsin by plant cell culture. Appl. Microbiol. Biotechnol. 1999, 52, 516–523. [Google Scholar] [CrossRef]
- Zhong, G.-Y.; Peterson, D.; Delaney, D.E.; Bailey, M.; Witcher, D.R.; Register Iii, J.C.; Bond, D.; Li, C.-P.; Marshall, L.; Kulisek, E.; et al. Commercial production of aprotinin in transgenic maize seeds. Mol. Breed. 1999, 5, 345–356. [Google Scholar] [CrossRef]
- Komarnytsky, S.; Borisjuk, N.V.; Borisjuk, L.G.; Alam, M.Z.; Raskin, I. Production of recombinant proteins in tobacco guttation fluid. Plant Physiol. 2000, 124, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, F.; Exposito, J.Y.; Bournat, P.; Gruber, V.; Perret, S.; Comte, J.; Olagnier, B.; Garrone, R.; Theisen, M. Triple helix assembly and processing of human collagen produced in transgenic tobacco plants. FEBS Lett. 2000, 469, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Staub, J.M.; Garcia, B.; Graves, J.; Hajdukiewicz, P.T.; Hunter, P.; Nehra, N.; Paradkar, V.; Schlittler, M.; Carroll, J.A.; Spatola, L.; et al. High-yield production of a human therapeutic protein in tobacco chloroplasts. Nat. Biotechnol. 2000, 18, 333–338. [Google Scholar] [CrossRef]
- De Cosa, B.; Moar, W.; Lee, S.B.; Miller, M.; Daniell, H. Overexpression of the Bt cry2Aa2 operon in chloroplasts leads to formation of insecticidal crystals. Nat. Biotechnol. 2001, 19, 71–74. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Mingo-Castel, A.; Miller, M.; Daniell, H. A chloroplast transgenic approach to hyper-express and purify Human Serum Albumin, a protein highly susceptible to proteolytic degradation. Plant Biotechnol. J. 2003, 1, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Wirth, S.; Calamante, G.; Mentaberry, A.; Bussmann, L.; Lattanzi, M.; Barañao, L.; Bravo-Almonacid, F. Expression of active human epidermal growth factor (hEGF) in tobacco plants by integrative and non-integrative systems. Mol. Breed. 2004, 13, 23–35. [Google Scholar] [CrossRef]
- Ding, S.H.; Huang, L.Y.; Wang, Y.D.; Sun, H.C.; Xiang, Z.H. High-level expression of basic fibroblast growth factor in transgenic soybean seeds and characterization of its biological activity. Biotechnol. Lett. 2006, 28, 869–875. [Google Scholar] [CrossRef]
- Arlen, P.A.; Falconer, R.; Cherukumilli, S.; Cole, A.; Cole, A.M.; Oishi, K.K.; Daniell, H. Field production and functional evaluation of chloroplast-derived interferon-alpha2b. Plant Biotechnol. J. 2007, 5, 511–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.G.; Baek, M.Y.; Lee, E.K.; Kwon, T.H.; Yang, M.S. Expression of human growth hormone in transgenic rice cell suspension culture. Plant Cell Rep. 2008, 27, 885–891. [Google Scholar] [CrossRef]
- Oey, M.; Lohse, M.; Kreikemeyer, B.; Bock, R. Exhaustion of the chloroplast protein synthesis capacity by massive expression of a highly stable protein antibiotic. Plant J. 2009, 57, 436–445. [Google Scholar] [CrossRef]
- Xu, J.; Okada, S.; Tan, L.; Goodrum, K.J.; Kopchick, J.J.; Kieliszewski, M.J. Human growth hormone expressed in tobacco cells as an arabinogalactan-protein fusion glycoprotein has a prolonged serum life. Transgenic Res. 2010, 19, 849–867. [Google Scholar] [CrossRef]
- An, N.; Ou, J.; Jiang, D.; Zhang, L.; Liu, J.; Fu, K.; Dai, Y.; Yang, D. Expression of a functional recombinant human basic fibroblast growth factor from transgenic rice seeds. Int. J. Mol. Sci. 2013, 14, 3556–3567. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; Du, X.; Wang, G.; Ji, J.; Jin, C.; Li, X. Expression of biologically active anti-thrombosis protein lumbrokinase in edible sunflower seed kernel. J. Plant Biochem. Biotechnol. 2014, 23, 257–265. [Google Scholar] [CrossRef]
- Tan, Y.; Wang, K.Y.; Wang, N.; Li, G.; Liu, D. Ectopic expression of human acidic fibroblast growth factor 1 in the medicinal plant, Salvia miltiorrhiza, accelerates the healing of burn wounds. BMC Biotechnol. 2014, 14, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limkul, J.; Misaki, R.; Kato, K.; Fujiyama, K. The combination of plant translational enhancers and terminator increase the expression of human glucocerebrosidase in Nicotiana benthamiana plants. Plant Sci. 2015, 240, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Sherman, A.; Doerfler, P.A.; Byrne, B.J.; Herzog, R.W.; Daniell, H. Oral delivery of Acid Alpha Glucosidase epitopes expressed in plant chloroplasts suppresses antibody formation in treatment of Pompe mice. Plant Biotechnol. J. 2015, 13, 1023–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.P.; Wei, Z.Y.; Zhong, X.F.; Lin, C.J.; Cai, Y.H.; Ma, J.; Zhang, Y.Y.; Liu, Y.Z.; Xing, S.C. Stable expression of basic fibroblast growth factor in chloroplasts of tobacco. Int. J. Mol. Sci. 2016, 17, 19. [Google Scholar] [CrossRef] [Green Version]
