Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants

 , ,
, ,
Abstract
:1. Introduction
2. Heavy Metal: From Essentiality to Toxicity
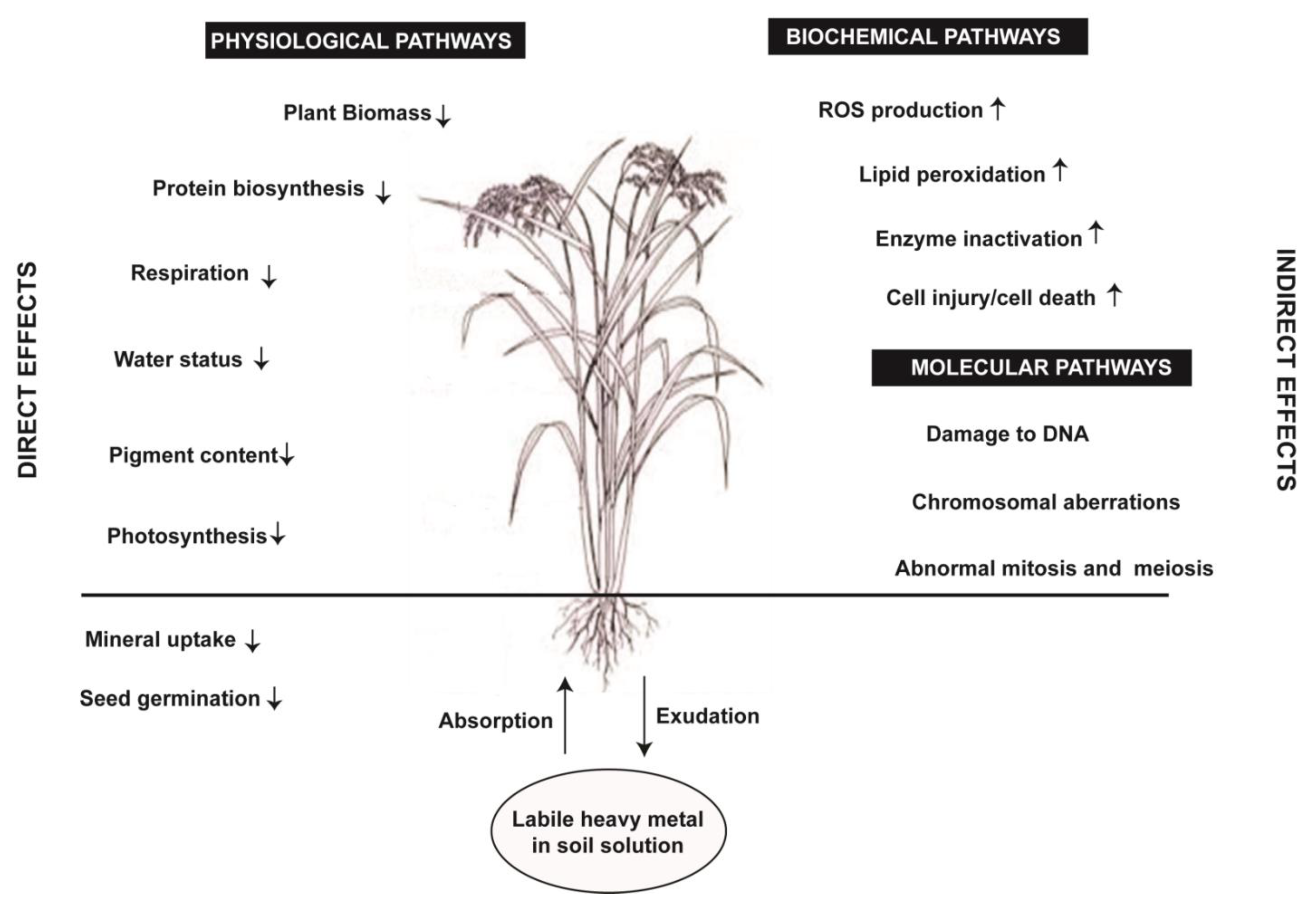
3. Heavy Metal Toxicity and Crop Plants
4. Silicon: A Multifaceted Element for Alleviating Heavy Metal Toxicity in Crops
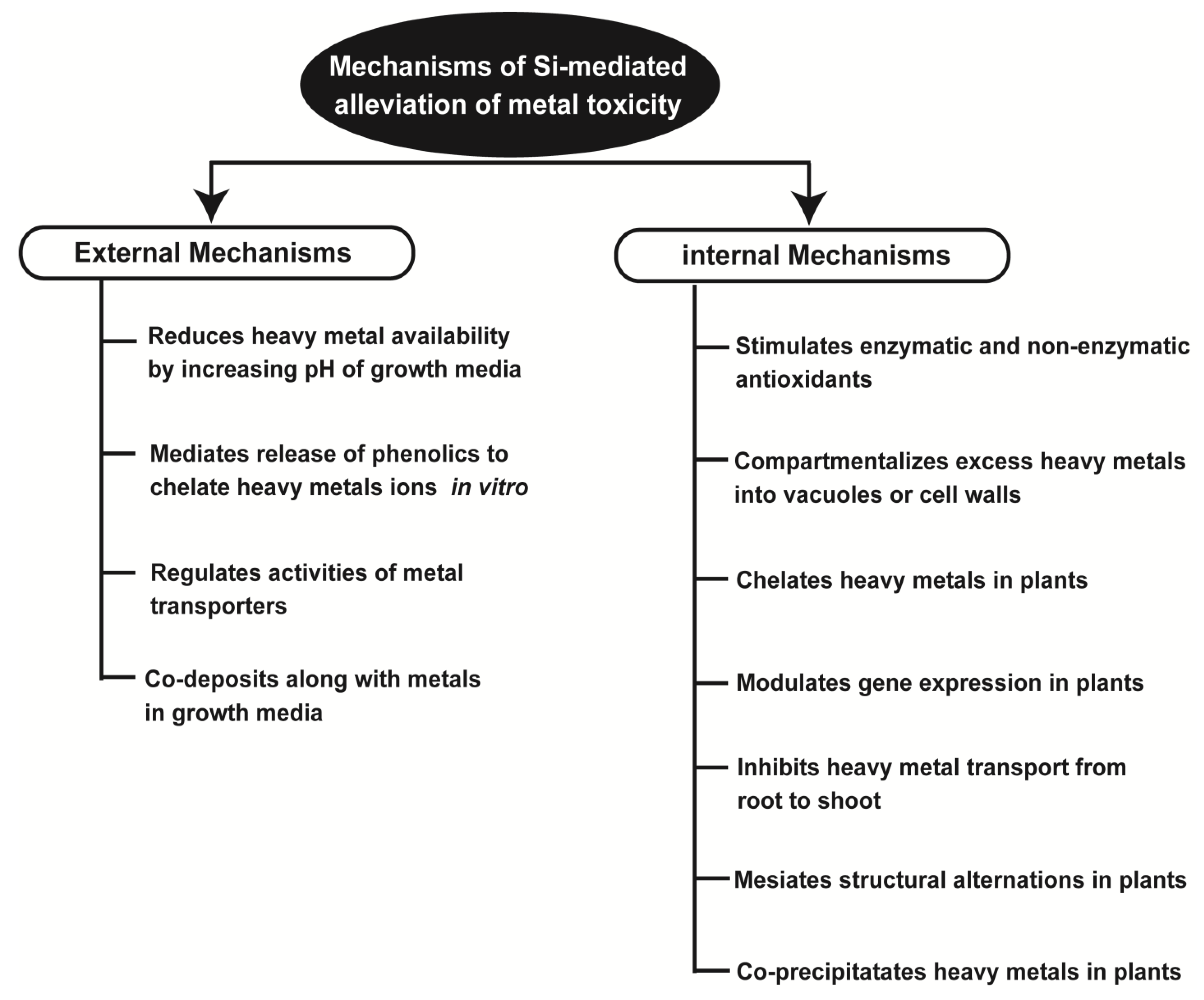
5. Silicon-Mediated Mechanisms of Metal Detoxification
5.1. Silicon Mediated Immobilization of Toxic Metal in the Soil
5.2. Stimulation of Antioxidant Defense System
5.3. Compartmentation within Plants
5.4. Chelation Mediated Metal Toxicity Reduction with Silicon Application
5.5. Regulation of Gene Expression
5.6. Structural Alterations Related to Metal Stress Tolerance in Plants
5.7. Co-Precipitation of Metals by Silicon Application in Soil
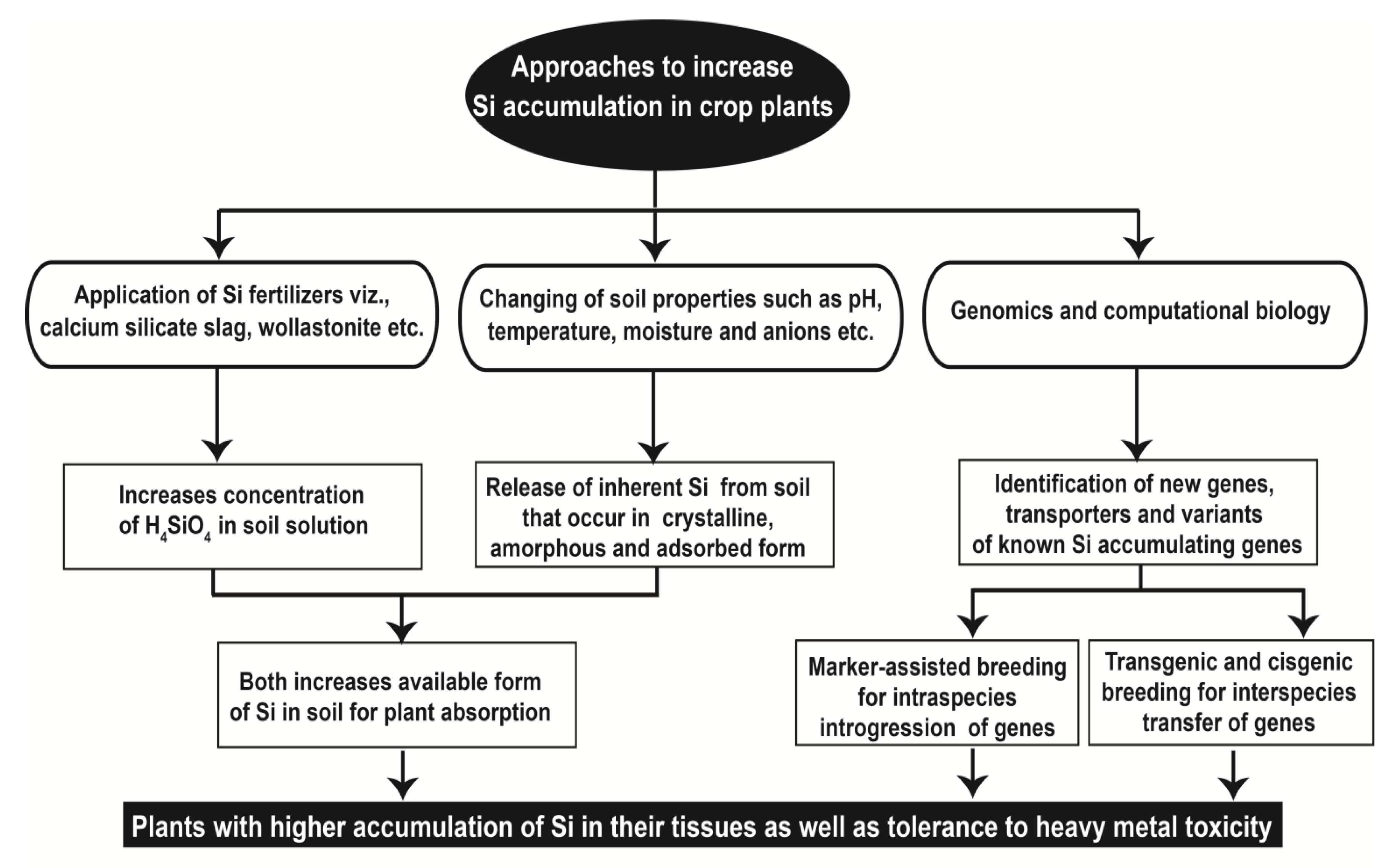
6. Approaches for Improving Silicon Accumulation in Crop Plants
6.1. Silicon Fertilization
6.2. Modification of Soil Properties
6.3. Approaches for Genetic Gain
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawkes, S.J. What Is a” Heavy Metal”? J. Chem. Educ. 1997, 74, 1374. [Google Scholar] [CrossRef]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Kim, D.-H.; Lee, S.-Y.; Kim, K.-M.; Waqas, M.; Jung, H.-Y.; Shin, J.-H.; Kim, J.-G.; Lee, I.-J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.-G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Ma, J.F.; Bélanger, R.R. Role of silicon in plants. Front. Plant Sci. 2017, 8, 1858. [Google Scholar] [CrossRef]
- Rasoolizadeh, A.; Labbé, C.; Sonah, H.; Deshmukh, R.K.; Belzile, F.; Menzies, J.G.; Bélanger, R.R. Silicon protects soybean plants against Phytophthora sojae by interfering with effector-receptor expression. BMC Plant Biol. 2018, 18, 97. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture: From Theory to Practice; Springer: Berlin, Germany, 2015. [Google Scholar]
- Chérif, M.; Asselin, A.; Bélanger, R. Defense responses induced by soluble silicon in cucumber roots infected by Pythium spp. Phytopathology 1994, 84, 236–242. [Google Scholar] [CrossRef]
- Wang, Y.; Stass, A.; Horst, W.J. Apoplastic binding of aluminum is involved in silicon-induced amelioration of aluminum toxicity in maize. Plant Physiol. 2004, 136, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Naseri Rad, S.; Ali, N.; Yvin, J.-C. The Ameliorative Effect of Silicon on Maize Plants Grown in Mg-Deficient Conditions. Int. J. Mol. Sci. 2019, 20, 969. [Google Scholar] [CrossRef] [PubMed]
- Fauteux, F.; Chain, F.; Belzile, F.; Menzies, J.G.; Bélanger, R.R. The protective role of silicon in the Arabidopsis–powdery mildew pathosystem. Proc. Natl. Acad. Sci. USA 2006, 103, 17554–17559. [Google Scholar] [CrossRef]
- Hodson, M.; White, P.; Mead, A.; Broadley, M. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Bélanger, R.R. Molecular evolution of aquaporins and silicon influx in plants. Funct. Ecol. 2016, 30, 1277–1285. [Google Scholar] [CrossRef]
- Kalaivanan, D.; Ganeshamurthy, A.N. Mechanisms of heavy metal toxicity in plants. In Abiotic Stress Physiology of Horticultural Crops; Springer: Berlin, Germany, 2016; pp. 85–102. [Google Scholar]
- Ali, S.; Bai, P.; Zeng, F.; Cai, S.; Shamsi, I.H.; Qiu, B.; Wu, F.; Zhang, G. The ecotoxicological and interactive effects of chromium and aluminum on growth, oxidative damage and antioxidant enzymes on two barley genotypes differing in Al tolerance. Environ. Exp. Bot. 2011, 70, 185–191. [Google Scholar] [CrossRef]
- Zia-ur-Rehman, M.; Sabir, M.; Nadeem, M. Remediating cadmium-contaminated soils by growing grain crops using inorganic amendments. In Soil Remediation and Plants: Prospects and Challenges; Elsevier Inc., Academic Press: Amsterdam, The Netherlands, 2015; pp. 367–396. [Google Scholar]
- Foucault, Y.; Lévèque, T.; Xiong, T.; Schreck, E.; Austruy, A.; Shahid, M.; Dumat, C. Green manure plants for remediation of soils polluted by metals and metalloids: Ecotoxicity and human bioavailability assessment. Chemosphere 2013, 93, 1430–1435. [Google Scholar] [CrossRef]
