Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants


 ,
,
Abstract
:1. Introduction
2. Breeding of Ornamental Crops
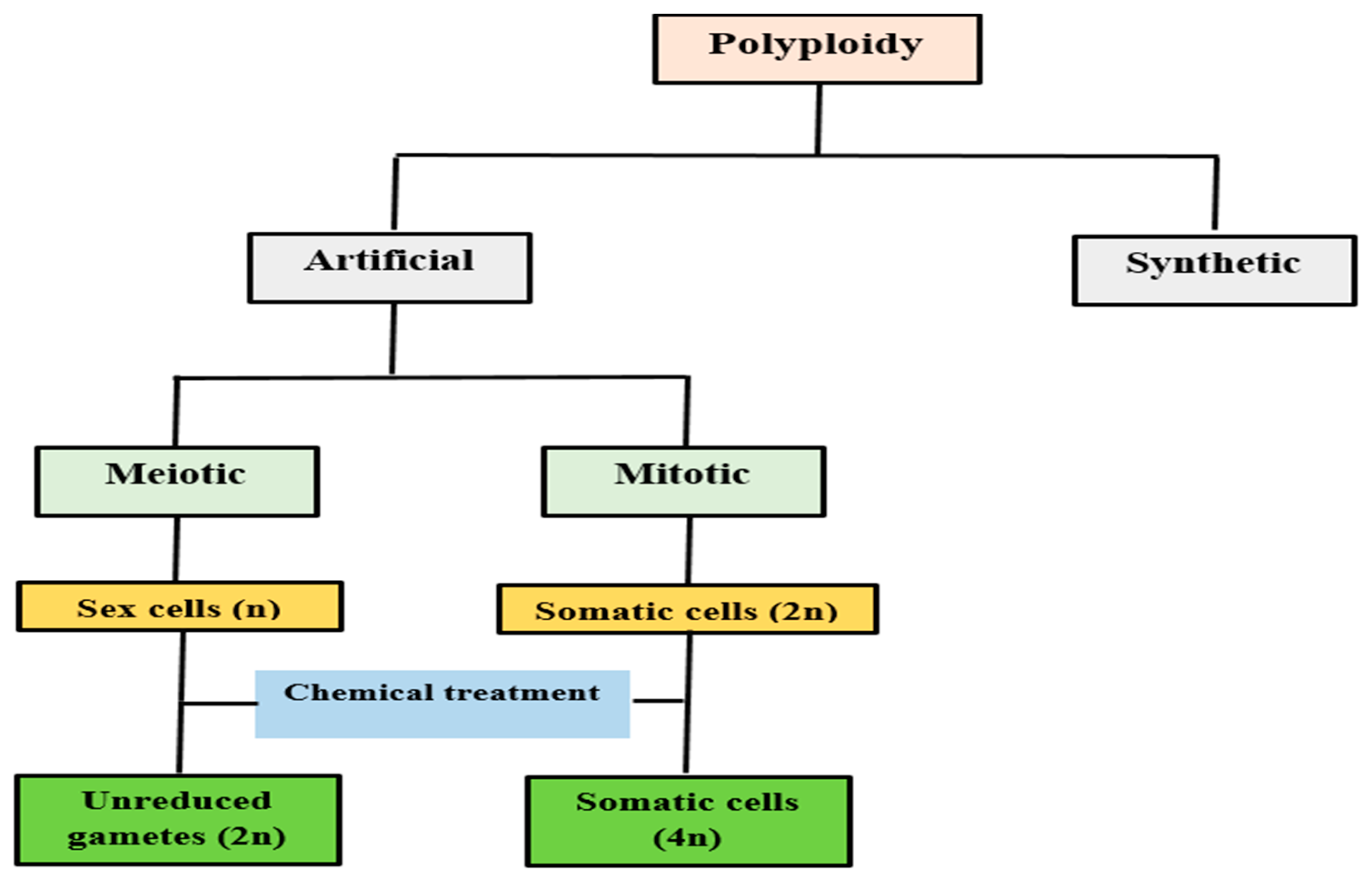
3. Induction of Polyploidy
3.1. Colchicine Induced Polyploidy/Mutation
3.2. Methods of Application
3.3. Confirmation
3.4. Chimeras
3.5. Improvement of Ornamentals by Polyploidization
3.6. Morphology
3.7. Physiology
3.8. Resorting Fertility in Wide Hybrids
3.9. Overcoming Hybridization Barrier
3.10. Pest Resistance and Stress Tolerance
4. Disadvantages
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alam, H.; Razaq, M.S. Induced polyploidy as a tool for increasing tea (Camellia sinensis L.) production. Northeast Agric. Univ. 2015, 22, 43–47. [Google Scholar] [CrossRef]
- Tavan, M.; Mirjalili, M.H.; Karimzadeh, G. In vitro polyploidy induction: Changes in morphological, anatomical and phytochemical characteristics of Thymus persicus (Lamiaceae). Plant Cell Tissue Organ Cult. 2015, 122, 573–583. [Google Scholar] [CrossRef]
- Canovas, F.M.; Luttage, U.; Matyssek, R.; Pretzsh, H. Progress in Botany; Springer: Berlin/Heidelbery, Germany, 2019; pp. 168–169. [Google Scholar]
- Zhang, C.; Cao, D.; Kang, L.; Duan, J.; Ma, X.; Yan, G.; Wang, Y. Ploidy variation and karyotype analysis in Hemerocallis spp.(Xanthorrhoeaceae) and implications on daylily breeding. N. Z. J. Crop. Hortic. Sci. 2014, 42, 183–193. [Google Scholar] [CrossRef]
- Kannan, M.; Vanitha, J.; Jiang, S.; Ramachandran, S. Impact of colchicine treatment on Sorghum bicolor BT × 623. Mol. Plant Breed. 2013, 4, 128–135. [Google Scholar] [CrossRef]
- Pereira, R.C.; Ferreira, M.T.M.; Davide, L.C.; Pasqual, M.; Mittelmann, A.; Techio, V.H. Chromosome duplication in Lolium multiflorum Lam. Crop Breed. Appl. Biotechnol. 2014, 14, 251–255. [Google Scholar] [CrossRef]
- Rêgo, M.D.; Rêgo, E.R.; Bruckner, C.H.; Finger, F.L.; Otoni, W.C. In vitro induction of autotetraploids from diploid yellow passion fruit mediated by colchicine and oryzalin. Plant Cell Tissue Organ Cult. 2011, 107, 451–459. [Google Scholar] [CrossRef]
- Mishra, B.K.; Pathak, S.; Sharma, A.; Trivedi, P.K.; Shukla, S. Modulated gene expression in newly synthesized auto-tetraploid of Papaver somniferum L. S. Afr. J. Bot. 2010, 76, 447–452. [Google Scholar] [CrossRef]
- Dewitte, A.; Van Laere, K.; Van Huylenbroeck, J. Use of 2n gametes in plant breeding. In Plant Breeding; Abdurakhmonov, I., Ed.; InTech Open Access Publisher: Rijeka, Croatia, 2011; pp. 59–86. [Google Scholar]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Comai, L. The advantages and disadvantages of being polyploid. Nature 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Ari, E.; Djapo, H.; Mutlu, N.; Gurbuz, E.; Karaguzel, O. Creation of variation through gamma irradiation and polyploidization in Vitex agnus-castus L. Sci. Hortic. 2015, 195, 74–81. [Google Scholar] [CrossRef]
- Sajjad, Y.; Jaskani, M.J.; Mehmood, A.; Ahmad, I.; Abbas, H. Effect of colchicine on in vitro polyploidy induction in African marigold (Tagetes erecta). Pak. J. Bot. 2013, 45, 1255–1258. [Google Scholar]
- Niu, L.; Tao, Y.B.; Chen, M.S.; Fu, Q.; Dong, Y.; He, H.; Xu, Z.F. Identification and characterization of tetraploid and octoploid Jatropha curcas induced by colchicine. Caryologia 2016, 69, 58–66. [Google Scholar] [CrossRef]
