The Effects of Postharvest Treatments and Sunlight Exposure on the Reproductive Capability and Viability of Phyllosticta citricarpa in Citrus Black Spot Fruit Lesions

 , ,
, ,
Abstract
:1. Introduction
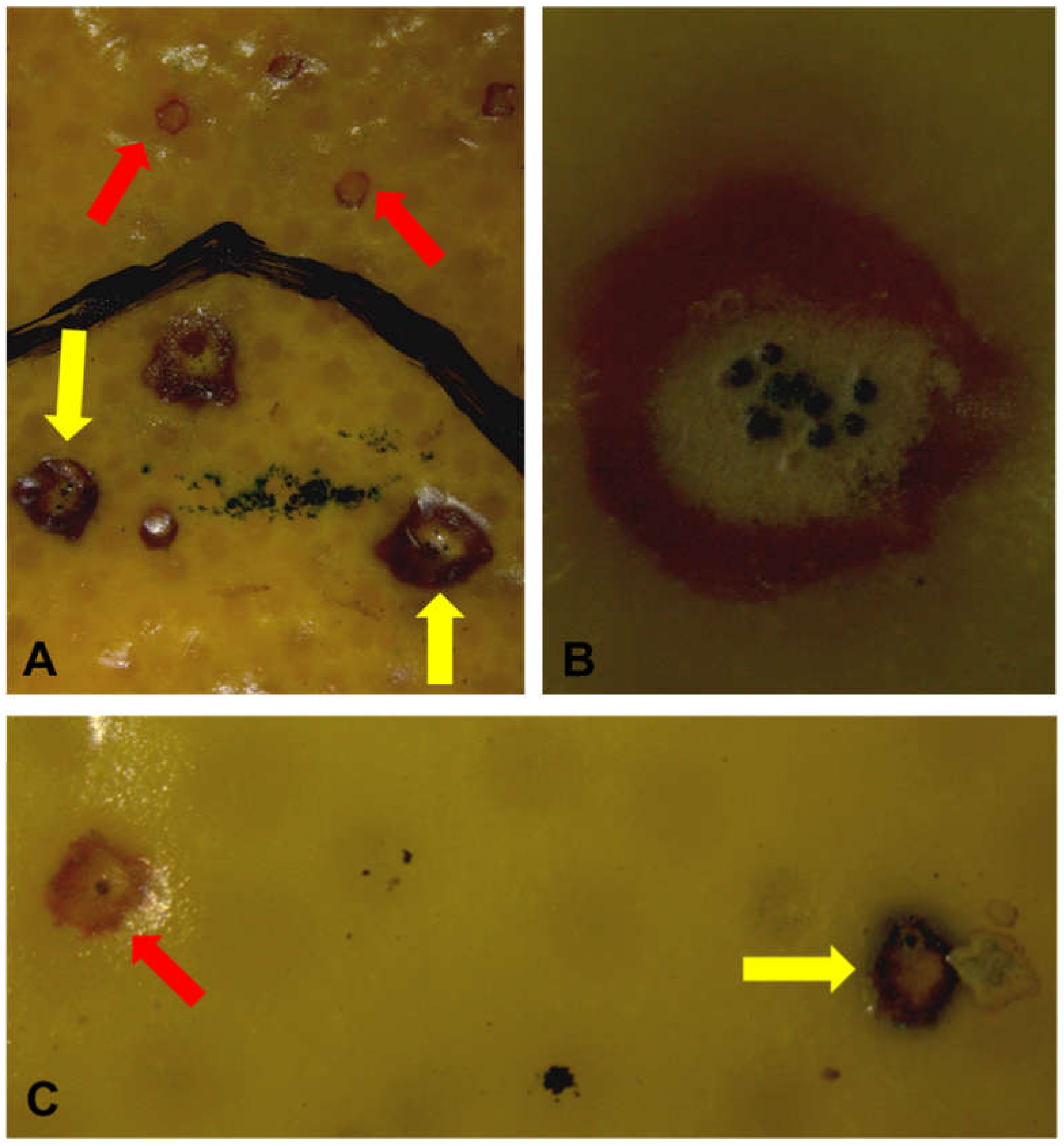
2. Results
2.1. Effects of Treatments on New CBS Lesion Development
2.2. Pycnidiospore Release and Viability after Sun Exposure of CBS Lesions
2.2.1. Whole Fruit and Peel Segments
2.2.2. Citrus Pulp
2.3. Verification of CBS Lesion Diagnosis
3. Discussion
4. Materials and Methods
4.1. Fruit Harvesting, Treatments, and Incubation
4.2. Sunlight Exposure of Whole Fruit and Peel Segments
4.3. Sunlight Exposure of Citrus Pulp
4.4. Pycnidiospore Release and Viability of CBS Lesions
4.5. Verification of CBS Lesion Diagnosis
4.6. Experimental Layout and Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CBS Expert Panel. Response to EFSA Panel on Plant Health, 2013—Draft Scientific Opinion on the Risk of Phyllosticta citricarpa (Guignardia citricarpa) for the EU Territory with Identification and Evaluation of Risk Reduction Options. 2013. Available online: http://www.citrusres.com/market-access (accessed on 27 November 2017).