- Wu, J.; Shi, Y.; Zhu, Y.; Wang, Yi.; Gong, H. Mechanisms of enhanced heavy metal tolerance in plants by silicon: A review. Pedosphere 2013, 23, 815–825. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Siti Nor Akmar, A.; Rafii, M.Y.; Azizi, P.; Tengoua, F.; Nurul Mayzaitul Azwa, J.; Shabanimofrad, M. Importance of silicon and mechanisms of biosilica formation in plants. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Cocker, K.M.; Evans, D.E.; Hodson, M.J. The amelioration of aluminium toxicity by silicon in higher plants: Solution chemistry or an in planta mechanism? Physiol. Plant. 1998, 104, 608–614. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, L.; Nie, Q.; Zhang, W.; Zhang, F. Long-term effects of exogenous silicon on cadmium translocation and toxicity in rice (Oryza sativa L.). Environ. Exp. Bot. 2008, 62, 300–307. [Google Scholar] [CrossRef]
- Shi, Z.; Yang, S.; Han, D.; Zhou, Z.; Li, X.; Liu, Y.; Zhang, B. Silicon alleviates cadmium toxicity in wheat seedlings (Triticum aestivum L.) by reducing cadmium ion uptake and enhancing antioxidative capacity. Environ. Sci. Pollut. Res. 2018, 25, 7638–7646. [Google Scholar] [CrossRef] [PubMed]
- Greger, M.; Landberg, T.; Vaculík, M. Silicon Influences Soil Availability and Accumulation of Mineral Nutrients in Various Plant Species. Plants 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.-H.; Qiu, H.; Tian, T.; Zhan, S.-S.; Chaney, R.L.; Wang, S.-Z.; Tang, Y.-T.; Morel, J.-L.; Qiu, R.-L. Mitigation effects of silicon rich amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on multi-metal contaminated acidic soil. Chemosphere 2011, 83, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Anwaar, S.A.; Ali, S.; Ali, S.; Ishaque, W.; Farid, M.; Farooq, M.A.; Najeeb, U.; Abbas, F.; Sharif, M. Silicon (Si) alleviates cotton (Gossypium hirsutum L.) from zinc (Zn) toxicity stress by limiting Zn uptake and oxidative damage. Environ. Sci. Pollut. Res. 2015, 22, 3441–3450. [Google Scholar] [CrossRef]
- Patrícia Vieira da Cunha, K.; Williams Araújo do Nascimento, C.; José da Silva, A. Silicon alleviates the toxicity of cadmium and zinc for maize (Zea mays L.) grown on a contaminated soil. J. Plant Nutr. Soil Sci. 2008, 171, 849–853. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, C.; Wang, H.; Zhang, F. Effect of Si on the distribution of Cd in rice seedlings. Plant Soil 2005, 272, 53–60. [Google Scholar] [CrossRef]
- Hammond, K.E.; Evans, D.E.; Hodson, M.J. Aluminium/silicon interactions in barley (Hordeum vulgare L.) seedlings. Plant Soil 1995, 173, 89–95. [Google Scholar] [CrossRef]
- Singh, V.P.; Tripathi, D.K.; Kumar, D.; Chauhan, D.K. Influence of exogenous silicon addition on aluminium tolerance in rice seedlings. Biol. Trace Elem. Res. 2011, 144, 1260–1274. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Xiao, X.; Dong, Z.; Chen, Y. Silicon effects on antioxidative enzymes and lipid peroxidation in leaves and roots of peanut under aluminum stress. Acta Physiol. Plant. 2014, 36, 3063–3069. [Google Scholar] [CrossRef]
- Haynes, R.J. A contemporary overview of silicon availability in agricultural soils. J. Plant Nutr. Soil Sci. 2014, 177, 831–844. [Google Scholar] [CrossRef]
- Li, L.; Zheng, C.; Fu, Y.; Wu, D.; Yang, X.; Shen, H. Silicate-mediated alleviation of Pb toxicity in banana grown in Pb-contaminated soil. Biol. Trace Elem. Res. 2012, 145, 101–108. [Google Scholar] [CrossRef]
- Liang, Y.; Wong, J.; Wei, L. Silicon-mediated enhancement of cadmium tolerance in maize (Zea mays L.) grown in cadmium contaminated soil. Chemosphere 2005, 58, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.J.; Evans, D.E. Aluminium/silicon interactions in higher plants. J. Exp. Bot. 1995, 46, 161–171. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, S.; Li, S.; Liao, X.; Wang, R. Silicon Mediated the Detoxification of Cr on Pakchoi (Brassica Chinensis L.) in Cr-contaminated Soil. Procedia Environ. Sci. 2013, 18, 58–67. [Google Scholar] [CrossRef]
- Shim, J.; Shea, P.J.; Oh, B.-T. Stabilization of heavy metals in mining site soil with silica extracted from corn cob. WaterAir Soil Pollut. 2014, 225, 2152. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Tripathi, D.K.; Deshmukh, R.; Singh, V.P.; Corpas, F.J. Revisiting the role of ROS and RNS in plants under changing environment. Environ. Exp. Bot. 2019, in press. [Google Scholar] [CrossRef]
