
Photosynthesis Performance at Different Growth Stages, Growth, and Yield of Rice in Saline Fields
 ,
,
Abstract
:1. Introduction
2. Results
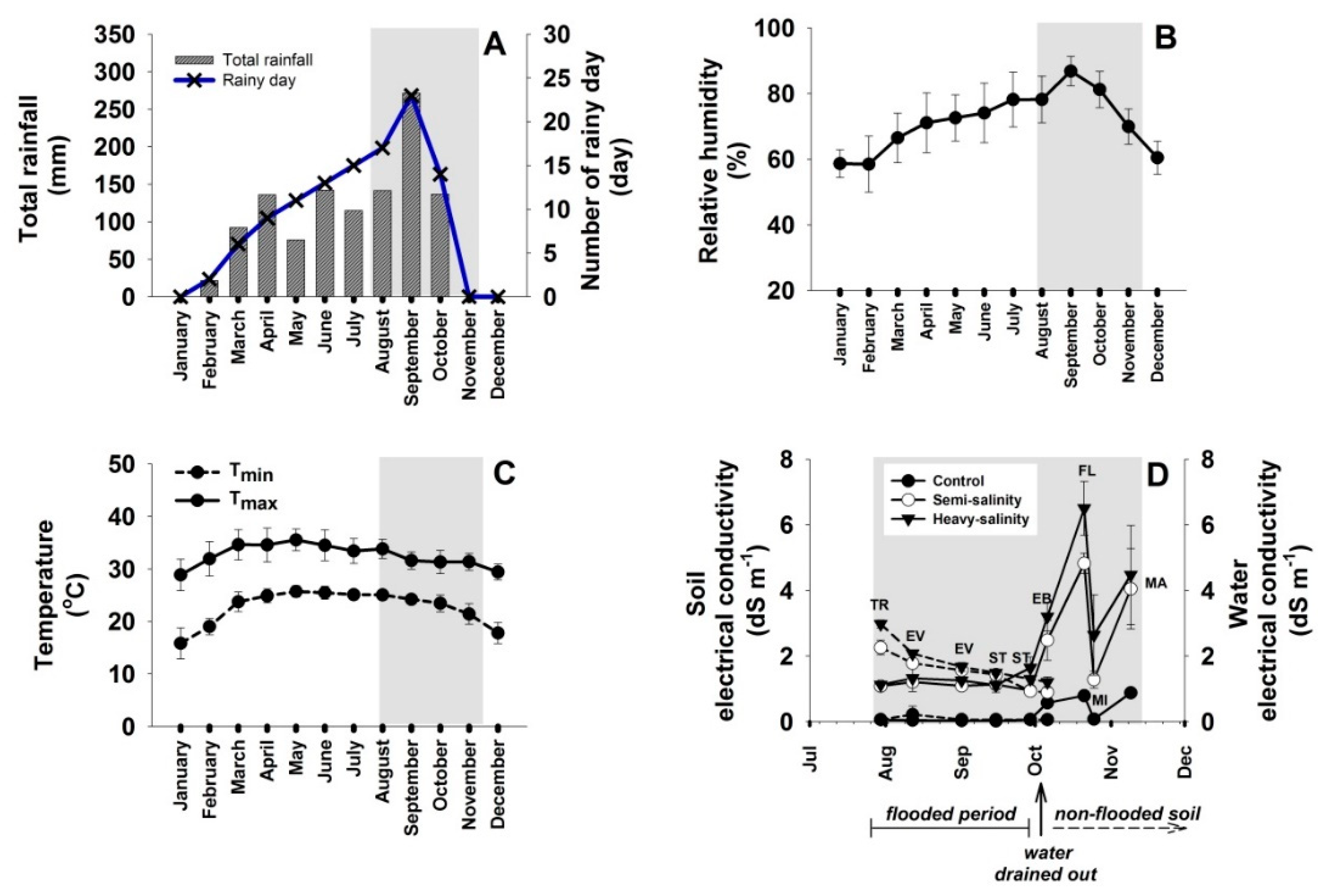
2.1. Environmental Conditions in the Area
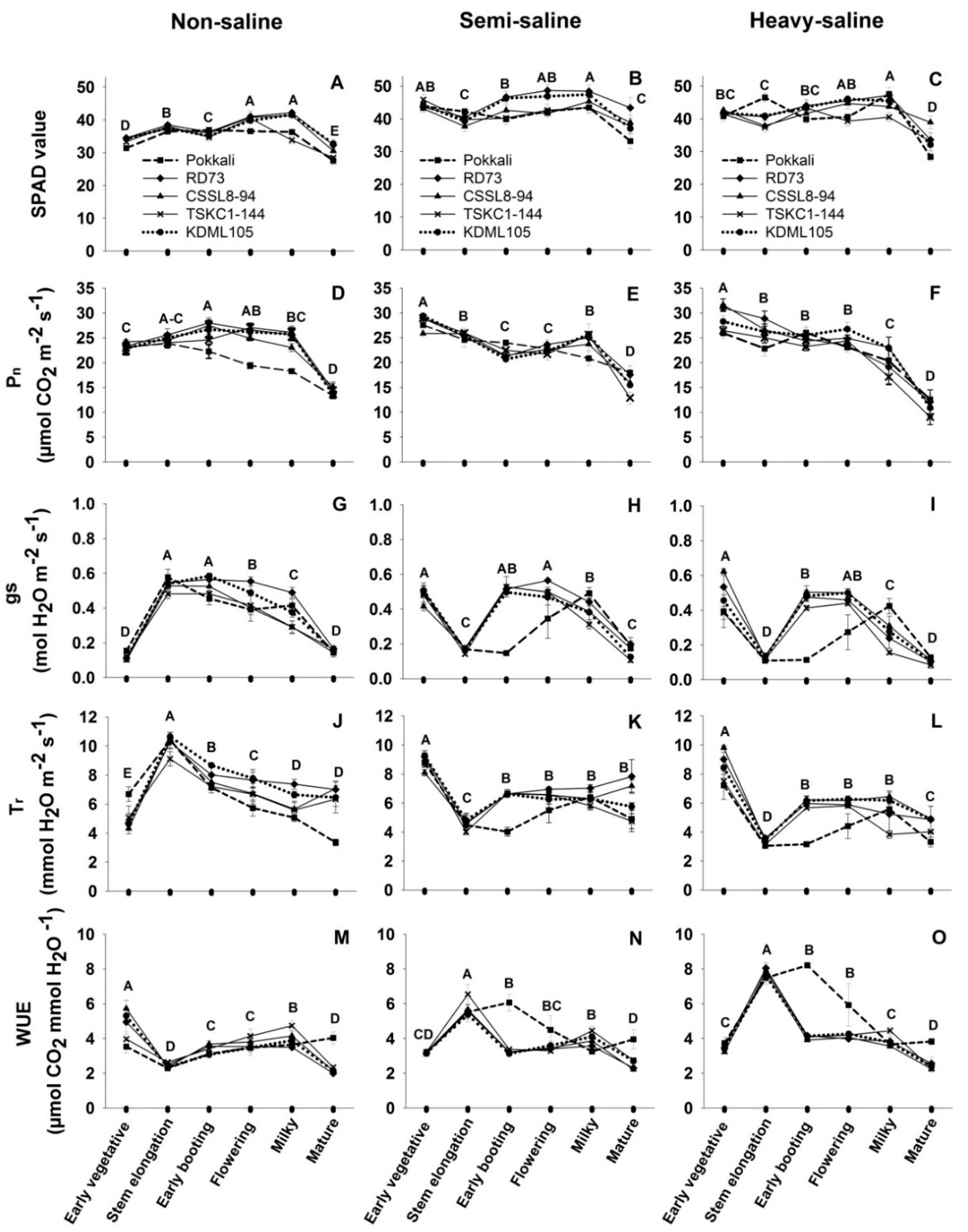
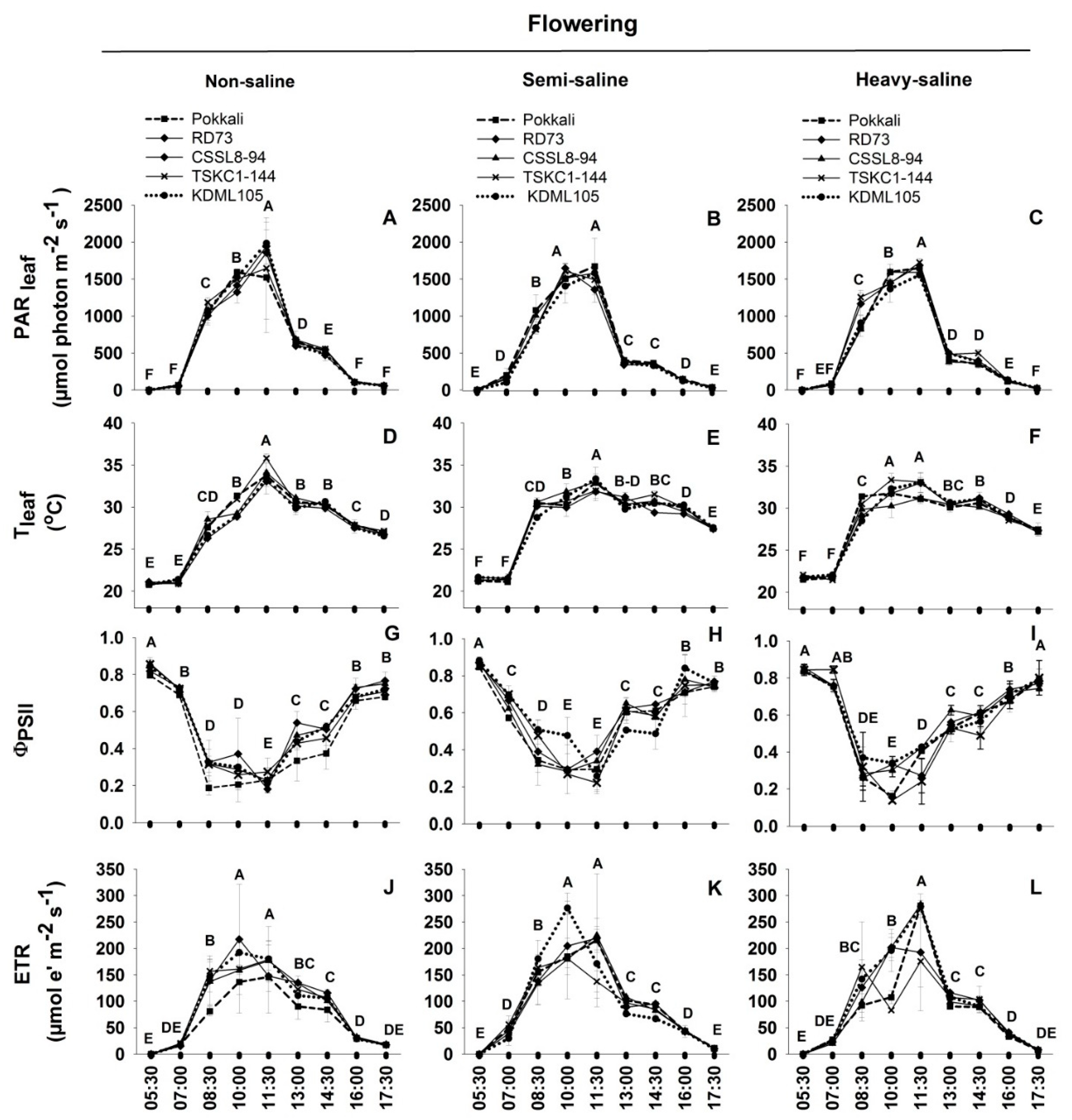
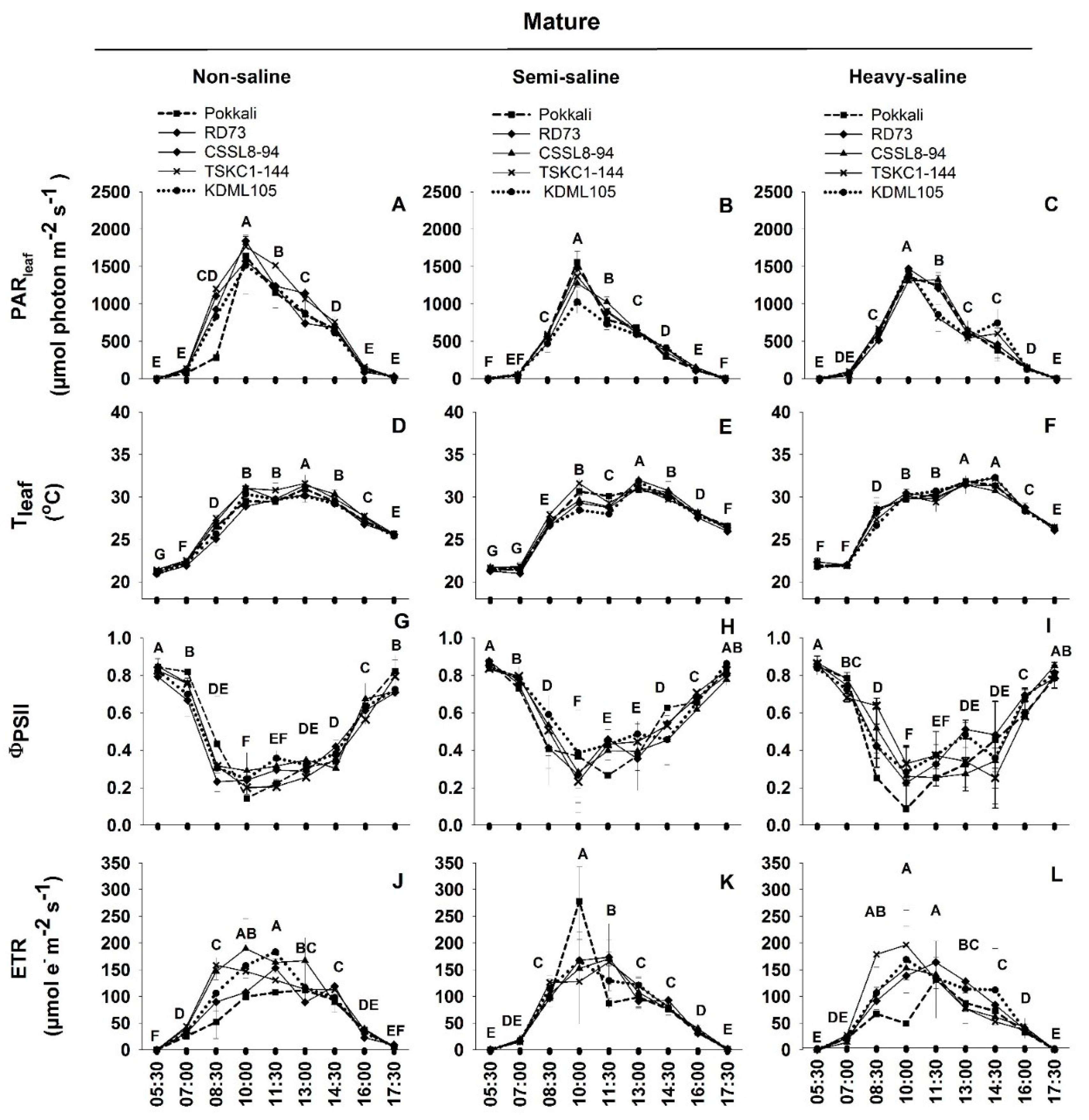
2.2. Photosynthetic Performance
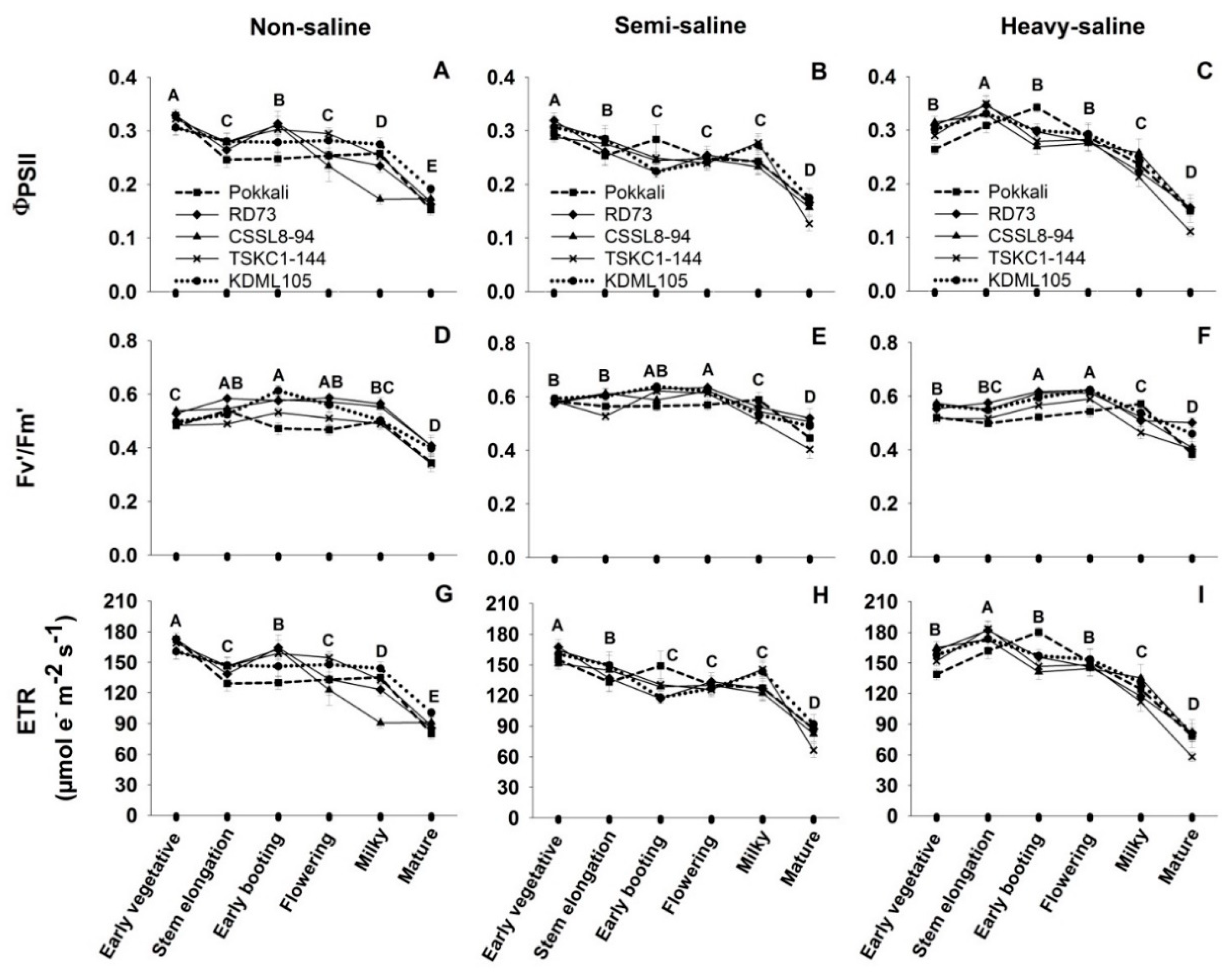
