

Comparative Analysis of Bioactive Compounds in Two Globe Artichoke Ecotypes Sanitized and Non-Sanitized from Viral Infections

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Morphological and Qualitative Traits
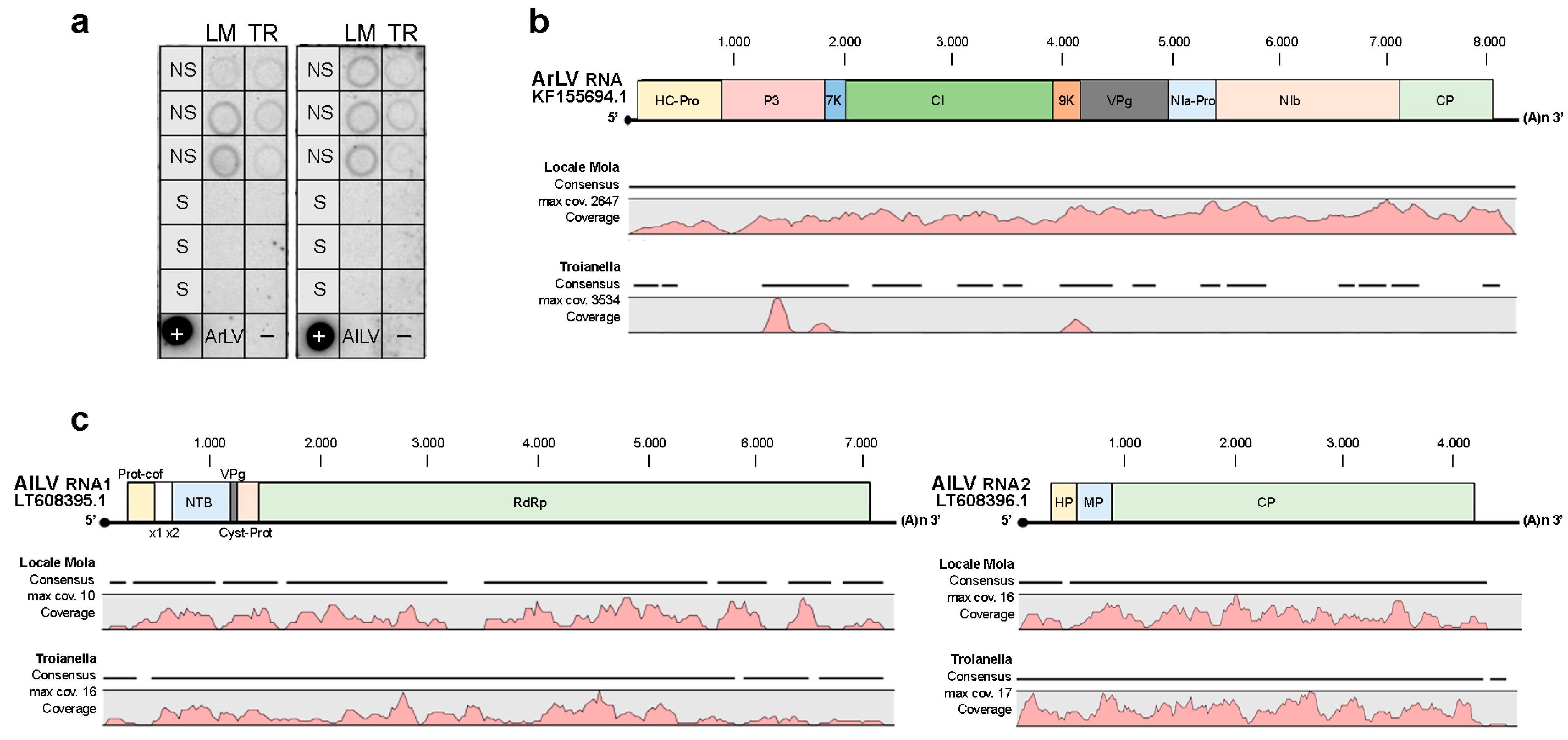
2.2. Detection of Virus Infections