- Shi, Q.; Bao, Z.; Zhu, Z.; He, Y.; Qian, Q.; Yu, J. Silicon-mediated alleviation of Mn toxicity in Cucumis sativus in relation to activities of superoxide dismutase and ascorbate peroxidase. Phytochemistry 2005, 66, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, H.; Zhang, Y.; Chai, T. Silicon attenuates cadmium toxicity in Solanum nigrum L. by reducing cadmium uptake and oxidative stress. Plant Physiol. Biochem. 2013, 68, 1–7. [Google Scholar] [CrossRef]
- Hussain, I.; Ashraf, M.A.; Rasheed, R.; Asghar, A.; Sajid, M.A.; Iqbal, M. Exogenous application of silicon at the boot stage decreases accumulation of cadmium in wheat (Triticum aestivum L.) grains. Braz. J. Bot. 2015, 38, 223–234. [Google Scholar] [CrossRef]
- Bhatti, K.; Anwar, S.; Nawaz, K.; Hussain, K.; Siddiqi, E.; Sharif, R.; Talat, A.; Khalid, A. Effect of heavy metal lead (Pb) stress of different concentration on wheat (Triticum aestivum L.). Middle-East J. Sci. Res. 2013, 14, 148–154. [Google Scholar]
- Lukačová, Z.; Švubová, R.; Kohanová, J.; Lux, A. Silicon mitigates the Cd toxicity in maize in relation to cadmium translocation, cell distribution, antioxidant enzymes stimulation and enhanced endodermal apoplasmic barrier development. Plant Growth Regul. 2013, 70, 89–103. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Kumar, D.; Chauhan, D.K. Impact of exogenous silicon addition on chromium uptake, growth, mineral elements, oxidative stress, antioxidant capacity, and leaf and root structures in rice seedlings exposed to hexavalent chromium. Acta Physiol. Plant. 2012, 34, 279–289. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q.; Liu, C.; Wu, L. Silicon alleviates cadmium toxicity in peanut plants in relation to cadmium distribution and stimulation of antioxidative enzymes. Plant Growth Regul. 2010, 61, 45–52. [Google Scholar] [CrossRef]
- Li, P.; Song, A.; Li, Z.; Fan, F.; Liang, Y. Silicon ameliorates manganese toxicity by regulating manganese transport and antioxidant reactions in rice (Oryza sativa L.). Plant Soil 2012, 354, 407–419. [Google Scholar] [CrossRef]
- Song, A.; Li, Z.; Zhang, J.; Xue, G.; Fan, F.; Liang, Y. Silicon-enhanced resistance to cadmium toxicity in Brassica chinensis L. is attributed to Si-suppressed cadmium uptake and transport and Si-enhanced antioxidant defense capacity. J. Hazard. Mater. 2009, 172, 74–83. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The controversies of silicon’s role in plant biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Vlamis, J. The effect of silicon on yield and manganese-54 uptake and distribution in the leaves of barley plants grown in culture solutions. Plant Physiol. 1957, 32, 404–409. [Google Scholar] [CrossRef]
- Keller, C.; Rizwan, M.; Davidian, J.-C.; Pokrovsky, O.; Bovet, N.; Chaurand, P.; Meunier, J.-D. Effect of silicon on wheat seedlings (Triticum turgidum L.) grown in hydroponics and exposed to 0 to 30 µM Cu. Planta 2015, 241, 847–860. [Google Scholar] [CrossRef]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A transporter regulating silicon distribution in rice shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef]
- Naeem, A.; Ghafoor, A.; Farooq, M. Suppression of cadmium concentration in wheat grains by silicon is related to its application rate and cadmium accumulating abilities of cultivars. J. Sci. Food Agric. 2015, 95, 2467–2472. [Google Scholar] [CrossRef]
- Vaculík, M.; Landberg, T.; Greger, M.; Luxová, M.; Stoláriková, M.; Lux, A. Silicon modifies root anatomy, and uptake and subcellular distribution of cadmium in young maize plants. Ann. Bot. 2012, 110, 433–443. [Google Scholar] [CrossRef]
- Dragišić Maksimović, J.; Mojović, M.; Maksimović, V.; Römheld, V.; Nikolic, M. Silicon ameliorates manganese toxicity in cucumber by decreasing hydroxyl radical accumulation in the leaf apoplast. J. Exp. Bot. 2012, 63, 2411–2420. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Maier, P.; Fecht, M.; Horst, W.J. Leaf apoplastic silicon enhances manganese tolerance of cowpea (Vigna unguiculata). J. Plant Physiol. 2002, 159, 167–173. [Google Scholar] [CrossRef]