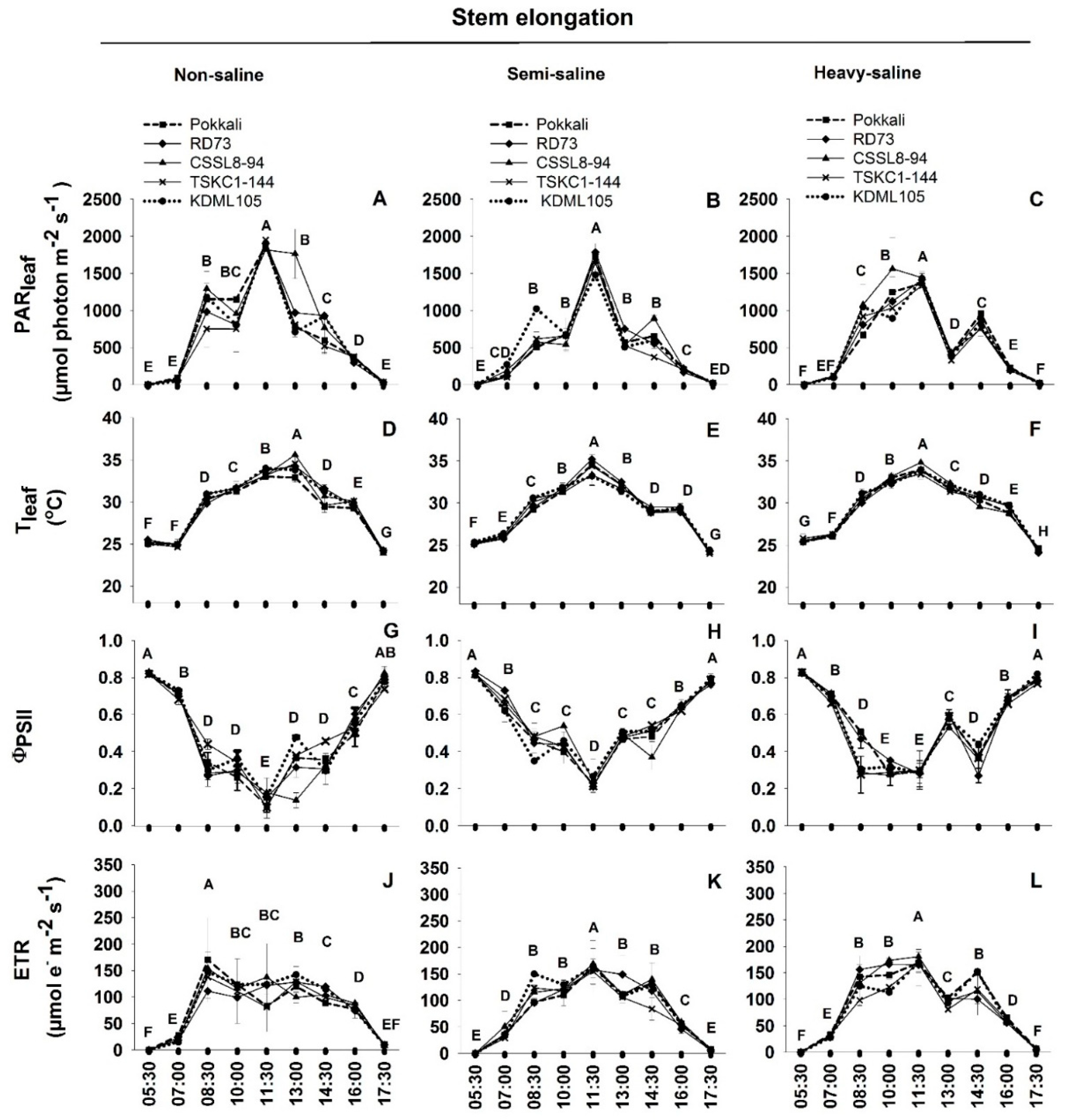
2.3. Diurnal Chlorophyll Fluorescence
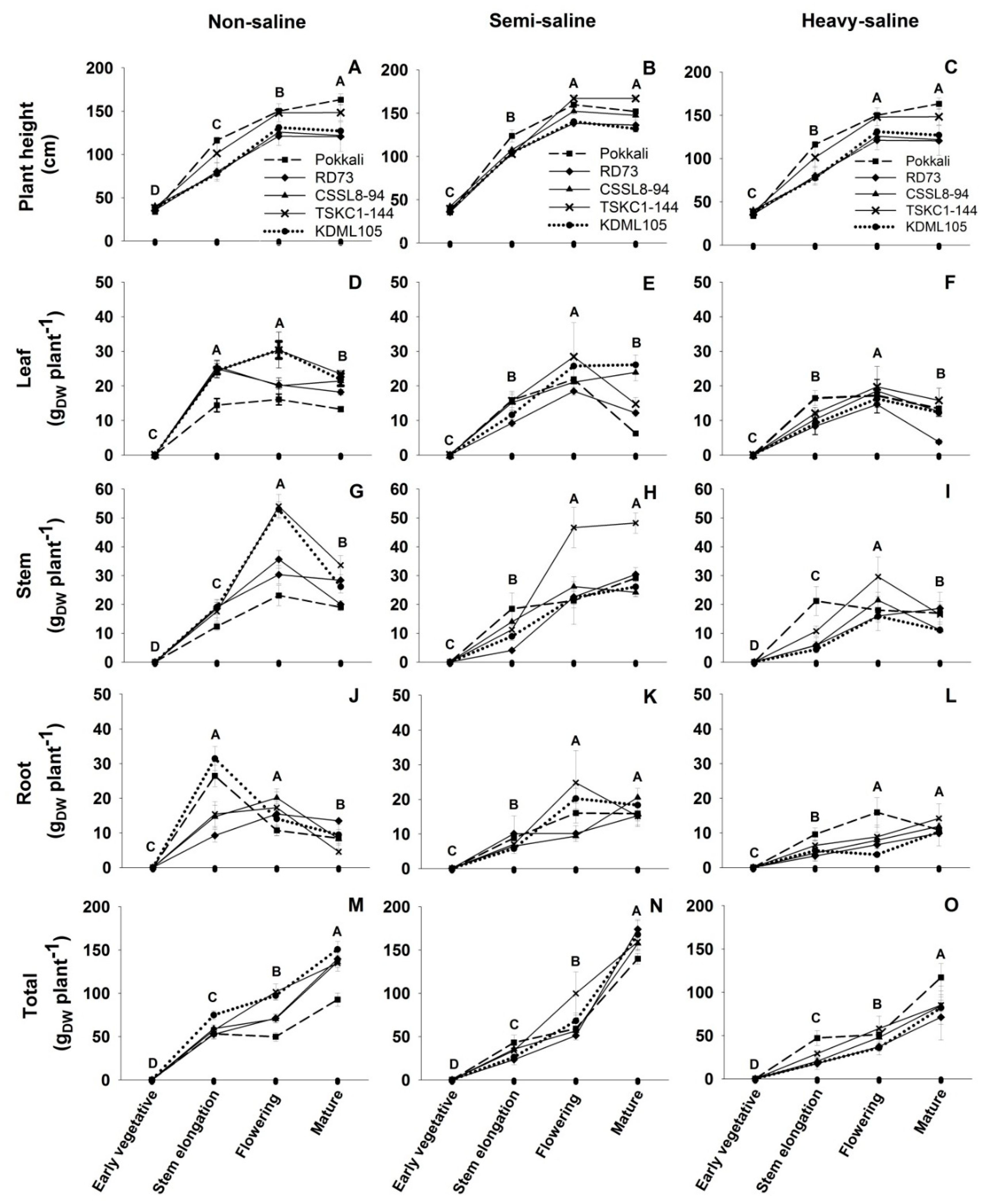
2.4. Growth and Biomass
2.5. Sodium (Na+) and Potassium (K+) Content and Yield
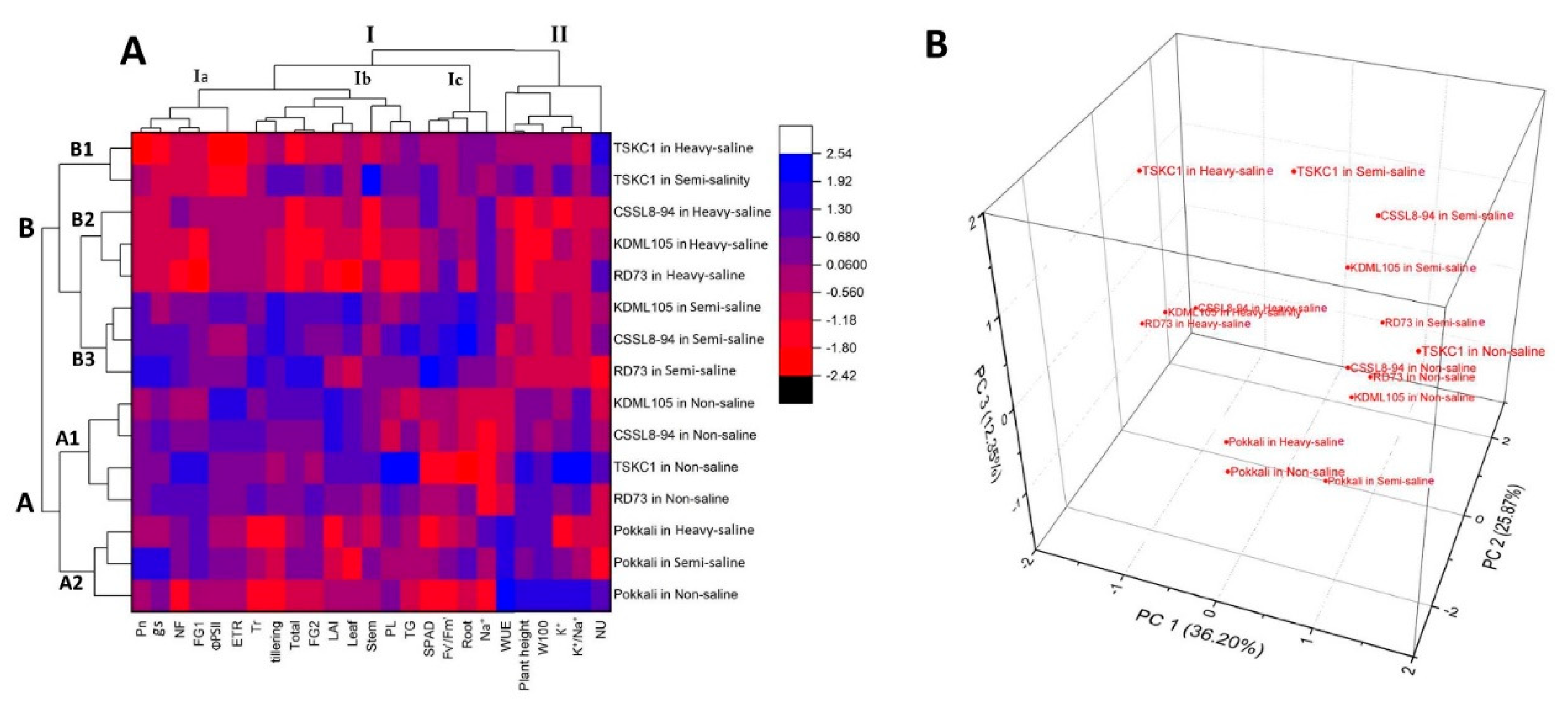
2.6. Principal Component Analysis (PCA) and Hierarchical Cluster Analysis (HCA) for Biomass, Yield, and Physiological Parameters
3. Discussion
4. Materials and Methods
4.1. Site Description and Microclimate
4.2. Plant Materials and Cultural Practice
4.3. Photosynthetic Performance
4.3.1. Leaf Chlorophyll Content, Leaf Gas Exchange and Chlorophyll Fluorescence
4.3.2. Diurnal Chlorophyll Fluorescence
4.4. Growth, Biomass and Yield
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 11 November 2022).
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for global nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Suebpongsang, P.; Ekasingh, B.; Cramb, R. Commercialisation of Rice Farming in Northeast Thailand. In White Gold the Commercialisation of Rice Farming in the Lower Mekong Basin; Cramb, R., Ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2019; pp. 39–68. [Google Scholar]
- Office of Agricultural Economics. Agricultural Statistics of Thailand 2019. Available online: https://www.opsmoac.go.th/saraburi-dwl-files-411191791833 (accessed on 10 November 2022).