- Eng, W.H.; Ho, W.S. Polyploidization using colchicine in horticultural plants: A review. Sci. Hortic. 2019, 246, 604–617. [Google Scholar] [CrossRef]
- Dibyendu, T. Cytogenetic characterization of induced autotetraploids in grass pea (Lathyrus sativus L.). Caryologia 2010, 63, 62–72. [Google Scholar] [CrossRef]
- Nagahatenna, D.S.K.; Peiris, S.E. Modification of plant architecture of Hemidesmus indicus (L.) R. Br. (Iramusu) by in vitro colchicine treatment. Trop. Agric. Res. 2008, 20, 234–242. [Google Scholar]
- Obute, G.C.; Ndukwu, B.C.; Chukwu, O.F. Targeted mutagenesis in Vigna unguiculata (L.) Walp. and Cucumeropsis mannii (NAUD) in Nigeria. Afr. J. Biotechnol. 2007, 6, 2467–2472. [Google Scholar] [CrossRef]
- Dhooghe, E.; Van Laere, K.; Eeckhaut, Y.; Leus, L.; Van Huylenbroeck, J. Mitotic chromosome doubling of plant tissues in vitro. Plant Cell Tissue Organ Cult. 2011, 104, 359–373. [Google Scholar] [CrossRef]
- Laere, K.V.; França, S.C.; Vansteenkiste, H.; Huylenbroeck, J.V.; Steppe, K.; Labeke, M.C.V. Influence of ploidy level on morphology, growth and drought susceptibility in Spathiphyllum wallisii. Acta Physiol. Plant. 2011, 33, 1149–1156. [Google Scholar] [CrossRef]
- Fatima, B.; Usman, M.; Khan, M.S.; Khan, I.A.; Khan, M.M. Identification of citrus polyploids using chromosome counts, morphological and SSR markers. Pak. J. Agric. Sci. 2015, 52, 107–114. [Google Scholar]
- Podwyszyńska, M.; Gabryszewska, E.; Dyki, B.; Stępowska, A.A.; Kowalski, A.; Jasiński, A. Phenotypic and genome size changes (variation) in synthetic tetraploids of daylily (Hemerocallis) in relation to their diploid counterparts. Euphytica 2015, 203, 1–16. [Google Scholar] [CrossRef]
- Ghani, A.; Neamati, S.H.; Azizi, M.; Saharkhiz, M.J.; Farsi, M. Artificial autotetraploidy induction possibility of two Iranian endemic mint (Mentha mozaffarianii) ecotypes. Not. Sci. Biol. 2014, 6, 185–191. [Google Scholar] [CrossRef]
- Dabkevičienė, G.; Kemešytė, V.; Lemežienė, N.; Butkutė, B. Production of slender cocksfoot (Dactylis polygama H.) tetraploid populations and their assessment for agromorphological characteristics. Žemdirb. (Agric.) 2013, 100, 303–310. [Google Scholar] [CrossRef]
- Azmi, T.K.K.; Sukma, D.; Aziz, S.A.; Syukur, M. Polyploidy induction of moth orchid (Phalaenopsis amabilis (L.) Blume) by colchicine treatment on pollinated flowers. J. Agric. Sci. 2016, 11, 62–73. [Google Scholar] [CrossRef]
- Marzougui, N.; Boubaya, A.; Thabti, I.; Elfalleh, W.; Guasmi, F.; Ferchichi, A. Polyploidy induction of Tunisian Trigonella foenumgreaum L. populations. Afr. J. Biotechnol. 2011, 10, 8570–8577. [Google Scholar]
- Kumar, M.K.; Rani, M.U. Colchiploidy in fruit breeding. A review. Hortic 2013, 2, 325–326. [Google Scholar]
- El-Nashar, Y.I.; Ammar, M.H. Mutagenic influences of colchicine on phenological and molecular diversity of Calendula officinalis L. Genet. Mol. Biol. 2015, 15, 1–15. [Google Scholar] [CrossRef]
- Ade, R.; Rai, M.K. Review: Colchicine, current advances and future prospects. Nusantara Biosci. 2010, 2, 90–96. [Google Scholar]
- Castro, M.; Castro, S.; Loureiro, J. Production of synthetic tetraploids as a tool for polyploid research. Web Ecol. 2018, 18, 129–141. [Google Scholar] [CrossRef]
- Pirkoohi, M.H.; Keyvanloo, M.; Hassanpur, M. Colchicine induced polyploidy in mint by seed treatment. Int. J. Agric. Crop Sci. 2011, 3, 102–104. [Google Scholar]