- Rogalla, H.; Römheld, V. Role of leaf apoplast in silicon-mediated manganese tolerance of Cucumis sativus L. Plant Cell Environ. 2002, 25, 549–555. [Google Scholar] [CrossRef]
- Kidd, P.; Llugany, M.; Poschenrieder, C.; Gunse, B.; Barcelo, J. The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in three varieties of maize (Zea mays L.). J. Exp. Bot. 2001, 52, 1339–1352. [Google Scholar] [PubMed]
- Barcelo, J.; Guevara, P.; Poschenrieder, C. Silicon amelioration of aluminium toxicity in teosinte (Zea mays L. ssp. mexicana). Plant Soil 1993, 154, 249–255. [Google Scholar] [CrossRef]
- Collin, B.; Doelsch, E.; Keller, C.; Cazevieille, P.; Tella, M.; Chaurand, P.; Panfili, F.; Hazemann, J.-L.; Meunier, J.-D. Evidence of sulfur-bound reduced copper in bamboo exposed to high silicon and copper concentrations. Environ. Pollut. 2014, 187, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Schaller, J.; Faucherre, S.; Joss, H.; Obst, M.; Goeckede, M.; Planer-Friedrich, B.; Peiffer, S.; Gilfedder, B.; Elberling, B. Silicon increases the phosphorus availability of Arctic soils. Sci. Rep. 2019, 9, 449. [Google Scholar] [CrossRef]
- Bolan, N.S.; Adriano, D.C.; Naidu, R. Role of phosphorus in (im) mobilization and bioavailability of heavy metals in the soil-plant system. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin, Germany, 2003; pp. 1–44. [Google Scholar]
- Gupta, D.; Chatterjee, S.; Datta, S.; Veer, V.; Walther, C. Role of phosphate fertilizers in heavy metal uptake and detoxification of toxic metals. Chemosphere 2014, 108, 134–144. [Google Scholar] [CrossRef]
- Rea, P.A. Phytochelatin synthase: Of a protease a peptide polymerase made. Physiol. Plant. 2012, 145, 154–164. [Google Scholar] [CrossRef]
- Li, J.; Leisner, S.M.; Frantz, J. Alleviation of copper toxicity in Arabidopsis thaliana by silicon addition to hydroponic solutions. J. Am. Soc. Hortic. Sci. 2008, 133, 670–677. [Google Scholar] [CrossRef]
- Khandekar, S.; Leisner, S. Soluble silicon modulates expression of Arabidopsis thaliana genes involved in copper stress. J. Plant Physiol. 2011, 168, 699–705. [Google Scholar] [CrossRef]
- Ma, J.; Cai, H.; He, C.; Zhang, W.; Wang, L. A hemicellulose-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells. New Phytol. 2015, 206, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Sonah, H.; Deshmukh, R.K.; Labbé, C.; Bélanger, R.R. Analysis of aquaporins in Brassicaceae species reveals high-level of conservation and dynamic role against biotic and abiotic stress in canola. Sci. Rep. 2017, 7, 2771. [Google Scholar] [CrossRef] [PubMed]
- Shivaraj, S.; Deshmukh, R.; Bhat, J.A.; Sonah, H.; Bélanger, R.R. Understanding Aquaporin Transport System in Eelgrass (Zostera marina L.), an Aquatic Plant Species. Front. Plant Sci. 2017, 8, 1334. [Google Scholar] [CrossRef] [PubMed]
- Shivaraj, S.; Deshmukh, R.K.; Rai, R.; Bélanger, R.; Agrawal, P.K.; Dash, P.K. Genome-wide identification, characterization, and expression profile of aquaporin gene family in flax (Linum usitatissimum). Sci. Rep. 2017, 7, 46137. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S.M.; Mahajan, R.; Bhat, J.A.; Nazir, M.; Deshmukh, R. Role of silicon in plant stress tolerance: Opportunities to achieve a sustainable cropping system. 3 Biotech 2019, 9, 73. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, X.; Lin, W.; Wang, G.; Cai, K. Transcriptome Analysis Reveals New Insights into the Bacterial Wilt Resistance Mechanism Mediated by Silicon in Tomato. Int. J. Mol. Sci. 2019, 20, 761. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef]
- Bharwana, S.; Ali, S.; Farooq, M.; Iqbal, N.; Abbas, F.; Ahmad, M. Alleviation of lead toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. J. Bioremed. Biodeg 2013, 4, 10.4172. [Google Scholar]
- Ali, S.; Farooq, M.A.; Yasmeen, T.; Hussain, S.; Arif, M.S.; Abbas, F.; Bharwana, S.A.; Zhang, G. The influence of silicon on barley growth, photosynthesis and ultra-structure under chromium stress. Ecotoxicol. Environ. Saf. 2013, 89, 66–72. [Google Scholar] [CrossRef]
- Doncheva, S.; Poschenrieder, C.; Stoyanova, Z.; Georgieva, K.; Velichkova, M.; Barceló, J. Silicon amelioration of manganese toxicity in Mn-sensitive and Mn-tolerant maize varieties. Environ. Exp. Bot. 2009, 65, 189–197. [Google Scholar] [CrossRef]