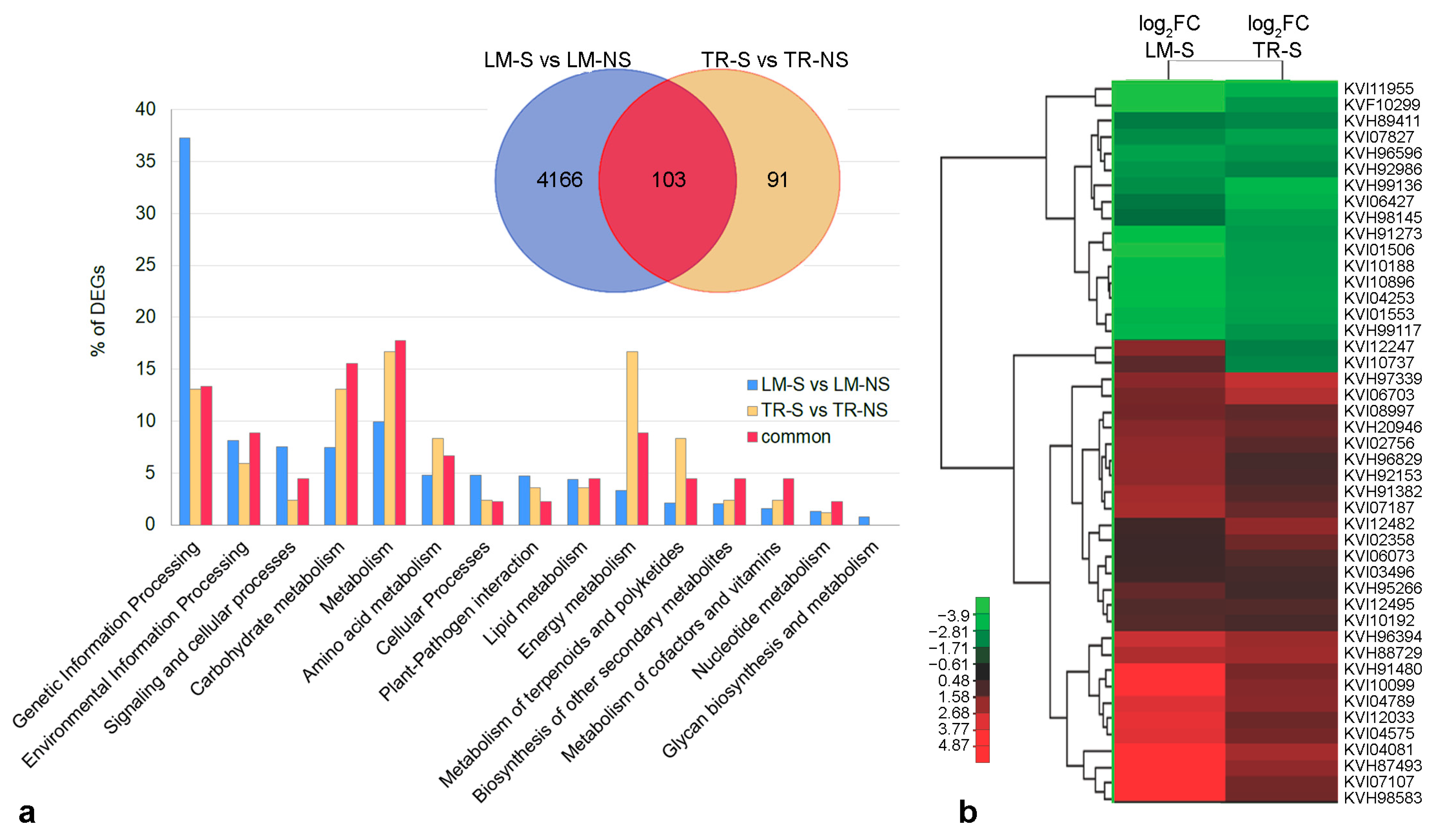
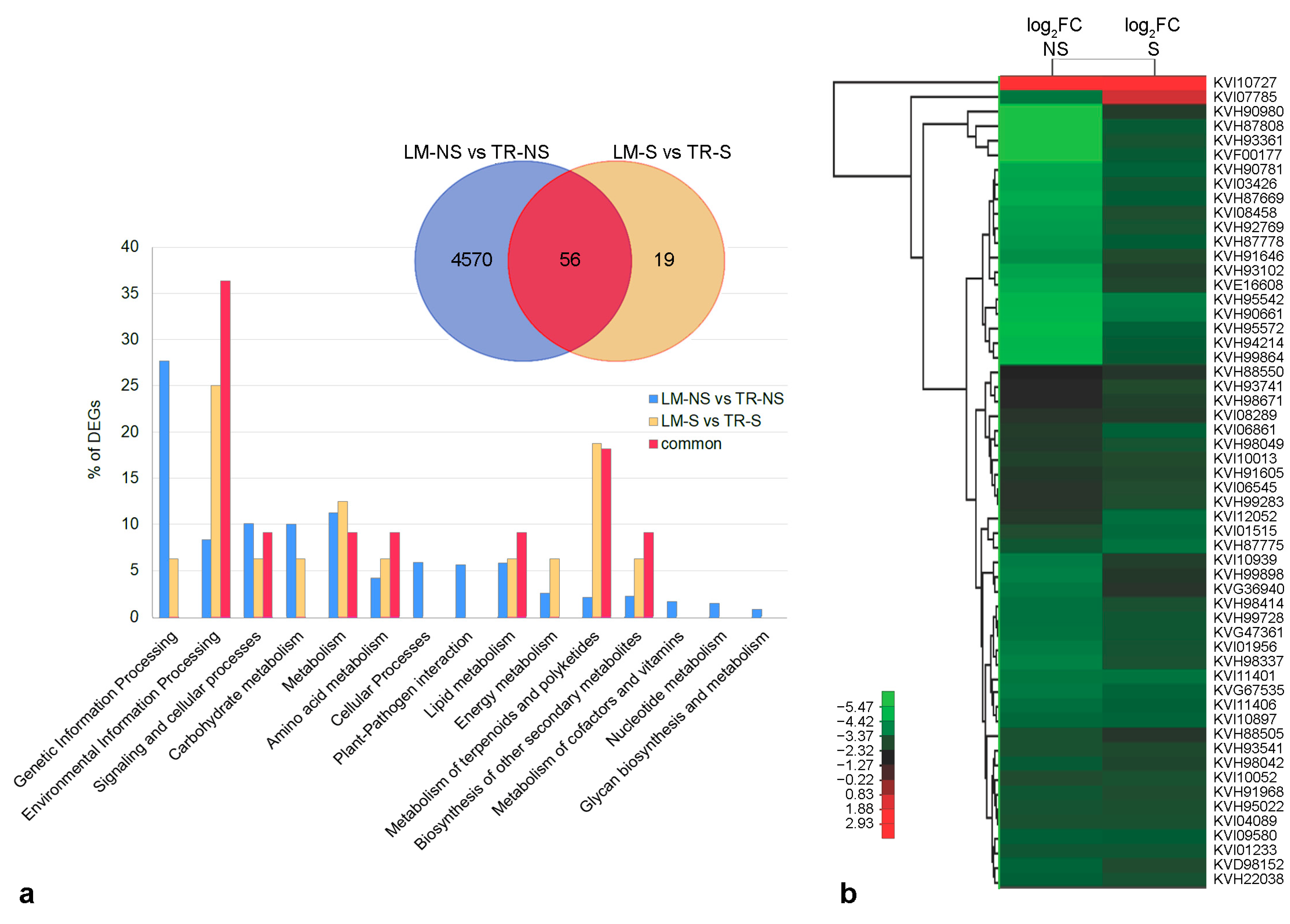
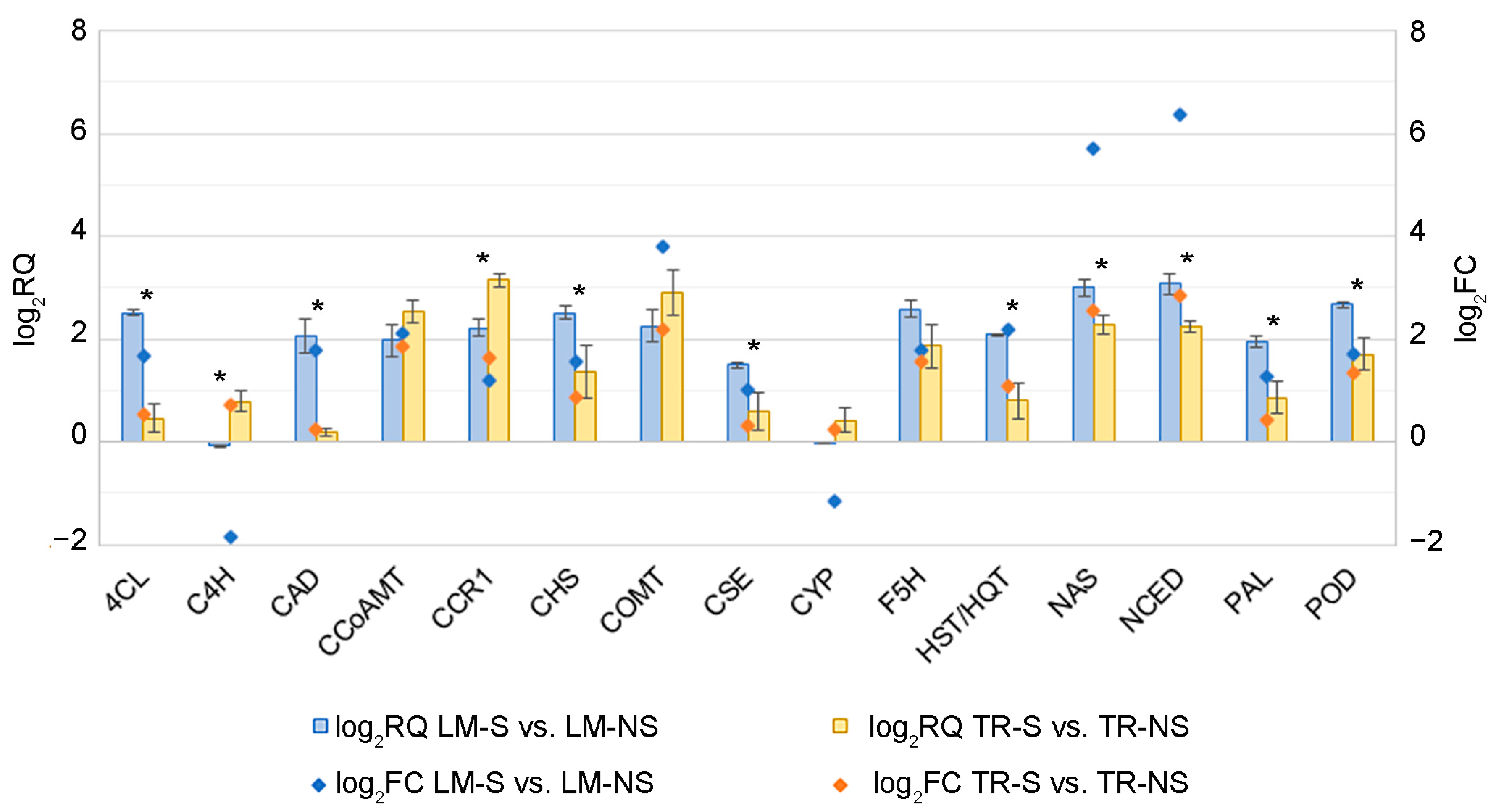