- Ranney, T.G. Polyploidy: From evolution to new plant development. Proc. Int. Plant Propagators Soc. 2006, 56, 137–142. [Google Scholar]
- Lehrer, J.M.; Brand, M.H.; Lubell, J.D. Induction of tetraploidy in meristematically active seeds of Japanese barberry (Berberis thunbergii var. atropurpurea) through exposure to colchicine and oryzalin. Sci. Hortic. 2008, 119, 67–71. [Google Scholar] [CrossRef]
- Samala, S.; Te-chatoi, S. Ploidy induction through secondary somatic embryo (SSE) of oil palm by colchicine treatment. J. Agric. Technol. 2012, 8, 337–352. [Google Scholar]
- Moghbel, N.; Borujeni, M.K.; Bernard, F. Colchicine effect on the DNA content and stomata size of Glycyrrhiza glabra var. Glandulifera and Carthamus tinctorius L. cultured in vitro. J. Genet. Eng. Biotechnol. 2015, 13, 1–6. [Google Scholar] [CrossRef]
- Sakhanokho, H.F.; Rajasekaran, K.; Kelley, R.Y.; Islam-Faridi, N. Induced polyploidy in diploid ornamental ginger (Hedychium muluense RM Smith) using colchicine and oryzalin. HortScience 2009, 44, 1809–1814. [Google Scholar] [CrossRef]
- Xu, L.; Najeeb, U.; Naeem, M.S.; Daud, M.K.; Cao, J.S.; Gong, H.J.; Sheen, W.Q.; Zhou, W.J. Induction of tetraploidy in Juncus effusus by colchicine. Biol. Plant. 2010, 54, 659–663. [Google Scholar] [CrossRef]
- Harbard, J.L.; Griffin, A.R.; Foster, S.; Brooker, C.; Kha, L.D.; Koutoulis, A. Production of colchicine-induced autotetraploids as a basis for sterility breeding in Acacia mangium Willd. Forestry 2012, 85, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Khaing, T.T.; Perera, A.L.T.; Sumanasinghe, V.A.; Wijesundara, D.S.A. Improvement of Gymnostachyum species by induced mutation. Trop. Agric. Res. 2007, 19, 265–272. [Google Scholar]
- Vichiato, M.R.M.; Vichiato, M.; Pasqual, M.; Castro, D.M.D.; Dutra, L.F. Tetraploidy induction and identification in Dendrobium nobile Lindl (Orchidaceae). Rev. Cienc. Agron. 2007, 38, 385–390. [Google Scholar]
- Balode, A. Applying colchicine and oryzaline in Lilium L polyploidization. L. J. Agron. 2008, 11, 22–28. [Google Scholar]
- Kobayashi, N.; Yamashita, S.; Ohta, K.; Hosoki, T. Morphological characteristics and their inheritance in colchicine-induced salvia polyploids. J. Jpn. Soc. Hortic. Sci. 2008, 77, 186–191. [Google Scholar] [CrossRef]
- El-Morsy, S.I.; Dorra, M.D.M.; Abd El-Hady, E.A.A.; Hiaba, A.A.A.; Mohamed, A.Y. Comparative studies on diploid and tetraploid levels of Nicotiana alata. Acad. J. Plant Sci. 2009, 2, 182–188. [Google Scholar]
- Majdi, M.; Karimzadeh, G.; Malboobi, M.A.; Omidbaigi, R.; Mirzaghaderi, G. Induction of tetraploidy to feverfew (Tanacetum parthenium Schulz-Bip.): Morphological, physiological, cytological, and phytochemical changes. HortScience 2010, 45, 16–21. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Mishra, S.K. Effect of colchicine on mitotic polyploidization and morphological characteristics of Phlox drummondi. Afr. J. Biotechnol. 2012, 11, 9336–9342. [Google Scholar] [CrossRef]
- Boonbongkarn, S.; Taychasinpitak, T.; Wongchaochant, S.; Kikuchi, S. Effect of colchicine tablets on morphology of Torenia fournieri. Int. Trans. J. Eng. Manag. Sci. Technol. 2013, 4, 299–309. [Google Scholar]