- da Cunha, K.P.V.; do Nascimento, C.W.A. Silicon effects on metal tolerance and structural changes in maize (Zea mays L.) grown on a cadmium and zinc enriched soil. Water Air Soil Pollut. 2009, 197, 323. [Google Scholar] [CrossRef]
- Vatehová, Z.; Kollárová, K.; Zelko, I.; Richterová-Kučerová, D.; Bujdoš, M.; Lišková, D. Interaction of silicon and cadmium in Brassica juncea and Brassica napus. Biologia 2012, 67, 498–504. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T.; Vaculik, M.; Lux, A. Silicon influences nutrient status in plants. In Proceedings of the 5th International Conference on Silicon in Agriculture, Beijing, China, 13–18 September 2011; The Organizing Committee of the 5th Silicon in Agriculture Conference: Beijing, China, 2011. [Google Scholar]
- Neumann, D.; zur Nieden, U.; Schwieger, W.; Leopold, I.; Lichtenberger, O. Heavy metal tolerance of Minuartia verna. J. Plant Physiol. 1997, 151, 101–108. [Google Scholar] [CrossRef]
- Neumann, D.; Zur Nieden, U. Silicon and heavy metal tolerance of higher plants. Phytochemistry 2001, 56, 685–692. [Google Scholar] [CrossRef]
- Rizwan, M.; Meunier, J.-D.; Miche, H.; Keller, C. Effect of silicon on reducing cadmium toxicity in durum wheat (Triticum turgidum L. cv. Claudio W.) grown in a soil with aged contamination. J. Hazard. Mater. 2012, 209, 326–334. [Google Scholar] [CrossRef]
- Dresler, S.; Wójcik, M.; Bednarek, W.; Hanaka, A.; Tukiendorf, A. The effect of silicon on maize growth under cadmium stress. Russ. J. Plant Physiol. 2015, 62, 86–92. [Google Scholar] [CrossRef]
- Ma, J.F.; Goto, S.; Tamai, K.; Ichii, M. Role of root hairs and lateral roots in silicon uptake by rice. Plant Physiol. 2001, 127, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Savant, N.K.; Datnoff, L.E.; Snyder, G.H. Depletion of plant-available silicon in soils: A possible cause of declining rice yields. Commun. Soil Sci. Plant Anal. 1997, 28, 1245–1252. [Google Scholar] [CrossRef]
- Meunier, J.; Guntzer, F.; Kirman, S.; Keller, C. Terrestrial plant-Si and environmental changes. Mineral. Mag. 2008, 72, 263–267. [Google Scholar] [CrossRef]
- Desplanques, V.; Cary, L.; Mouret, J.-C.; Trolard, F.; Bourrié, G.; Grauby, O.; Meunier, J.-D. Silicon transfers in a rice field in Camargue (France). J. Geochem. Explor. 2006, 88, 190–193. [Google Scholar] [CrossRef]
- Datnoff, L.; Deren, C.; Snyder, G. Silicon fertilization for disease management of rice in Florida. Crop Prot. 1997, 16, 525–531. [Google Scholar] [CrossRef]
- Henriet, C.; Draye, X.; Oppitz, I.; Swennen, R.; Delvaux, B. Effects, distribution and uptake of silicon in banana (Musa spp.) under controlled conditions. Plant Soil 2006, 287, 359–374. [Google Scholar] [CrossRef]
- Tubaña, B.S.; Heckman, J.R. Silicon in soils and plants. In Silicon and Plant Diseases; Springer: Berlin, Germany, 2015; pp. 7–51. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- Jarosz, Z. The effect of silicon application and type of substrate on yield and chemical composition of leaves and fruit of cucumber. J. Elem. 2013, 18, 403–414. [Google Scholar] [CrossRef]
- Ouellette, S.; Goyette, M.-H.; Labbé, C.; Laur, J.; Gaudreau, L.; Gosselin, A.; Dorais, M.; Deshmukh, R.K.; Bélanger, R.R. Silicon transporters and effects of silicon amendments in strawberry under high tunnel and field conditions. Front. Plant Sci. 2017, 8, 949. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.; Song, A.; Fan, F.; Li, Z.; Liang, Y. Effects of slag-based silicon fertilizer on rice growth and brown-spot resistance. PLoS ONE 2014, 9, e102681. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek, D.; Sommer, M. Silikon cycle in terrestrial biogeosystems of temperate climate. Soil Sci. Annu. 2004, 55, 221–230. [Google Scholar]