- Food and Agriculture Organization of the United Nations (FAO). A Climate Change Impacts in Food Security: Overview of Latest Knowledge. In Climate Change and Food Security: Risks and Responses; FAO: Rome, Italy, 2015; pp. 3–34. [Google Scholar]
- Yadav, M.; Gupta, S.; Seth, C.S. Foliar application of α-lipoic acid attenuates cadmium toxicity on photosynthetic pigments and nitrogen metabolism in Solanum lycopersicum L. Acta Physiol. Plant 2022, 44, 112. [Google Scholar] [CrossRef]
- Gupta, S.; Seth, C.S. Salicylic acid alleviates chromium (VI) toxicity by restricting its uptake, improving photosynthesis and augmenting antioxidant defense in Solanum lycopersicum L. Physiol. Mol. Biol. Plants 2021, 27, 2651–2664. [Google Scholar] [CrossRef]
- Hussain, S.; Bai, Z.; Huang, J.; Cao, X.; Zhu, L.; Zhu, C.; Khaskheli, M.A.; Zhong, C.; Jin, Q.; Zhang, J. 1-Methylcyclopropene modulates physiological, biochemical, and antioxidant responses of rice to different salt stress levels. Front. Plant Sci. 2019, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Land Development Department (LDD). Soil Management. Available online: https://www.ldd.go.th/Web_Soil/salty.htm (accessed on 11 November 2022).
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance–current assessment. J. Irrig. Drain. Div. ASCE 1977, 103, 115–134. [Google Scholar] [CrossRef]
- Hussain, S.; Ahmad, S.; Hussain, S.; SamiUl-Allah, L.; Nawaz, A. Chapter Six-Rice in saline soils: Physiology, biochemistry, genetics, and management. Agronomy 2018, 148, 231–287. [Google Scholar]
- Gao, J.; Chao, D.; Lin, H. Understanding abiotic stress tolerance mechanisms: Recent studies on stress response in rice. J. Integr. Plant Biol. 2007, 49, 742–750. [Google Scholar] [CrossRef]
- Grattan, S.R.; Zeng, L.; Shannon, M.C.; Roberts, S.R. Rice is more sensitive to salinity than previously thought. Calif. Agric. 2002, 56, 189–198. [Google Scholar] [CrossRef]
- Umali, D.L. Irrigation-Induced Salinity a Growing Problem for Development and the Environment; The Word Bank: Washington, DC, USA, 1993; pp. 8–12. [Google Scholar]
- Asch, F.; Wopereis, M.C.S. Responses of field-grown irrigated rice cultivars to varying levels of floodwater salinity in a semi-arid environment. Field Crops Res. 2001, 70, 127–137. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Redoña, E.; Refuerzo, L. Varietal Improvement for Abiotic Stress Tolerance in Crop Plants: Special Reference to Salinity in Rice. In Abiotic Stress Adaptation in Plants; Springer: Dordrecht, The Netherlands, 2009; pp. 387–415. [Google Scholar]
- Khatun, S.; Rizzo, C.A.; Flowers, T.J. Genotypic variation in the effect of salinity on fertility in rice. Plant Soil 1995, 173, 239–250. [Google Scholar] [CrossRef]
- Rao, P.S.; Mishra, B.; Gupta, S.R. Effects of soil salinity and alkalinity on grain quality of tolerant, semi-tolerant and sensitive rice genotypes. Rice Sci. 2013, 20, 284–291. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhang, H.; Li, Z.; Shen, S.; Sun, J.; Wang, M.; Liao, D.; Liu, X.; Wang, X.; Xiao, F.; et al. Evaluation of genetic diversity of rice landraces (Oryza sativa L.) in Yunnan, China. Breed Sci. 2007, 57, 91–99. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.; Naidu, S.L.; Ort, D. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob. Chang. Biol. 2020, 27, 27–49. [Google Scholar] [CrossRef]
- Lekklar, C.; Suriya-Arunroj, D.; Pongpanich, M.; Comai, L.; Kositsup, B.; Chadchawan, S.; Buaboocha, T. Comparative genomic analysis of rice with contrasting photosynthesis and grain production under salt stress. Genes 2019, 10, 562. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef]
- Cha-um, S.; Charoenpanich, A.; Roytrakul, S.; Kirdmanee, C. Sugar accumulation, photosynthesis and growth of two indica rice varieties in response to salt stress. Acta Physiol. Plant 2009, 31, 477–486. [Google Scholar] [CrossRef]
- Pamuta, D.; Siangliw, M.; Sanitchon, J.; Pengrat, J.; Siangliw, J.L.; Toojinda, T.; Theerakulpisut, P. Photosynthetic performance in improved ‘KDML105’ rice (Oryza sativa L.) lines containing drought and salt tolerance genes under drought and salt stress. Pertanika J. Trop. Agric. Sci. 2020, 43, 653–675. [Google Scholar] [CrossRef]
- Kaur, N.; Dhawan, M.; Sharma, I.; Pati, P.K. Interdependency of Reactive Oxygen Species generating and scavenging system in salt sensitive and salt tolerant cultivars of rice. BMC Plant Biol. 2016, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Chen, K.C.; Cheng, T.S.; Lee, C.; Lin, S.; Tung, C. Chlorophyll fluorescence analysis in diverse rice varieties reveals the positive correlation between the seedlings salt tolerance and photosynthetic efficiency. BMC Plant Biol. 2019, 19, 403. [Google Scholar] [CrossRef] [PubMed]
- Makino, A. Photosynthesis improvement for enhancing productivity in rice. J. Soil Sci. Plant Nutr. 2021, 67, 513–519. [Google Scholar] [CrossRef]
- Nounjan, N.; Wuttipong Mahakham, W.; Siangliw, J.L.; Toojinda, T.; Theerakulpisut, P. Chlorophyll retention and high photosynthetic performance contribute to salinity tolerance in rice carrying drought tolerance quantitative trait Loci (QTLs). Agriculture 2020, 10, 620. [Google Scholar] [CrossRef]
- Zeng, L.; Shanon, M. Salinity effects on rice seedlings and yield components. Crop Sci. 2000, 40, 996–1003. [Google Scholar] [CrossRef]
- Sultana, N.; Ikeda, T.; Itoh, R. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. EEB 1999, 42, 211–220. [Google Scholar] [CrossRef]
- Singh, R.K.; Kota, S.; Flowers, T.J. Salt tolerance in rice: Seedling and reproductive stage QTL mapping come of age. Theor. Appl. Genet. 2021, 134, 3495–3533. [Google Scholar] [CrossRef]
- Gerona, M.E.B.; Deocampo, M.P.; Egdane, J.A.; Ismail, A.M.; Dionisio-Sese, M.L. Physiological responses of contrasting rice genotypes to salt stress at reproductive stage. Rice Sci. 2019, 26, 207–219. [Google Scholar] [CrossRef]
- Silas, W.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Photosynthesis and salinity: Are they mutually exclusive? Photosynthetica 2008, 56, 366–381. [Google Scholar]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Seth, C.S. 24-Epibrassinolide regulates functional components of nitric oxide signaling and antioxidant defense pathways to alleviate salinity stress in Brassica juncea L. cv. Varuna. J. Plant Growth Regul. 2022. [CrossRef]