- Grouh, M.S.H.; Meftahizade, H.; Lotfi, N.; Rahimi, V.; Baniasadi, B. Doubling the chromosome number of Salvia hains using colchicine: Evaluation of morphological traits of recovered plants. J. Med. Plants Res. 2011, 5, 4892–4898. [Google Scholar]
- Kwon, S.J.; Roy, S.K.; Cho, K.Y.; Moon, Y.J.; Woo, S.H.; Kim, H.H. Tetraploid induction approach induced by colchicine of Prunella vulgaris L. F. albiflora Nakai. Int. J. Sci. Res. Pub. 2014, 4, 1–7. [Google Scholar]
- Urwin, N.A. Generation and characterization of colchicine-induced polyploid Lavandula × intermedia. Euphytica 2014, 197, 331–339. [Google Scholar] [CrossRef]
- Vichiato, M.R.D.M.; Vichiato, M.; Pasqual, M.; Rodrigues, F.A.; Castro, D.M.D. Morphological effects of induced polyploidy in Dendrobium nobile Lindl. (Orchidaceae). Crop Breed. Appl. Biot. 2014, 14, 154–159. [Google Scholar] [CrossRef]
- He, Y.; Sun, Y.; Zheng, R.; Ai, Y.; Cao, Z.; Bao, M. Induction of tetraploid male sterile Tagetes erecta by colchicine treatment and its application for interspecific hybridization. Hortic. Plant J. 2016, 2, 284–292. [Google Scholar] [CrossRef]
- Feng, H.; Wang, M.L.; Cong, R.C.; Dai, S.L. Colchicine-and trifluralin-mediated polyploidization of Rosa multiflora Thunb. var. inermis and Rosa roxburghii F. normalis. J. Hortic. Sci. Biotechnol. 2017, 92, 279–287. [Google Scholar] [CrossRef]
- Mori, S.; Yamane, T.; Yahata, M.; Shinoda, K.; Murata, N. Chromosome doubling in Limonium bellidifolium (Gouan) Dumort. by colchicine treatment of seeds. Hortic. J. Preview 2016, 85, 366–371. [Google Scholar] [CrossRef]
- Li, Z.; Ruter, J.M. Development and Evaluation of diploid and polyploid Hibiscus moscheutos. HortScience 2017, 52, 676–681. [Google Scholar] [CrossRef]
- Basumatari, M.; Das, B.N. Karyomorphological studies in two species of Bauhinia Linn. and induction of polyploidy in Bauhinia acuminata Linn. Int. J. Life Sci. Sci. Res. 2017, 3, 1223–1229. [Google Scholar] [CrossRef]
- Wang, W.; He, Y.; Cao, Z.; Deng, Z. Induction of tetraploids in impatiens (Impatiens walleriana) and characterization of their changes in morphology and resistance to downy mildew. HortScience 2018, 53, 925–931. [Google Scholar] [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Baig, M.M.Q.; Quresh, A.A.; Shah, M.K.N.; Hafiz, I.A. Induction and identification of colchicine induced polyploidy in Gladiolus grandiflorus ‘White Prosperity’. Folia Hortic. 2018, 30, 307–319. [Google Scholar] [CrossRef]
- Suzuki, K.; Takatsu, Y.; Gonai, T.; Kasumi, M. Plant regeneration and chromosome doubling of wild Gladiolus species. Acta Hortic. 2005, 673, 175–181. [Google Scholar] [CrossRef]
- Anurita, D.; Girjesh, K. Morphogenetic analysis of colchitetraploids in Impatiens balsamina L. Caryologia 2007, 60, 199–202. [Google Scholar]
- Seneviratne, K.A.C.N.; Wijesundara, D.S.A. First African violets (Saintpaulia ionantha, H. Wendl.) with a changing colour pattern induced by mutation. Am. J. Plant Physiol. 2007, 2, 233–236. [Google Scholar]
- Ning, G.G.; Shi, X.P.; Hu, H.R.; Yan, Y.; Bao, M.Z. Development of a range of polyploid lines in Petunia hybrida and the relationship of ploidy with the single-double-flower trait. HortScience 2009, 44, 250–255. [Google Scholar] [CrossRef]
- Ye, Y.M.; Tong, J.; Shi, X.P.; Yuan, W.; Li, G.R. Morphological and cytological studies of diploid and colchicine-induced tetraploid lines of crape myrtle (Lagerstroemia indica L.). Sci. Hortic. 2010, 124, 95–101. [Google Scholar] [CrossRef]