- Höhn, A.; Sommer, M.; Kaczorek, D.; Schalitz, G.; Breuer, J. Silicon fractions in histosols and gleysols of a temperate grassland site. J. Plant Nutr. Soil Sci. 2008, 171, 409–418. [Google Scholar] [CrossRef]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Mitani, N.; Chiba, Y.; Yamaji, N.; Ma, J.F. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009, 21, 2133–2142. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Gautam, A.; Dubey, R.S. Transport and detoxification of metalloids in plants in relation to plant-metalloids tolerance. Plant Gene 2019, 17, 1–7. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guérin, V.; Carpentier, G.; Sonah, H.; Labbé, C.; Isenring, P.; Belzile, F.J.; Bélanger, R.R. A precise spacing between the NPA domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Deren, C. Plant genotype, silicon concentration, and silicon-related responses. In Studies in Plant Science; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 149–158. [Google Scholar]
- Ma, J.F.; Higashitani, A.; Sato, K.; Takeda, K. Genotypic variation in silicon concentration of barley grain. Plant Soil 2003, 249, 383–387. [Google Scholar]
- Ma, J.F.; Yamaji, N.; Tamai, K.; Mitani, N. Genotypic difference in silicon uptake and expression of silicon transporter genes in rice. Plant Physiol. 2007, 145, 919–924. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Crop Species | Heavy Metal | Phytotoxicity | References # |
|---|---|---|---|
| Rice | Hg | Reduction in grain yield; reduced tiller and panicle formation; decrease in plant height | S1 |
| As | Reduction in seed germination and seedling height; reduced leaf area and dry matter production | S2 | |
| Cd | Inhibition of root growth; The appearance of black spots in the cortex and pericycle | S3 | |
| Wheat | Pb | Reduced shoot/root length, shoot fresh/dry weights, number of tillers; decreased photosynthetic pigments such as chla and chlb | S4 |
| Pb | The decrease in the contents of Chla, Chlb and proline content; growth inhibition and a decrease in dry weights of plant parts | S5 | |
| Ni | Reduction in the total shoot and root lengths; decrease in Chla and Chlb contents and enhancement of chlorophyll a/b ratio | S6 | |
| Cd | Reduction in seed germination; decrease in plant nutrient content; reduced shoot and root length | S7 | |
| Cr | Inhibition of shoot and root growth | S8 | |
| Ni | Reduction in plant nutrient acquisition | S9 | |
| Maize | Cd & Zn | Reduction in shoot/root biomass; decrease in length as well as dry weights of shoot/root | S10 |
| Cd & Ni | Inhibition of root growth; reduction in the length of primary roots | S11 | |
| Cd | Inhibition of shoot and root growth | S12 | |
| Barley | Cd | Reduction of Chla, Chlb, and carotenoids; decreased plant growth and biomass | S13 |
| Cu & Cd | Reduction of plant dry weights, root length, and shoot height; alteration of photosynthetic pigments and lipid peroxidation | S14 | |
| Cd & Zn | Reduction of total biomass; decrease in the length of roots and leaves | S15 | |
| Sorghum | Cu & Cd | The decrease in root diameter, width, and thickness of leaf midrib and diameter of xylem vessels; reduction of yield and yield contributing traits | S16 |
| Cd | The decrease in chlorophyll pigments, plant growth, and root characteristics | S17 | |
| Oat | Pd | Inhibition of enzyme activity which affected CO2 fixation | S18 |
| Soybean | Hg | Reduction of oil content; changes in major and minor fatty acid concentration of soybean seed | S19 |
| Cd | The decrease in the accumulated amount of dry matter as well as the content of mineral elements; inhibition of chlorophyll biosynthesis; decrease in the Mg uptake | S20 | |
| Co & Pb | Reduced seed germination, seedling growth, leaf area, root development, and biomass production | S21 | |
| Cd | Reduction of nodulation and leghaemoglobin content; a decrease in crop productivity | S22 | |
| Bean | Cu | Root malformation and accumulation of Cu in plant roots; reduction of root growth | S23 |
| Zn | The decrease in plant growth, development, and metabolism; induction of oxidative damage | S24 | |
| Zn | Reduction of photosynthetic pigments including Chla and Chlb; disruption of absorption and translocation of Fe and Mg into the chloroplast | S25 | |
| Chickpea | Cd | Inhibition of seed germination and root growth; disruption of vascular tissues as well as associated tissues | S26 |
| Cu | Reduction of root/shoot length and RWC; lipid peroxidation | S27 | |
| Co | Inhibition of photosynthetic process and activity of antioxidative enzymes; The increase in proline content and lipid peroxidation | S28 | |