- USDA-APHIS. Risk Assessment of Citrus spp. Fruit as a Pathway for the Introduction of Guignardia citricarpa Kiely, the Organism That Causes Citrus Black Spot Disease; Center for Plant Health Science and Technology, Plant Epidemiology and Risk Analysis Laboratory: Raleigh, NC, USA, 2010. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health). Scientific Opinion on the risk of Phyllosticta citricarpa (Guignardia citricarpa) for the EU territory with identification and evaluation of risk reduction options. EFSA J. 2014, 12, 243. [Google Scholar]
- Guarnaccia, V.; Gehrmann, T.; Silva-Junior, G.J.; Fourie, P.H.; Haridas, S.; Vu, D.; Spatafora, J.; Martin, F.M.; Robert, V.; Grigoriev, I.V.; et al. Phyllosticta citricarpa and sister species of global importance to Citrus. Mol. Plant Pathol. 2019, 20, 1619–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotzé, J.M. Epidemiology and control of citrus black spot in South Africa. Plant Dis. 1981, 65, 945–950. [Google Scholar] [CrossRef]
- Fourie, P.; Schutte, T.; Serfontein, S.; Swart, F. Modeling the effect of temperature and wetness on Guignardia pseudothecium maturation and ascospore release in citrus orchards. Phytopathology 2013, 103, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Moyo, P.; Du Raan, S.; Fourie, P.H. Models for predicting pseudothecium maturity and ascospore release of Phyllosticta spp. in South African citrus orchards. S. Afr. J. Sci. 2020, 116, 7955. [Google Scholar] [CrossRef]
- Kiely, T.B. Preliminary studies on Guignardia citricarpa (n. sp.), the ascigerous stage of Phoma citricarpa McAlp., and its relation to blackspot of citrus. Proc. Linn. Soc. N. S. W. 1948, 73, 249–292. [Google Scholar]
- Spósito, M.B.; Amorim, L.; Bassanezi, R.B.; Bergamin Filho, A.; Hau, B. Spatial pattern of black spot incidence within citrus trees related to disease severity and pathogen dispersal. Plant Pathol. 2008, 57, 103–108. [Google Scholar] [CrossRef]
- Sposito, M.B.; Amorim, L.; Bassanezi, R.B.; Yamamoto, P.; Felippe, M.R.; Czermainski, A.B.C. Relative importance of inoculum sources of Guignardia citricarpa on the citrus black spot epidemic in Brazil. Crop Prot. 2011, 30, 1546–1552. [Google Scholar] [CrossRef] [Green Version]
- Wager, V.A. The black spot disease of citrus in South Africa. Sci. Bull. 1952, 303, 1–52. [Google Scholar]
- McOnie, K.C. Source of infection for black spot of citrus. S. Afr. Citrus J. 1965, 378, 5–9. [Google Scholar]
- Hendricks, K.E.; Christman, M.; Roberts, P.D. Spatial and temporal patterns of commercial citrus trees affected by Phyllosticta citricarpa in Florida. Sci. Rep. 2017, 1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, K.E.; Christman, M.C.; Roberts, P.D. The effect of weather and location of fruit within the tree on the incidence and severity of citrus black spot on Fruit. Sci. Rep. 2020, 1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstens, E.; Linde, C.C.; Slabbert, R.; Miles, A.K.; Donovan, N.J.; Hongye, L.; Dewdney, M.M.; Glienke, C.; Schutte, G.C.; Fourie, P.H.; et al. A global perspective on the population structure and reprodcutive system of Phyllosticta citricarpa, a pathogen of citrus. Phytopathology 2017, 107, 758–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstens, E.; Linde, C.C.; Fourie, P.H.; Bester-van der Merwe, A.E.; Langenhoven, S.D.; McLeod, A. Spatial and temporal genetic analyses of Phyllosticta citricarpa in two lemon orchards in South Africa reveal a role of asexual reproduction within sexually reproducing populations. Phytopathology 2020, in press. [Google Scholar] [CrossRef]
- McOnie, K.C. Germination and infection of citrus by ascospores of Guignardia citricarpa in relation to control of black spot. Phytopathology 1967, 57, 743–746. [Google Scholar]