- Sadhukhan, R.; Ganguly, A.; Singh, P.K.; Sarkar, H.K. Study of Induced polyploidy in African marigold (Tagetes ecrecta L.). Environ. Ecol. 2014, 32, 1219–1222. [Google Scholar]
- He, M.; Gao, W.; Gao, Y.; Liu, Y.; Yang, X.; Jiao, H.; Zhou, Y. Polyploidy induced by colchicine in Dendranthema indicum var. Aromaticum, a scented chrysanthemum. Eur. J. Hortic. Sci. 2016, 81, 219–226. [Google Scholar] [CrossRef]
- Anitha, K.; Jawaharlal, M.; Joel, J.; Surendranath, R. Induction of polyploids and isolation of ploidy variants through stomatal parameters in bougainvillea. Int. J. Agric. 2017, 7, 451–458. [Google Scholar]
- Kushwah, K.S.; Verma, R.C.; Patel, S.; Jain, N.K. Colchicine induced polyploidy in Chrysanthemum carinatum L. J. Phylogenet. Evol. Biol. 2018, 6, 2–4. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, F.; Zhao, X.; Yang, W. Identification of tetraploid mutants of Platycodon grandiflorus by colchicine induction. Caryologia 2011, 64, 343–349. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, F.; Zhao, X.; Yang, W.D. Cytogenetic characterization of induced tetraploids in medicinal plant (Platycodon grandiflorus). Caryologia 2012, 65, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, W.; Dong, J.; Yang, N.; Zhao, X.; Yang, W. Tetraploid induction and cytogenetic characterization for Clematis heracleifolia. Caryologia 2013, 66, 215–220. [Google Scholar] [CrossRef]
- Fetouh, M.I.; Kareem, A.; Knox, G.W.; Wilson, S.B.; Deng, Z. Induction, identification, and characterization of tetraploids in Japanese privet (Ligustrum japonicum). HortScience 2016, 51, 1371–1377. [Google Scholar] [CrossRef]
- Liu, G.; Li, Z.; Bao, M. Colchicine-induced chromosome doubling in Platanus acerifolia and its effect on plant morphology. Euphytica 2007, 157, 145–154. [Google Scholar] [CrossRef]
- Jadrná, P.; Plavcová, O.; Kobza, F. Morphological changes in colchicine treated Pelargonium×Hortorum LH bailey greenhouse plants. HortScience 2010, 37, 27–33. [Google Scholar]
- Du, X.; Sun, Y.; Yuan, S.; Li, Q.; Gong, Z. Identification of colchicines induced polyploid plants in two species of Impatiens balsamina. Acta Agric. Bor. Sin. 2011, 20, 56–59. [Google Scholar]
- Hosseini, H.; Chehrazi, M.; Sorestani, M.M.; Ahmadi, D. Polyploidy and comparison of diploid and autotetraploid seedling of Madagascar periwinkle (Catharanthus roseus cv. Alba). Int. Res. J. Appl. Basic Sci. 2013, 4, 402–406. [Google Scholar]
- Gu, Y. Cut Flower Productivity and Economic Analysis, Polyploidy Induction in Two Zinnia Varieties, Zinnia Pollination Mechanisms and DNA Content of Zinnia Species. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2015. [Google Scholar]
- Talebi, S.F.; Saharkhiz, M.J.; Kermani, M.J.; Sharafi, Y.; Raouf Fard, F. Effect of different antimitotic agents on polyploid induction of anise hyssop (Agastache foeniculum L.). Caryologia 2017, 70, 184–193. [Google Scholar] [CrossRef]
- Soetopo, L.; Hosnia, D. In vivo polyploid-induction by colchicine on orchids Phalaenopsis pulcherrima (Lindl.) JJ Smith. Biosci. Res. 2018, 15, 941–949. [Google Scholar]
- Lertsutthichawan, A.; Ruamrungsri, S.; Duangkongsan, W.; Saetiew, K. Induced mutation of chrysanthemum by colchicine. Int. J. Agric. Technol. 2017, 13, 2325–2332. [Google Scholar]