| Pigeonpea | Ni | The decrease in stomatal conductance and chlorophyll content; decreased enzyme activity which affected the Calvin cycle and CO2 fixation | S29 |
| Pd & Cd | Reduction in photosynthetic activity; decrease in chlorophyll content and stomatal conductance | S30 | |
| Hg & Cd | Reduction of germination percentage, root and shoot length, fresh and dry weight of seedlings; inhibition of root elongation | S31 | |
| Faba bean | Mn | Mn accumulation in shoot and root; reduction of the shoot and root length; chlorosis | S32 |
| Cd | Cd accumulation in shoot and root; decrease in photosynthetic pigments as well as root fresh and dry weights | S33 | |
| Pea | Mn | The decrease in Chla and Chlb content; reduction in relative growth rate; reduced photosynthetic O2 evolution activity and photosystem II activity | S34 |
| Zn | Reduction in chlorophyll content; alteration in the structure of chloroplast; reduction in photosystem II activity; reduced plant growth | S35 | |
| Mung bean | Co | Reduction in antioxidant enzyme activities; decrease in plant sugar, starch, amino acids, and protein content | S36 |
| Groundnut | Cd, Pb & As | Reduction in germination rate, root length, shoot length, biomass and seedling vigor index | S37 |
| Cr | Increase in lipid peroxidation; decrease in photosynthetic pigments; induced oxidative stress | S38 | |
| Ni | Reduction of seed germination, root/shoot growth and fresh weight of seedlings | S39 | |
| Cotton | Cd | The decrease in plant height, biomass and leaf area; reduction of Chla, Chlb, photosynthetic rate, stomatal conductance, and transpiration rate | S40 |
| Pb | The decrease in net photosynthetic rate, stomatal conductance, transpiration rate, water use efficiency, chlorophyll, carotenoids, and the Soil Plant Analysis Development (SPAD) chlorophyll meter value | S41 | |
| Tomato | As | The decrease in leaf fresh weight; Reduced fruit yield | S42 |
| Co | Reduction in plant nutrient content | S43 | |
| Cr | Decrease in plant nutrient acquisition | S44 | |
| Hg | Reduction in germination percentage; reduced plant height; reduction in flowering and fruit weight; chlorosis | S45 | |
| Canola | As | Wilting, chlorosis and stunted growth | S46 |
| Brassica juncea | Cd & Pb | Reduction of growth and biomass yield; decrease of chlorophyll and carotenoid pigments | S47 |
| Garlic | Cd | Reduced shoot growth; Cd accumulation | S48 |
| Onion | Cr | Inhibition of germination process; reduction of plant biomass | S49 |
| Radish | Co | Decrease in shoot and root length as well as total leaf area; reduction in chlorophyll content, plant nutrient content and antioxidant enzyme activity | S50 |
| Crop | Heavy Metal | Mechanisms | References # |
|---|---|---|---|
| Rice | Cd, Zn, Cu & Pb | Immobilization of heavy metals in culture media and decrease of phytoavailability which further suppressed metal uptake | S51 |
| As | Overcomes heavy metal uptake by competes with arsenate ions for root entry point | S52 | |
| Zn | Strong binding of Zn in the cell wall of less bioactive tissues, especially in sclerenchyma of root | S51 | |
| Pb | Preventing Pb transfer from rice roots to aboveground parts, and blocking Pb accumulation in rice grains | S53 | |
| Cd | Si bound to cell wall inhibits apoplastic Cd uptake by covalently bonding with Cd and trapping Cd as it diffuses through the cell wall and intracellular spaces. | S54 | |
| Cd | Increased the activities of antioxidant enzymes and preventing membrane oxidative damage of plant tissue | S55 | |
| Cd | Decreased Cd accumulation in rice shoots by compartmentalization of Cd in the root cell walls | S56 | |
| Maize | Cd & Zn | Increased diameter of xylem, thickness of leaf mesophyll and epidermis, and transversal area occupied by collenchyma and mid vein | S57 |
| Al | Formation of hydroxyaluminosilicates in the apoplast of the root apex reducing the mobility of apoplastic Al | S58 | |
| Cd | Formation of colloidal silicon in cell walls which has high specific adsorption property to Cd preventing Cd uptake into the cell | S59 | |
| Cd | Cd co-precipitation with silicates, resulting in strong binding of Cd to cell walls, thereby reducing the concentration of Cd in the symplast | S60 | |