- Asch, F.; Dingkuhn, M.; Dorffling, K.; Miezen, K. Leaf K/Na ratio predicts salinity induced yield loss in irrigated rice. Euphytica 2000, 113, 109–118. [Google Scholar] [CrossRef]
- Pal, M.; Singh, D.K.; Rao, L.S.; Singh, K.P. Photosynthetic characteristics and activity of antioxidant enzymes in salinity tolerant and sensitive rice cultivars. Indian J. Plant Physiol. 2004, 9, 407–412. [Google Scholar]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Kakar, N.; Jumaa, S.H.; Redoña, E.D.; Warburton, M.L.; Reddy, R.K. Evaluating rice for salinity using pot-culture provides a systematic tolerance assessment at the seedling stage. Rice 2019, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.H.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Alboresi, A.; Storti, M.; Morosinotto, T. Balancing protection and efficiency in the regulation of photosynthetic electron transport across plant evolution. New Phytol. 2019, 221, 105–109. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Miao, Y.; Cai, Y.; Wu, H.; Wang, D. Diurnal and seasonal variations in the photosynthetic characteristics and the gas exchange simulations of two rice cultivars grown at ambient and elevated CO2. Front. Plant Sci. 2021, 12, 651606. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Chen, Y.-Z.; Hubbart, S.; Peng, S.; Horton, P. Interactions between senescence and leaf orientation determine in situ patterns of photosynthesis and photoinhibition in field-grown rice. Plant Physiol. 1999, 119, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Chen, Y.; Li, W.; Chen, Y.; Ma, J.; Fu, A. Effects of groundwater decline on Populus euphratica at hyper-arid regions: The lower reaches of the tarim river in Xinjiang, China. FEB 2011, 20, 3326–3337. [Google Scholar]
- Yang, Y.; Ye, R.; Srisutham, M.; Nontasri, T.; Sritumboon, S.; Maki, M.; Yoshida, K.; Oki, K.; Koki Homma, K. Rice production in farmer fields in soil salinity classified areas in Khon Kaen, Northeast Thailand. Sustainability 2022, 14, 9873. [Google Scholar] [CrossRef]
- Park, H.; Seo, B.; Jeong, Y.; Yang, H.I.; Park, S.; Baek, N.; Jin-Hyeob Kwak, J.; Woo-Jung Choi, W. Soil salinity, fertility and carbon content, and rice yield of salt-affected paddy with different cultivation period in southwestern coastal area of South Korea. J. Soil Sci. Plant Nutr. 2022, 68, 53–63. [Google Scholar] [CrossRef]
- Ren, Z.; Gao, J.; Li, L.; Cai, X.; Huang, W.; Chao, D.; Zhu, M.; Wang, Z.; Luan, S.; Lin, H. Arice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Shohan, M.U.S.; Sinha, S.; Nabila, F.H.; Dastidar, S.G.; Seraj, I.Z. HKT1;5 transporter gene expression and association of amino acid substitutions with salt tolerance across rice genotypes. Front. Plant Sci. 2019, 10, 1420. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, A.; Chakraborty, K.; Mondal, S.; Jena, P.; Panda, R.K.; Samal, K.C.; Chattopadhyay, K. Relative contribution of ion exclusion and tissue tolerance traits govern the differential response of rice towards salt stress at seedling and reproductive stages. Environ. Exp. Bot. 2023, 206, 105131. [Google Scholar] [CrossRef]
- Prajapati, P.; Gupta, S.; Kharwar, R.N.; Seth, C.S. Nitric oxide mediated regulation of ascorbate-glutathione pathway alleviates mitotic aberrations and DNA damage in Allium cepa L. under salinity stress. Int. J. Phytoremediat. 2022, 25, 403–414. [Google Scholar] [CrossRef]
- Clemont–Dauphin, C.; Suwannang, N.; Grunberger, O.; Hammerker, C.; Maeght, J.L. Yield of rice under water and soil salinity risks in farmers’ fields in northeast Thailand. Field Crops Res. 2010, 118, 289–296. [Google Scholar] [CrossRef]
- Pamuta, D.; Siangliw, M.; Sanitchon, J.; Pengrat, J.; Siangliw, J.L.; Toojinda, T.; Theerakulpisut, P. Physio-biochemical traits in improved ‘KDML105’ jasmine rice lines containing drought and salt tolerance gene under drought and salt stress. Chil. J. Agric. Res. 2022, 82, 97–110. [Google Scholar] [CrossRef]
- Suriyaarunroj, D.; Khongsuwan, P.; Lertna, S.; Hintung, P.; Budjun, I.; Boonmee, N.; Saleetoe, S.; Jumpagate, R.; Sripanom, P.; Gaewkumjang, C.; et al. RD73, a non-glutinous rice variety. Thai Rice Res. J. 2018, 9, 25–50. [Google Scholar]
- Ezeaku, P.; Ene, J.; Shehu, J. Application of different reclamation methods on salt affected soils for crop production. Am. J. Exp. Agric. 2015, 9, 1–11. [Google Scholar] [CrossRef]
- Hussain, S.; Zhang, J.H.; Zhong, C.; Zhu, L.F.; Cao, X.C.; Yu, S.M.; Bohr, J.A.; Hu, J.J.; Jin, Q.Y. Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 5, 259–263. [Google Scholar] [CrossRef]
- Thai Meteorological Department. Climate of Thailand Report. Available online: https://www.tmd.go.th/en/archive/thailand_climate.pdf (accessed on 8 November 2022).
- Soil Resources Survey and Research Division, Land Develop Department. Esarn Soil Series. Available online: http://oss101.ldd.go.th/web_thaisoilinf/northeast/Khonkaen/kk_map/kk_man62/4001/400111_home.html# (accessed on 11 November 2022).
- Norman, A.G. Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; Soil Science Society of America: Madison, WI, USA, 1983. [Google Scholar] [CrossRef]
- Vanavichit, A.; Kamolsukyeunyong, W.; Siangliw, M.; Siangliw, J.L.; Traprab, S.; Ruengphayak, S.; Chaichoompu, E.; Saensuk, C.; Phuvanartnarubal, E.; Toojinda, T.; et al. Thai Hom Mali Rice: Origin and breeding for subsistence rainfed lowland rice system. Rice 2018, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Kanjoo, V.; Punyawaew, K.; Siangliw, J.L.; Jearakongman, S.; Vanavichit, A.; Toojinda, T. Evaluation of agronomic traits in chromosome segment substitution lines of KDML105 containing drought tolerance QTL under drought stress. Rice Sci. 2012, 19, 117–124. [Google Scholar] [CrossRef]
- Pamuta, D. Development of Rice for Salt and Drought Tolerance Using Backcross Breeding and Marker Assisted Selection. Ph.D. Thesis, Khon Kaen University, Khon Kaen, Thailand, 2021. [Google Scholar]
- Wolf, B. The Fertile Triangle: The Interrelationship of Air, Water, and Nutrients in Maximizing Soil Productivity; The Haworth Press: New York, NY, USA, 1999. [Google Scholar]
- Bureau of Rice Research and Development, Rice Department of Thailand. Advice on the Use of Chemical Fertilizers in Rice Fields According to Soil Analysis Values. Available online: http://webold.ricethailand.go.th/rkb3/Eb_014.pdf (accessed on 11 November 2022).