- Botelho, F.B.S.; Rodrigues, C.S.; Bruzi, A.T. Ornamental plant breeding. Ornam Hortic. 2015, 21, 9–16. [Google Scholar] [CrossRef]
- Prabhukumar, K.M.; Thomas, V.P.; Sabu, M.; Prasanth, M.V.; Mohanan, K.V. Induced mutation in ornamental gingers (Zingiberaceae) using chemical mutagens viz. colchicine, acridine and ethyl methane sulphonate. J. Hortic. For. Biotechnol. 2015, 19, 18–27. [Google Scholar]
- Rathod, A.D.; Patil, S.R.; Taksande, P.N.; Karad, G.W.; Kalamkar, V.B.; Jayade, V.S. Effect of colchicine on morphological and biometrical traits in African marigold. J. Soils Crops 2018, 28, 72–80. [Google Scholar]
- Mostafa, G.G.; Alhamd, M.F.A. Detection and evaluation the tetraploid plants of Celosia argentea induced by colchicine. Int. J. Plant Breed. Genet. 2016, 10, 110–115. [Google Scholar] [CrossRef]
- Brits, G.J.; Ling, L. Polyploid breeding of wild South African Plectranthus (spurflowers) as new flowering pot plants. Acta Hortic. 2008, 774, 437–442. [Google Scholar] [CrossRef]
- Bharadwaj, D.N. Plant Biology and Biotechnology: Polyploidy in Crop Improvement and Evolution; Springer: New Dehli, India, 2015; pp. 619–638. [Google Scholar]
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity 2013, 110, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Wakachaure, R.; Ganguly, S. Chromosomal polyploidy: A brief review. Pha. Biol. Eval. 2016, 3, 510–512. [Google Scholar]
- Manzoor, A. Induction and Identification of Polyploidy in Gladiolus Grandiflorus through Chemical Treatment. Master’s Thesis, Arid Agriculture University, Rawalpindi, Pakistan, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Method of Application | Most Successful Treatment | Reference |
|---|---|---|---|
| Gymnostachyum (Gymnostachyum zeylanicum) | Dipping of shoot cuttings in colchicine solution | Colchicine 0.1%, 24 h | Khaing et al. (2007) [39] |
| Orchid (Dendrobium nobile) | Immersing whole plant in colchicine solution | Colchicine 0.1%, 96 h | Vichiato et al. (2007) [40] |
| Lily (Lilium) | Treating bulb scales with colchicine solution | Colchicine 1%, 24 h | Balode (2008) [41] |
| Salvia (Salvia coccinea cv. Coral Nymph and cv. Lady in Red and Salvia patens cv. Blue Angel) | Seeds soaked in colchicine solution | Colchicine 0.05%, 24 h (S. coccinea cv. Coral Nymph) | Kobayashi et al. (2008) [42] |
| Japanese barberry (Berberis thunbergii) | Pre-germinated seeds immersed in colchicine solution | Colchicine 0.2%, 24 h | Lehrer et al. (2008) [33] |
| Jasmine tobacco (Nicotiana alata) | Dipping seeds in colchicine solution | Colchicine 0.5%, 24 h | El-Morsy et al. (2009) [43] |
| Feverfew (Tanacetum parthenium) | Dipping roots in colchicine solution | Colchicine 0.05% 6 h | Majdi et al. (2010) [44] |
| Phlox (Phlox drummondi) | Seed soaking method | Colchicine 0.5%, 36 h | Tiwari and Mishra (2012) [45] |
| Wishbone flower (Torenia fournieri) | Dipping of leaf petiole in colchicine solution | Colchicine 0.015% 3 d | Boonbongkarn et al. (2013) [46] |
| Salvia (Salvia hains) | Seeds dipped in colchicine aqueous solution | Colchicine 0.3 to 0.5%, 24 h | Grouh et al. (2013) [47] |
| Self-heal (Prunella vulgaris L.F. abiflora nakai) | Soaking of seeds in colchicine solution | Colchicine 0.05% and 0.1%, 6 h | Kwon et al. (2014) [48] |