| Zn | Formation of less soluble zinc-silicates in the cytoplasm | S61 | |
| Wheat | Cd | Enhanced antioxidant enzymes activities, and preventing lipid peroxidation as well as membrane oxidative damage of plant tissue | S62 |
| Cd | Decreased Cd uptake as well as translocation of Cd to shoots and grains | S63 | |
| Cu | Cu form complex with organic acids and reduced the Cu translocation to shoots | S64 | |
| Cd | Formation of apoplasmic barriers in endodermis closer to the wheat root apex | S65 | |
| Barley | Al | Exclusion of Al from the subtending tissue as a result of silicon deposition at the epidermis, restricting total overall Al uptake into the root | S66 |
| Cr | Increased plant height, number of tillers, root length and leaf size of barley plants | S67 | |
| Cucumber | Mn | Reduction of lipid peroxidation, and increase of enzymatic and non-enzymatic antioxidants levels | S68 |
| Mn | Si increased Mn fraction in the cell wall of shoots, thereby reduced Mn concentration of symplast | S69 | |
| Cowpea | Mn | Si reduced the apoplastic Mn concentration and modify the cation binding capacity of the cell wall | S70 |
| Mn | Enhanced adsorption of Mn on cell walls reducing the amount of soluble apoplastic Mn | S71 | |
| Mn | Co-precipitation of Si and Mn in leaf apoplast of cowpea plants, and increases Mn fraction in the cell wall of shoots | S71 | |
| Peanut | Cd | Increased activities of antioxidant enzymes; inhibition of Cd transport from roots to shoots | S72 |
| Arabidopsis | Cu | Stimulated the genes responsible for the production of metallothioneins (MTs) that can chelate toxic metals | S73 |
| Rapeseed | Cd | Mediated extensive development of suberin lamellae in endoderm closer to the root tips | S74 |
| Minuartia verna | Zn | Co-precipitation as Zn silicates in the cell walls of leaf epidermis | S75 |
| Cardaminopsis halleri | Zn | Formation of Si–Zn complexes in leaves surface of Cardaminopsishalleri grown on a Zn contaminated soil amended with Si. | S76 |
| Pakchoi | Cd | Increased activities of enzymatic and non-enzymatic antioxidants levels, protein thiols (NPT) and ascorbic acid | S77 |
| Cotton | Cd | Enhanced activities of antioxidant enzymes as well as reduced electrolytic leakage, malondialdehyde and hydrogen peroxide contents, thereby preventing plant tissue from oxidative damage | S78 |
| Pb | Increased the activities of antioxidant enzymes and preventing membrane oxidative damage of plant tissue | S41 |
| Crop Species | Transporter | Type | Expression Site | References # |
|---|---|---|---|---|
| Maize | ZmLsi1 | Influx | Root | S79 |
| ZmLsi6 | Influx | Leaf Sheaths and blades | S80 | |
| ZmLsi2 | Efflux | Roots | S80 | |
| Rice | OsLsi1 | Influx | Roots | S81 |
| OsLsi2 | Efflux | Roots | S82 | |
| OsLsi6 | Influx | Leaf | S83 | |
| OsLsi3 | Influx | Panicles | S84 | |
| Barley | HvLsi2 | Efflux | Root | S80 |
| HvLsi1 | Influx | Root | S85 | |
| HvLsi6 | Influx | Leaf Sheaths and blades | S86 | |
| Soybean | GmNIP2-1 | Influx | Root and shoot | S87 |
| GmNIP2-2 | Influx | Root and shoot | S87 | |
| Wheat | TaLsi1 | Influx | Root | S88 |
| Pumpkin | CmLsi1 | Influx | Root and shoot | S89 |
| CmLsi2 | Efflux | Root and shoot | S89 | |
| Horsetail | EaNIP3;1 | Influx | Root and shoot | S90 |
| EaNIP3;3, | Influx | Root and shoot | S90 | |
| EaLsi2-1 | Efflux | Root and shoot | S91 | |
| EaLsi2-2 | Efflux | Root and shoot | S91 | |
| Potato | StLsi1 | Influx | Root and leaves | S92 |
| Tomato | SlNIP2-1 (V140del) * | Influx | Root and leaves | S93 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants 2019, 8, 71. https://doi.org/10.3390/plants8030071
Bhat JA, Shivaraj SM, Singh P, Navadagi DB, Tripathi DK, Dash PK, Solanke AU, Sonah H, Deshmukh R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants. 2019; 8(3):71. https://doi.org/10.3390/plants8030071
Chicago/Turabian StyleBhat, Javaid Akhter, S. M. Shivaraj, Pritam Singh, Devanna B. Navadagi, Durgesh Kumar Tripathi, Prasanta K. Dash, Amolkumar U. Solanke, Humira Sonah, and Rupesh Deshmukh. 2019. "Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants" Plants 8, no. 3: 71. https://doi.org/10.3390/plants8030071