- Darnell-Smith, G.P. An account of some observations upon the life-history of Phoma citricarpa McAlph., the cause of the black spot disease in citrus fruit in New South Wales. Proc. Linn. Soc. N. S. W. 1918, 43, 868–881. [Google Scholar]
- Kotzé, J.M. Black spot. In Compendium of Citrus Diseases; Timmer, L.W., Garnsey, S.M., Graham, J.H., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2000; pp. 23–25. [Google Scholar]
- Brentu, F.C.; Oduro, K.A.; Offei, S.K.; Odamtten, G.T.; Vicent, A.; Peres, N.A.; Timmer, L.W. Crop loss, aetiology, and epidemiology of citrus black spot in Ghana. Eur. J. Plant Pathol. 2012, 133, 657–670. [Google Scholar] [CrossRef]
- Marques, J.P.R.; Spósito, M.B.; Mello, A.F.S.; Amorim, L.; Mondin, M.; Appezzato-da-Gloria, B. Histopathology of black spot symptoms in sweet oranges. Eur. J. Plant Pathol. 2012, 133, 439–448. [Google Scholar] [CrossRef]
- FAO. ISPM No 27. Diagnostic Protocols for Regulated Pests Intentional Standards for Phytosanitary Measures, Publication No 27: Annex 5. Phyllosticta Citricarpa (Mcalpine) AA on Fruit; Secretariat of the International Plant Protection, Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Lourenço, S.A.; Gasparoto, M.C.G.; Spósito, M.B.; Amorim, L.; Gottwald, T.R. Infectious period of citrus black spot on sweet orange. In Proceedings of the XII International Citrus Congress, Valencia, Spain, 18–23 November 2012. [Google Scholar]
- Schutte, G.C.; Kotze, C.; Korf, H.J.G. Influence of sunlight exposure on fertility of Phyllosticta citricarpa pycnidia in citrus black spot lesions on grapefruit and Valencia orange rinds. SA Fruit J. 2014, 13, 54–57. [Google Scholar]
- Korf, H.J.G. Survival of Phyllosticta citricarpa, Anamorph of the Citrus Black Spot Pathogen. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 1998. [Google Scholar]
- Korf, H.J.G.; Schuttle, G.C.; Kotzé, J.M. Effect of packinghouse procedures of the viability of Phyllosticta citricarpa, anamorph of the black spot pathogen. Afr. Plant Protect. 2001, 7, 103–109. [Google Scholar]
- Schreuder, W.; du Plooy, W.; Erasmus, A.; Savage, C.; Basson, E.; Lennox, C.; Fourie, P.H. Postharvest fungicide treatments and cold storage control citrus black spot infections. Crop Prot. 2018, 112, 332–342. [Google Scholar] [CrossRef]
- Erasmus, A.; Lennox, C.; Jordaan, H.; Smilanick, J.; Lesar, K.; Fourie, P.H. Imazalil residue loading and green mould control in citrus packhouses. Postharvest Biol. Technol. 2011, 62, 193–203. [Google Scholar] [CrossRef]
- Njombolwana, N.S.; Erasmus, A.; Fourie, P.H. Evaluation of curative and protective control of Penicillium digitatum following imazalil application in wax coating. Postharvest Biol. Technol. 2013, 77, 102–110. [Google Scholar] [CrossRef]
- Seberry, J.A.; Leggo, D.; Kiely, T.B. Effect of skin coatings on the development of black spot in stored Valencia Oranges. Aust. J. Exp. Agric. Anim. Husb. 1967, 7, 593–600. [Google Scholar] [CrossRef]
- Agostini, J.; Peres, N.; Mackenzie, S.; Adaskaveg, J.; Timmer, L. Effect of fungicides and storage conditions on postharvest development of citrus black spot and survival of Guignardia citricarpa in fruit tissues. Plant Dis. 2006, 90, 1419–1424. [Google Scholar] [CrossRef] [Green Version]
- Lanfranchi, M. Economic analysis on the enhancement of citrus waste for energy production. J. Essent. Oil Res. 2012, 24, 583–591. [Google Scholar] [CrossRef]
- Zema, D.A.; Calabro, P.S.; Folino, A.; Tamburino, V.; Zappia, G.; Zimbone, S.M. Valorisation of citrus processing waste: A review. Waste Manag. 2017, 80, 252–273. [Google Scholar] [CrossRef]