2.3. Comparative Analysis of Whole-Transcriptome of Artichoke Ecotypes
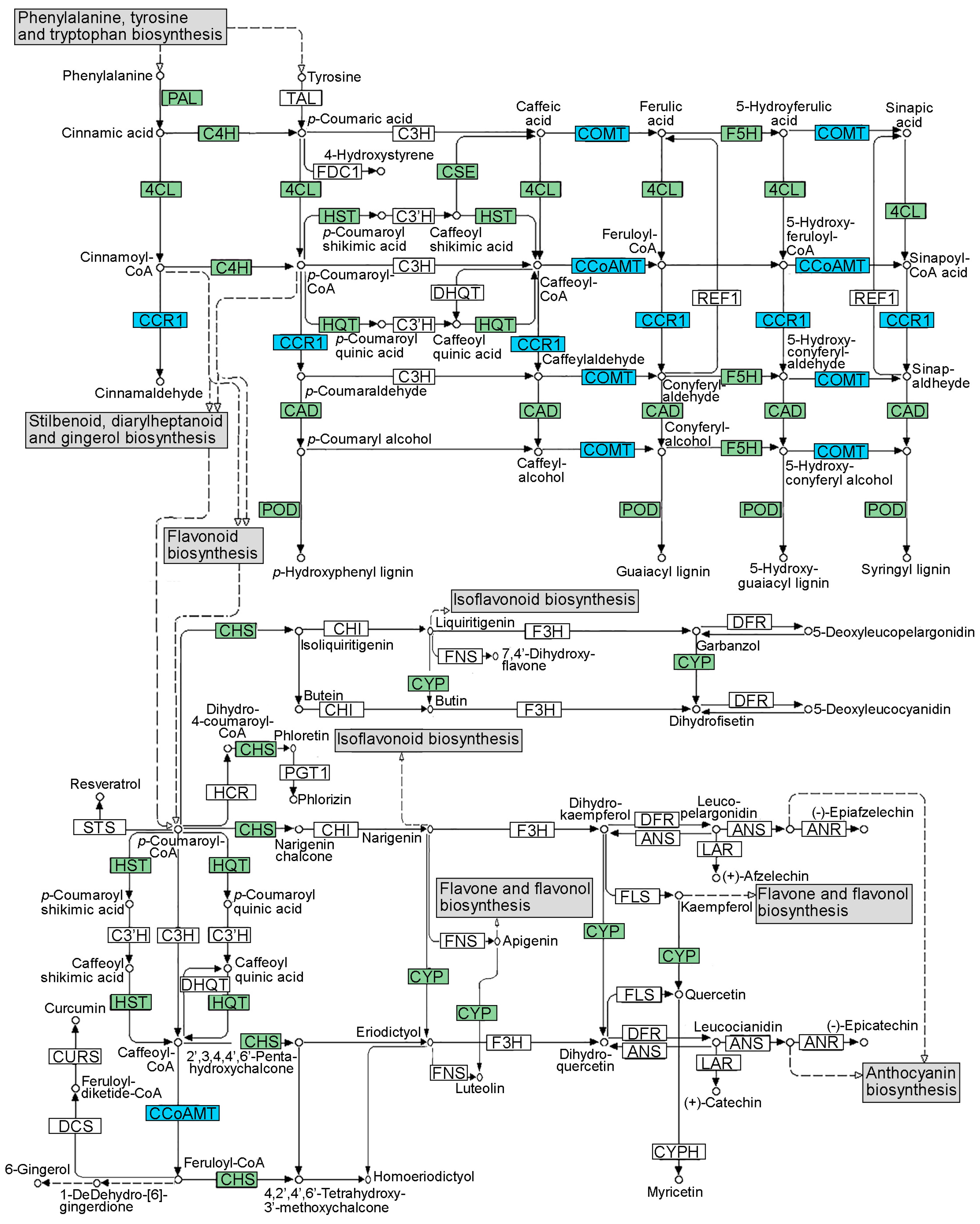