| Lavandins (Lavandula × intermedia cvs. Grosso and Seal) | First method: Treatment of upper nodes of whole plant with colchicine Second method: Fresh cuttings completely submerged in colchicine | Colchicine 0.1%, 6 h (cutting method) | Urwin (2014) [49] |
| Orchid (Dendrobium nobile) | Plant of height 5cm was dipped in colchicine solution | Colchicine 0.1%, 96 h | Vichiato et al. (2014) [50] |
| Chaste tree (Vitex agnus castus L) | Seed soaking method | Colchicine 0.05%, 36 h | Ari et al. (2015) [12] |
| Calendula (Calendula officinalis) | Soaking of seeds in colchicine solution | Colchicine 0.08%, 4 h | El-Nashar and Ammar, (2015) [28] |
| Marigold (Tagetes erecta) | Pre-germinated seeds treated with colchicine | Colchicine 0.1%, 3-6 h: 0.2%, 3 h | He et al. (2016) [51] |
| Rose (Rosa multiflora var inermis, Rosa roxburgii var normalis) | First method: Germinated seeds soaked in colchicine solution Second method: Seedlings at cotyledon stage treated with colchicine | Germinating seeds method Colchicine 0.2%, 12 h | Feng et al. (2016) [52] |
| Matted sea-lavender (Limonium bellidifolium) | Seed soaking treatment | Colchicine 0.05%, 72 h | Mori et al. (2016) [53] |
| Swamp rosemallow (Hibiscus moscheutos) | Seedlings at cotyledon stage soaked in colchicine solution | Colchicine 0.1%, 24 h | Li and Ruter, (2017) [54] |
| White orchid tree (Bauhinia acuminata) | Treatment of seeds with colchicine solution | Colchicine 0.1%, 12 h (3 consecutive days | Basumatari and Das, (2017) [55] |
| Sultana (Impatiens walleriana) | Seed soaking method | Colchicine 0.05%, 48 h (2 days) | Wang and He, (2018) [56] |
| Gladiolus (Gladiolus grandiflorus) | Corm soaked in colchicine solution | Colchicine 0.1-0.3%, 24 h | Manzoor et al. (2018) [57] |
| Crop | Method of Application | Most Successful Treatment | Reference |
|---|---|---|---|
| Gladiolus wild species (Gladiolus tristis) | Colchicine applied to apical buds of corms through cotton wool | -------- | Suzuki et al. (2005) [58] |
| Garden balsam (Impatiens balsamina) | Application of colchicine to seedlings through cotton plug | Colchicine 0.4%, 3 d | Anurita and Girjesh (2007) [59] |
| African violets (Saintpaulia ionantha) | Leaf petiole treated by cotton-based method | All colchicine concentration (0.04%, 0.06% and 0.09%), 22.5-23.5 h | Seneviratine and Wijesundra (2007) [60] |
| Petunia (Petunia hybrida) | Shoot apics of seedlings were treated with colchicine through cotton wool | Colchicine 0.2%, 48 h | Ning et al. (2009) [61] |
| Crape myrtle (Lagerstroemia indica cvs. Zi Wei, Hong Wei and Yin Wei) | Colchicine applied to apical meristerm of young seedlings through cotton wool | Colchicine 0.5%, 72 h (cv. Zi Wei); 0.8%, 48 h (cv. Hong Wei.); 0.2%, 96 h; 0.5%, 48 h; 0.8%, 72 h (cv. Yin Wei.) | Ye et al. (2010) [62] |
| African marigold (Tagates erecta) | First method: Placing of whole plant with roots in a colchicine solution Second method: Treatment of apical buds with colchicine-soaked cotton | Colchicine 0.0005%, 6 h | Sadhukhan et al. (2014) [63] |
| Chrysanthemum (Dendranthema indicum var. aromaticum) | First method: Grin seeds soaked in colchicine solution Second method: Pre-germinated seeds dipped in colchicine aqueous solution Third method: Cotton balls placed on shoot tips; colchicine poured on cotton balls through micropipette | Colchicine 0.1%, 24 h (grin seeds); 0.1%, 7 d (shoot tips) | He et al. (2016) [64] |