- Wang, N.Y.; Dewdney, M.M. The effect of relative humidity on colonization of citrus twigs by Phyllosticta citricarpa, the citrus black spot pathogen. Phytopathology 2015, 105, 145–146. [Google Scholar]
- Brodrick, H.T.; Rabie, C.J. Light and temperature effects on symptom development and sporulation of Guignardia citricarpa Kiley, on Citrus sinensis (Linn) Osbeck. Phytophylactica 1970, 2, 157–164. [Google Scholar]
- Baldassari, R.B.; Brandimarte, I.; Gustavo de Andrade, A.; Goncalves de Souza, D.C.; Moretto, C.; De Goes, A. Induction of the precoce expression of Guignardia citricarpa symptoms in fruits of Pera-Rio sweet orange. Rev. Bras. Frutic. 2007, 29, 269–275. [Google Scholar] [CrossRef]
- Truter, M. Epidemiology of Citrus Black Spot Disease in South Africa and Its Impact on Phytosanitary Trade Restrictions. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2010. [Google Scholar]
- Hu, J.; Johnson, E.G.; Wang, N.Y.; Davoglio, T.; Dewdney, M.M. qPCR quantification of pathogenic Guignardia citricarpa and nonpathogenic G. mangiferae in citrus. Plant Dis. 2014, 98, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Blanca, M.J.; Alarcón, R.; Arnau, J.; Bono, R.; Bendayan, R. Non-normal data: Is ANOVA still a valid option? Psicothema 2017, 29, 552–557. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Treatments | Mean Number of New CBS Lesions per Fruit | New Lesions with Pycnidia (%) | |
|---|---|---|---|
| 2019 | 2020 | ||
| ‘Eureka’ lemons * | |||
| Untreated fruit | 17.60 a | 7.90 b | 6.61 a |
| Treated fruit | 2.10 c | 1.30 c | 1.40 b |
| ‘Midknight’ Valencia oranges ** | |||
| Untreated fruit | 14.21 a | ̶ | 4.43 a |
| Treated fruit | 2.33 b | ̶ | 0.77 b |
| Sun-Exposure Period (Hours) | ‘Eureka’ Lemon | ‘Midknight’ Valencia Oranges | ||
|---|---|---|---|---|
| Untreated | Treated | Untreated | Treated | |
| 0 | 100.0 (0.000) a | 0.0 (0.000) c | 84.0 (3.685) a | 0.0 (0.000) c |
| 2 | 82.8 (5.003) b | 0.0 (0.000) c | 82.0 (3.861) a | 0.0 (0.000) c |
| 4 | 81.0 (5.193) b | 0.0 (0.000) c | 59.0 (4.943) b | 0.0 (0.000) c |
| 6 | 74.1 (5.800) b | 0.0 (0.000) c | 58.0 (4.960) b | 0.0 (0.000) c |
| ‘Eureka’ Lemon | ‘Midknight’ Valencia Oranges | |||
|---|---|---|---|---|
| Sun-Exposure Period (Days) | Untreated | Treated | Untreated | Treated |
| 0 | 54.3 (4.225) a | 4.3 (1.718) c | 19.3 (3.346) a | 8.6 (2.374) b |
| 1 | 21.4 (3.480) b | 3.6 (1.574) c | 17.9 (3.249) a | 7.9 (2.282) b |
| 2 | 8.6 (2.374) c | 1.4 (1.007) c | 2.9 (1.413) b | 1.4 (1.007) b |
| 3 | 3.6 (1.574) c | 0.0 (0.000) c | 0.0 (0.000) b | 0.0 (0.000) b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyo, P.; Fourie, P.H.; Masikane, S.L.; de Oliveira Fialho, R.; Mamba, L.C.; du Plooy, W.; Hattingh, V. The Effects of Postharvest Treatments and Sunlight Exposure on the Reproductive Capability and Viability of Phyllosticta citricarpa in Citrus Black Spot Fruit Lesions. Plants 2020, 9, 1813. https://doi.org/10.3390/plants9121813
Moyo P, Fourie PH, Masikane SL, de Oliveira Fialho R, Mamba LC, du Plooy W, Hattingh V. The Effects of Postharvest Treatments and Sunlight Exposure on the Reproductive Capability and Viability of Phyllosticta citricarpa in Citrus Black Spot Fruit Lesions. Plants. 2020; 9(12):1813. https://doi.org/10.3390/plants9121813
Chicago/Turabian StyleMoyo, Providence, Paul H. Fourie, Siyethemba L. Masikane, Régis de Oliveira Fialho, Lindokuhle C. Mamba, Wilma du Plooy, and Vaughan Hattingh. 2020. "The Effects of Postharvest Treatments and Sunlight Exposure on the Reproductive Capability and Viability of Phyllosticta citricarpa in Citrus Black Spot Fruit Lesions" Plants 9, no. 12: 1813. https://doi.org/10.3390/plants9121813