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dabrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Single-Factor Experiments. In Statistical Procedures for Agricultural Research; Gomez, K.A., Gomez, A.A., Eds.; John Wiley & Sons, Inc.: Hoboken, NY, USA, 1984; pp. 8–13. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Field | Genotypes | Stage | Field × Genotypes | Field × Stage | Genotypes × Stage | Field × Genotypes × Stage | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| SPAD | 2 | 185.2 | <0.001 | 4 | 16.89 | <0.001 | 5 | 85.88 | <0.001 | 8 | 1.96 | 0.051 | 10 | 7.08 | <0.001 | 20 | 4.16 | <0.001 | 40 | 2.00 | <0.001 |
| Pn (µmol CO2 m−2 s−1) | 2 | 1.14 | 0.321 | 4 | 8.385 | <0.001 | 5 | 232.0 | <0.001 | 8 | 4.20 | <0.001 | 10 | 17.44 | <0.001 | 20 | 1.81 | 0.019 | 40 | 2.00 | <0.001 |
| gs (mol H2O m−2 s−1) | 2 | 21.41 | <0.001 | 4 | 13.44 | <0.001 | 5 | 160.9 | <0.001 | 8 | 3.69 | <0.001 | 10 | 73.60 | <0.001 | 20 | 9.12 | <0.001 | 40 | 1.97 | <0.001 |
| Tr (mmol H2O m−2 s−1) | 2 | 79.37 | <0.001 | 4 | 21.33 | <0.001 | 5 | 29.43 | <0.001 | 8 | 1.83 | 0.072 | 10 | 69.83 | <0.001 | 20 | 3.46 | <0.001 | 40 | 2.04 | <0.001 |
| WUE (µmol CO2 mmol H2O−1) | 2 | 96.92 | <0.001 | 4 | 15.09 | <0.001 | 5 | 115.2 | <0.001 | 8 | 6.53 | <0.001 | 10 | 75.88 | <0.001 | 20 | 9.60 | <0.001 | 40 | 2.60 | <0.001 |
| ΦPSII | 2 | 11.38 | <0.001 | 4 | 1.734 | 0.143 | 5 | 182.3 | <0.001 | 8 | 1.52 | 0.151 | 10 | 9.99 | <0.001 | 20 | 1.91 | 0.012 | 40 | 1.78 | 0.004 |
| Fv’/Fm’ | 2 | 81.39 | <0.001 | 4 | 34.5 | <0.001 | 5 | 140.6 | <0.001 | 8 | 1.45 | 0.177 | 10 | 3.29 | <0.001 | 20 | 3.62 | <0.001 | 40 | 1.24 | 0.167 |
| ETR (µmol e− m−2 s−1) | 2 | 11.39 | <0.001 | 4 | 1.717 | 0.146 | 5 | 182.2 | <0.001 | 8 | 1.53 | 0.148 | 10 | 9.99 | <0.001 | 20 | 1.91 | 0.012 | 40 | 1.78 | 0.004 |
| Parameter | Field | Genotypes | Stage | Field × Genotypes | Field × Stage | Genotypes × Stage | Field × Genotypes × Stage | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | df | F | p | |
| Plant height (cm) | 2 | 69.44 | <0.001 | 4 | 35.89 | <0.001 | 3 | 1605 | <0.001 | 8 | 2.28 | 0.024 | 6 | 8.94 | <0.001 | 12 | 6.27 | <0.001 | 24 | 1.12 | 0.325 |
| Leaf dry weight (gDW plant−1) | 2 | 37.81 | <0.001 | 4 | 9.63 | <0.001 | 3 | 231 | <0.001 | 8 | 3.77 | <0.001 | 6 | 7.43 | <0.001 | 12 | 3.18 | <0.001 | 24 | 1.93 | 0.009 |
| Stem dry weight (gDW plant−1) | 2 | 51.97 | <0.001 | 4 | 15.04 | <0.001 | 3 | 283 | <0.001 | 8 | 5.98 | <0.001 | 6 | 16.65 | <0.001 | 12 | 7.44 | <0.001 | 24 | 2.43 | <0.001 |
| Root dry weight (gDW plant−1) | 2 | 17.52 | <0.001 | 4 | 1.723 | 0.147 | 3 | 86 | <0.001 | 8 | 1.52 | 0.153 | 6 | 16.64 | <0.001 | 12 | 2.05 | 0.022 | 24 | 2.86 | <0.001 |
| Total dry weight (gDW plant−1) | 2 | 60.01 | <0.001 | 4 | 2.88 | 0.024 | 3 | 561 | <0.001 | 8 | 5.08 | <0.001 | 6 | 21.98 | <0.001 | 12 | 2.62 | 0.003 | 24 | 1.37 | 0.128 |
| Parameter | Field | Genotypes | Field × Genotypes | ||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| Na content (%) | 2 | 121.59 | <0.001 | 1 | 14.09 | <0.001 | 2 | 8.39 | <0.001 |
| K content (%) | 2 | 14.49 | <0.001 | 1 | 0.51 | 0.478 | 2 | 1.08 | 0.343 |
| K/Na ratio | 2 | 149.11 | <0.001 | 1 | 10.16 | 0.002 | 2 | 3.37 | 0.038 |
| Panicle length (cm) | 2 | 5.01 | 0.011 | 4 | 2.65 | 0.046 | 8 | 3.36 | 0.004 |
| Total grain number panicle−1 | 2 | 2.19 | 0.124 | 4 | 1.92 | 0.124 | 8 | 1.02 | 0.438 |
| No. of filled grains panicle−1 | 2 | 3.32 | 0.050 | 4 | 0.56 | 0.692 | 8 | 1.92 | 0.081 |
| No. of unfilled grains panicle−1 | 2 | 0.54 | 0.589 | 4 | 6.07 | <0.001 | 8 | 2.55 | 0.022 |
| Filled grain weight panicle−1 (g) | 2 | 5.84 | 0.006 | 4 | 0.71 | 0.588 | 8 | 2.36 | 0.033 |
| Filled grain weight plant−1 (g) | 2 | 36.29 | <0.001 | 4 | 1.81 | 0.143 | 8 | 4.30 | <0.001 |
| Weight of 100 filled grains (g) | 2 | 55.80 | <0.001 | 4 | 51.16 | <0.001 | 8 | 2.10 | 0.056 |
| Rice Genotype | Genotype | Saline Field | 2 Critical-p | ||
|---|---|---|---|---|---|
| Non-Saline | Semi-Saline | Heavy-Saline | Value | ||
| Pokkali | 0.006 ± 0.001 C | 0.014 ± 0.006 cB | 0.027 ± 0.003 A | p < 0.01 | |
| RD73 | 0.007 ± 0.001 B | 0.024 ± 0.006 bA | 0.028 ± 0.006 A | p < 0.01 | |
| Na content (%) | CSSL8-94 | 0.007 ± 0.002 B | 0.030 ± 0.001 aA | 0.031 ± 0.002 A | p < 0.01 |
| TSKC1-144 | 0.007 ± 0.001 C | 0.016 ± 0.005 cB | 0.026 ± 0.008 A | p < 0.01 | |
| KDML105 | 0.008 ± 0.003 B | 0.030 ± 0.002 aA | 0.030 ± 0.002 A | p < 0.01 | |
| 1 Critical-p value | ns | p < 0.01 | ns | ||
| mean | 0.007 C | 0.023 B | 0.028 A | p < 0.01 | |
| Pokkali | 0.852 ± 0.218 abA | 0.683 ± 0.207 AB | 0.472 ± 0.141 B | p < 0.01 | |
| RD73 | 0.732 ± 0.124 bcA | 0.546 ± 0.149 B | 0.488 ± 0.187 B | p < 0.01 | |
| K content (%) | CSSL8-94 | 0.655 ± 0.095 cA | 0.642 ± 0.100 A | 0.432 ± 0.203 B | p < 0.01 |
| TSKC1-144 | 0.952 ± 0.115 aA | 0.754 ± 0.320 B | 0.602 ± 0.124 B | p < 0.01 | |
| KDML105 | 0.650 ± 0.232 c | 0.591 ± 0.109 | 0.592 ± 0.268 | ns | |
| 1 Critical-p value | p < 0.01 | ns | ns | ||
| mean | 0.768 A | 0.643 B | 0.517 C | p < 0.01 | |
| Pokkali | 123 ± 25 aA | 52 ± 23 aB | 16 ± 4 C | p < 0.01 | |
| RD73 | 94 ± 20 bA | 22 ± 4 bB | 18 ± 5 B | p < 0.01 | |
| K/Na ratio | CSSL8-94 | 91 ± 25 bA | 21 ± 4 bB | 13 ± 6 B | p < 0.01 |
| TSKC1-144 | 138 ± 24 aA | 45 ± 12 aB | 24 ± 6 C | p < 0.01 | |
| KDML105 | 81 ± 27 bA | 19 ± 3 bB | 19 ± 10 B | p < 0.01 | |