| Bougainvillea (Bougainvillea spp cvs Lalbagh, Mahara) | Colchicine soaked cotton placed on dormant nodal buds | Colchicine 0.4% for 72 h (cv. Lalbagh); 0.3% for 72 h (cv. Mahara) | Anitha et al. (2017) [65] |
| Chrysanthemum (Chrysanthemum carinatum) | Apical buds treated with colchicine-soaked cotton swab | Colchicine 0.2 %, 3 days with 6 h duration per day | Kushwah et al. (2018) [66] |
| Crop | Method of Application | Most Successful Treatment | Reference |
|---|---|---|---|
| Balloon flower (Platycodon grandiflorus) | Warm semi solid (1% agar) colchicine applied to the apical buds of the seedlings | Colchicine 0.5%, 72 h | Wu et al. (2011) [67] |
| Balloon flower (Platycodon grandiflorus) | Treatment of apical buds with colchicine | Colchicine 0.5%, 72 h | Wu et al. (2012) [68] |
| Clematis (Clematis heracleifolia) | Apical buds treated with warm semi solid colchicine (1% agar) | Colchicine 0.2%, 48 h | Wu et al. (2013) [69] |
| Japanese privet (Ligustrum japonicum) | Newly growing points treated with colchicine stock (0.55% semi solid agar) | Colchicine 0.2%, three consecutive days | Fetouh et al. (2016) [70] |
| Crop | Method of Application | Most Successful Treatment | Reference |
|---|---|---|---|
| London plane (Planatus acerifolius) | First method: Seeds soaked in colchicine solution Second method: Apical meristems of seedling treated with colchicine solution through dropping method | Colchicine 0.3%-0.4%, 24 h | Liu et al. (2007) [71] |
| Pelargonium (Pelargonium graveolens cvs. Black Velvet Scarlet F1 and Gizela F1) | Colchicine applied on seedling apics at true leaf stage | Colchicine 0.5%, 3 d (cv Black Velvet Scarlet F1); 1.0%, 2 d; 2.5%, 2 d; 0.5%, 5 d (cv. Gizela F1) | Jadrna et al. (2010) [72] |
| Garden balsam (Impatiens balsamina sp Xinhua impatiens and Camellia impatiens) | Application of colchicine to shoot apics of seedlings | Colchicine 0.5%, 96 h (Xinhua impatiens); 0.5%, 72 h (Camellia impatiens) | Xiaohua et al. (2011) [73] |
| Madagascar periwinkle (Catharanthus roseus) | Seedling shoot apics treatment with colchicine | Colchicine 0.4%, 7 d | Hosseini et al. (2013) [74] |
| Zinnia (Zinnia violacea cv. Oklahoma White and Zinnia angustifolia cv. Crystal Orange) | Colchicine applied drop wise to growing points | Colchicine 0.33% | Gu (2015) [75] |
| Anise hyssop (Agastache foeniculum) | Apical meristems treated drop wise with colchicine | Colchicine 0.006% for three successive days | Talebi et al. (2017) [76] |
| Moth orchid (Phalaenopsis pulcherrima) | Treatment of seeds with colchicine through dropping method | Colchicine, 0.5% | Soetopo and Hosnia. (2018) [77] |
| Chrysanthemum (Dendranthema grandiflorus) | Cotton covered shoots treated drop wise with colchicine | Colchicine, 0.8% for 6 days | Lertsutthichawan et al. (2018) [78] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants. Plants 2019, 8, 194. https://doi.org/10.3390/plants8070194
Manzoor A, Ahmad T, Bashir MA, Hafiz IA, Silvestri C. Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants. Plants. 2019; 8(7):194. https://doi.org/10.3390/plants8070194
Chicago/Turabian StyleManzoor, Ayesha, Touqeer Ahmad, Muhammad Ajmal Bashir, Ishfaq Ahmad Hafiz, and Cristian Silvestri. 2019. "Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants" Plants 8, no. 7: 194. https://doi.org/10.3390/plants8070194