- Castiglia, D.; Sannino, L.; Marcolongo, L.; Ionata, E.; Tamburino, R.; De Stradis, A.; Cobucci-Ponzano, B.; Moracci, M.; La Cara, F.; Scotti, N. High-level expression of thermostable cellulolytic enzymes in tobacco transplastomic plants and their use in hydrolysis of an industrially pretreated Arundo donax L. biomass. Biotechnol. Biofuels 2016, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Rattanapisit, K.; Abdulheem, S.; Chaikeawkaew, D.; Kubera, A.; Mason, H.S.; Ma, J.K.; Pavasant, P.; Phoolcharoen, W. Recombinant human osteopontin expressed in Nicotiana benthamiana stimulates osteogenesis related genes in human periodontal ligament cells. Sci. Rep. 2017, 7, 17358. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.R.; Kaewpungsup, P.; Khorattanakulchai, N.; Rattanapisit, K.; Pavasant, P.; Phoolcharoen, W. Recombinant human dentin matrix protein 1 (hDMP1) expressed in Nicotiana benthamiana potentially induces osteogenic differentiation. Plants 2019, 8, 566. [Google Scholar] [CrossRef] [Green Version]
- Menary, J.; Hobbs, M.; Mesquita de Albuquerque, S.; Pacho, A.; Drake, P.M.W.; Prendiville, A.; Ma, J.K.; Fuller, S.S. Shotguns vs Lasers: Identifying barriers and facilitators to scaling-up plant molecular farming for high-value health products. PLoS ONE 2020, 15, e0229952. [Google Scholar] [CrossRef]
- Twyman, R.M.; Stoger, E.; Schillberg, S.; Christou, P.; Fischer, R. Molecular farming in plants: Host systems and expression technology. Trends Biotechnol. 2003, 21, 570–578. [Google Scholar] [CrossRef]
- Yusibov, V.; Rabindran, S. Recent progress in the development of plant derived vaccines. Expert Rev. Vaccines 2008, 7, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Viral vectors for the expression of proteins in plants. Curr. Opin. Biotechnol. 2007, 18, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Wong, G.; Audet, J.; Bello, A.; Fernando, L.; Alimonti, J.B.; Fausther-Bovendo, H.; Wei, H.; Aviles, J.; Hiatt, E.; et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 2014, 514, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales-Mendoza, S. Will plant-made biopharmaceuticals play a role in the fight against COVID-19? Expert Opin. Biol. Ther. 2020, 20, 545–548. [Google Scholar] [CrossRef] [Green Version]
- Shanmugaraj, B.; Siriwattananon, K.; Wangkanont, K.; Phoolcharoen, W. Perspectives on monoclonal antibody therapy as potential therapeutic intervention for Coronavirus disease-19 (COVID-19). Asian Pac. J. Allergy Immunol. 2020, 38, 10–18. [Google Scholar]
- Capell, T.; Twyman, R.M.; Armario-Najera, V.; Ma, J.K.C.; Schillberg, S.; Christou, P. Potential applications of plant biotechnology against SARS-CoV-2. Trends Plant Sci. 2020, 25, 635–643. [Google Scholar] [CrossRef]
- Buyel, J.F. Plant molecular farming—Integration and exploitation of side streams to achieve sustainable biomanufacturing. Front. Plant Sci. 2018, 9, 1893. [Google Scholar] [CrossRef] [PubMed]
- Maliga, P. Engineering the plastid genome of higher plants. Curr. Opin. Plant Biol. 2002, 5, 164–172. [Google Scholar] [CrossRef]
- Bock, R. Plastid biotechnology: Prospects for herbicide and insect resistance, metabolic engineering and molecular farming. Curr. Opin. Biotechnol. 2007, 18, 100–106. [Google Scholar] [CrossRef]
- Jin, S.; Daniell, H. The engineered chloroplast genome just got smarter. Trends Plant Sci. 2015, 20, 622–640. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Shanmugaraj, B.; Daniell, H. Expression and functional evaluation of biopharmaceuticals made in plant chloroplasts. Curr. Opin. Chem. Biol. 2017, 38, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, N. On the way to commercializing plant cell culture platform for biopharmaceuticals: Present status and prospect. Pharm. Bioprocess. 2014, 2, 499–518. [Google Scholar] [CrossRef] [Green Version]
- Tekoah, Y.; Shulman, A.; Kizhner, T.; Ruderfer, I.; Fux, L.; Nataf, Y.; Bartfeld, D.; Ariel, T.; Gingis–Velitski, S.; Hanania, U.; et al. Large-scale production of pharmaceutical proteins in plant cell culture—The protalix experience. Plant Biotechnol. J. 2015, 13, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.B.; Abranches, R.; Fischer, R.; Sack, M.; Holland, T. Putting the spotlight back on plant suspension cultures. Front. Plant Sci. 2016, 7, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurusamy, P.D.; Schafer, H.; Ramamoorthy, S.; Wink, M. Biologically active recombinant human erythropoietin expressed in hairy root cultures and regenerated plantlets of Nicotiana tabacum L. PLoS ONE 2017, 12, e0182367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy root cultures—A versatile tool with multiple applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shanmugaraj, B.; I. Bulaon, C.J.; Phoolcharoen, W. Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production. Plants 2020, 9, 842. https://doi.org/10.3390/plants9070842
Shanmugaraj B, I. Bulaon CJ, Phoolcharoen W. Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production. Plants. 2020; 9(7):842. https://doi.org/10.3390/plants9070842
Chicago/Turabian StyleShanmugaraj, Balamurugan, Christine Joy I. Bulaon, and Waranyoo Phoolcharoen. 2020. "Plant Molecular Farming: A Viable Platform for Recombinant Biopharmaceutical Production" Plants 9, no. 7: 842. https://doi.org/10.3390/plants9070842