| 1 Critical-p value | p < 0.01 | p < 0.01 | ns | ||
| mean | 106 A | 32 B | 18 C | p < 0.01 | |
| Trait | Genotype | Saline Field | 1 Critical-p | ||
|---|---|---|---|---|---|
| Non-Saline | Semi-Saline | Heavy-Saline | Value | ||
| Pokkali | 28.9 ± 1.6 bc | 27.5 ± 0.9 (−5%) | 28.5 ± 1.1 (−1%) | ns | |
| RD73 | 30.7 ± 1.3 abA | 28.8 ± 1.1 AB (−6%) | 24.1 ± 1.7 B (−21%) | p < 0.01 | |
| Panicle length | CSSL8-94 | 25.4 ± 2.1 c | 29.9 ± 0.7 (+18%) | 27.9 ± 0.3 (+10%) | ns |
| (cm) | TSKC1-144 | 33.7 ± 0.8 aA | 29.6 ± 1.3 AB (−12%) | 26.9 ± 1.4 B (−20%) | p < 0.01 |
| KDML105 | 26.9 ± 0.7 bc | 27.3 ± 0.4 (+1%) | 26.3 ± 1.3 (−2%) | ns | |
| 2 Critical-p value | p < 0.05 | ns | ns | ||
| mean | 29.1 ± 1.6 A | 28.6 ± 0.6 A | 26.7 ± 0.8 B | p< 0.05 | |
| Pokkali | 162 ± 20 | 170 ± 17 (+5%) | 165 ± 10 (+2%) | ns | |
| RD73 | 181 ± 21 | 184 ± 16 (+2%) | 151 ± 20 (−17%) | ns | |
| Total grain | CSSL8-94 | 183 ± 22 | 213 ± 15 (+14%) | 166 ± 5 (−1%) | ns |
| number/panicle | TSKC1-144 | 233 ± 27 | 181 ± 24 (−23%) | 181 ± 27 (−36%) | ns |
| KDML105 | 152 ± 4 | 185 ± 22 (+22%) | 154 ± 11 (−6%) | ns | |
| 2 Critical-p value | ns | ns | ns | ||
| mean | 182 ± 15 | 186 ± 7 | 163 ± 7 | ns | |
| Pokkali | 106 ± 12 c | 157 ± 17 (+48%) | 143 ± 9 (+35%) | ns | |
| RD73 | 163 ± 19 ab | 172 ± 17 (+6%) | 101 ± 34 (−38%) | ns | |
| No. of filled | CSSL8-94 | 146 ± 12 ab | 167 ± 6 (+14%) | 145 ± 1 (−1%) | ns |
| grains/panicle | TSKC1-144 | 175 ± 14 a | 119 ± 25 (−32%) | 112 ± 38 (−36%) | ns |
| KDML105 | 127 ± 2 bc | 155 ± 17 (+22%) | 119 ± 16 (−6%) | ns | |
| 2 Critical-p value | p < 0.05 | ns | ns | ||
| mean | 143 ± 13 | 154 ± 10 | 124 ± 9 | ns | |
| Pokkali | 55 ± 15 A | 13 ± 1 bcB (−76%) | 15 ± 4 cB (−73%) | p < 0.01 | |
| RD73 | 18 ± 3 B | 12 ± 2 cB (−33%) | 50 ± 15 abA (+178%) | p < 0.01 | |
| No. of unfilled | CSSL8-94 | 37 ± 12 | 46 ± 11 ab (+24%) | 20 ± 4 c (−46%) | ns |
| grains/panicle | TSKC1-144 | 58 ± 14 | 61 ± 20 a (+5%) | 69 ± 13 a (+19%) | ns |
| KDML105 | 24 ± 5 | 30 ± 11 a–c (+25%) | 35 ± 5 bc (+46%) | ns | |
| 2 Critical-p value | ns | p < 0.05 | p < 0.05 | ||
| mean | 38 ± 9 | 32 ± 10 | 38 ± 11 | ns | |
| Pokkali | 3.42 ± 0.38 c | 4.77 ± 0.54 (+39%) | 4.42 ± 0.31 (+29%) | ns | |
| RD73 | 4.70 ± 0.53 abA | 4.38 ± 0.52 A (−7%) | 2.22 ± 0.79 B (−53%) | p < 0.01 | |
| Filled grain | CSSL8-94 | 3.98 ± 0.29 bc | 4.28 ± 0.11 (+8%) | 3.58 ± 0.03 (−10%) | ns |
| weight/panicle | TSKC1-144 | 5.08 ± 0.21 a | 3.38 ± 0.75 (−33%) | 3.03 ± 1.08 (−40%) | ns |
| (g) | KDML105 | 3.76 ± 0.07 bc | 4.00 ± 0.38 (+6%) | 2.80 ± 0.50 (−26%) | ns |
| 2 Critical-p value | p < 0.05 | ns | ns | ||
| mean | 4.19 ± 0.34 A | 4.16 ± 0.26 A | 3.21 ± 0.42 B | p < 0.05 | |
| Pokkali | 14.98 ± 1.78 c | 24.62 ± 1.59 (+64%) | 21.27 ± 3.33 a (+42%) | ns | |
| RD73 | 25.27 ± 1.19 aB | 31.26 ± 1.94 A (+24%) | 13.95 ± 3.42 bC (−45%) | p < 0.01 | |
| Filled grain | CSSL8-94 | 21.73 ± 1.08 abB | 24.58 ± 1.33 A (+13%) | 14.07 ± 1.79 bB (−35%) | p < 0.01 |
| weight/plant | TSKC1-144 | 21.10 ± 1.31 b | 22.41 ± 3.20 (+6%) | 14.41 ± 1.29 b (−32%) | ns |
| (g) | KDML105 | 25.36 ± 1.02 aA | 26.46 ± 2.80 A (+4%) | 10.77 ± 1.04 bB (−58%) | p < 0.01 |
| 2 Critical-p value | p < 0.01 | ns | p < 0.05 | ||
| mean | 21.68 ± 2.12 B | 25.86 ± 1.67 A | 14.89 ± 1.93 C | p< 0.01 | |
| Pokkali | 3.39 ± 0.06 aA | 3.18 ± 0.02 aB (−6%) | 3.17 ± 0.04 aB (−6%) | p < 0.05 | |
| RD73 | 3.02 ± 0.04 bA | 2.58 ± 0.07 bB (−15%) | 2.40 ± 0.07 cB (−21%) | p < 0.05 | |
| Weight | CSSL8-94 | 2.70 ± 0.06 c | 2.59 ± 0.06 b (−4%) | 2.41 ± 0.09 bc (−11%) | ns |
| of 100 filledgrains | TSKC1-144 | 3.09 ± 0.09 b | 2.71 ± 0.11 b (−12%) | 2.62 ± 0.08 b (−15%) | ns |
| (g) | KDML105 | 2.93 ± 0.03 b | 2.51 ± 0.08 b (−14%) | 2.38 ± 0.01 c (−19%) | p < 0.05 |
| 2 Critical-p value | p < 0.05 | p < 0.05 | p < 0.05 | ||
| mean | 3.03 ± 0.12 A | 2.71 ± 0.13 B | 2.59 ± 0.17 C | p< 0.05 | |
| Soil Physicochemical Properties | Non-Saline | Semi-Salinity | Heavy-Salinity |
|---|---|---|---|
| Physical properties | |||
| Soil Texture | Sandy Loam | Loam | Loam |
| Sand (%) | 71.82 | 40.75 | 37.33 |
| Silt (%) | 23.44 | 33.67 | 37.56 |
| Clay (%) | 4.74 | 25.58 | 25.11 |
| Chemical properties at pre-planting | |||
| Total N (%) | 0.031 ± 0.001 | 0.083 ± 0.001 | 0.041 ± 0.001 |
| Available P (mg kg−1) | 46.15 ± 0.81 | 15.56 ± 0.92 | 9.23 ± 0.44 |
| Exchangeable K (mg kg−1) | 31.96 ± 0.86 | 69.48 ± 1.25 | 52.68 ± 1.45 |
| pH (1:1 H2O) | 6.67 ± 0.01 | 7.176 ± 0.02 | 7.113 ± 0.01 |
| EC (dS m−1) | 0.12 ± 0.01 | 2.87 ± 0.02 | 2.53 ± 0.01 |
| SOM (%) | 0.59 ± 0.03 | 1.21 ± 0.04 | 0.44 ± 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santanoo, S.; Lontom, W.; Dongsansuk, A.; Vongcharoen, K.; Theerakulpisut, P. Photosynthesis Performance at Different Growth Stages, Growth, and Yield of Rice in Saline Fields. Plants 2023, 12, 1903. https://doi.org/10.3390/plants12091903
Santanoo S, Lontom W, Dongsansuk A, Vongcharoen K, Theerakulpisut P. Photosynthesis Performance at Different Growth Stages, Growth, and Yield of Rice in Saline Fields. Plants. 2023; 12(9):1903. https://doi.org/10.3390/plants12091903
Chicago/Turabian StyleSantanoo, Supranee, Watanachai Lontom, Anoma Dongsansuk, Kochaphan Vongcharoen, and Piyada Theerakulpisut. 2023. "Photosynthesis Performance at Different Growth Stages, Growth, and Yield of Rice in Saline Fields" Plants 12, no. 9: 1903. https://doi.org/10.3390/plants12091903