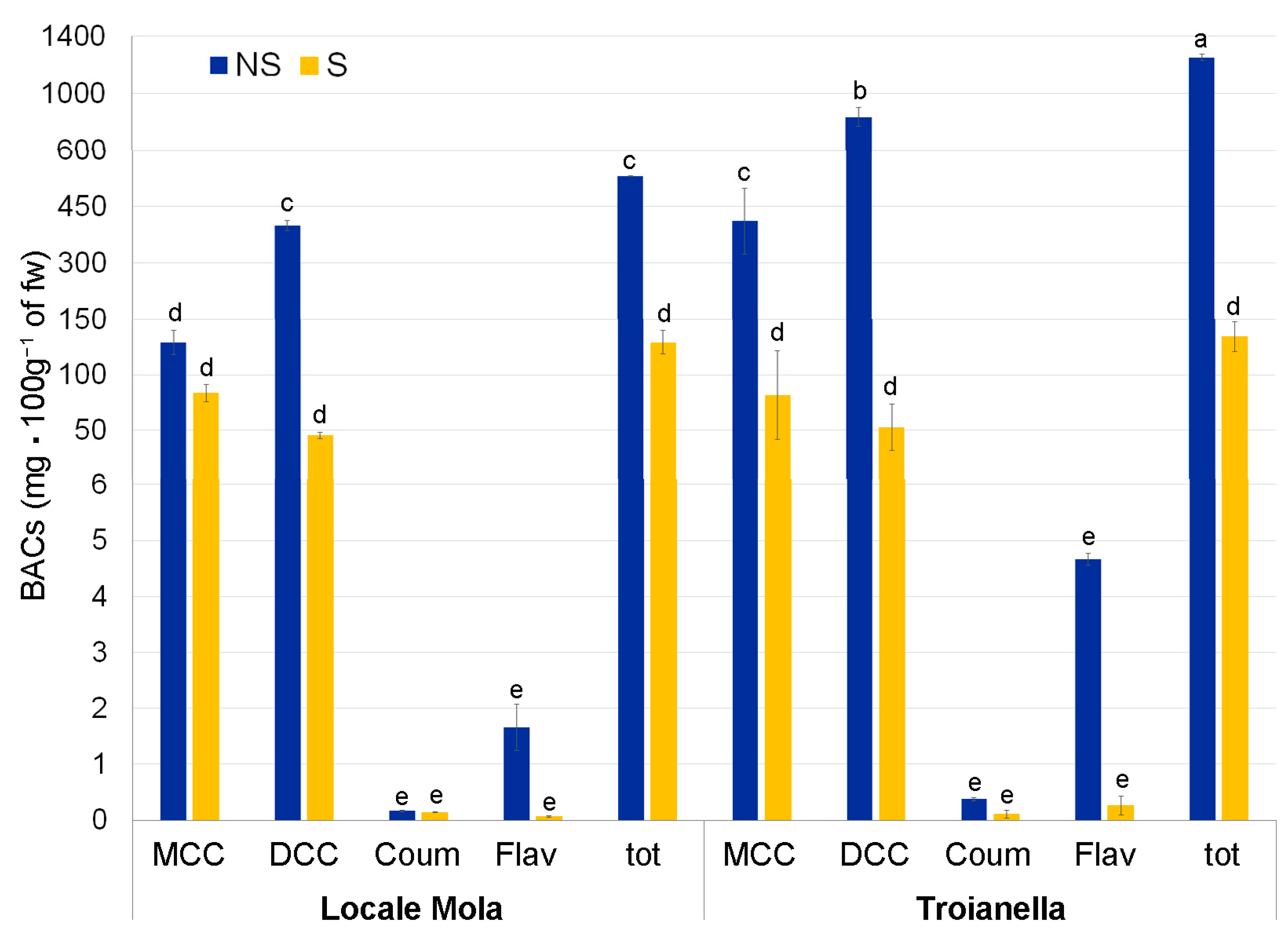
2.4. BAC Chemical Analysis and Characterization
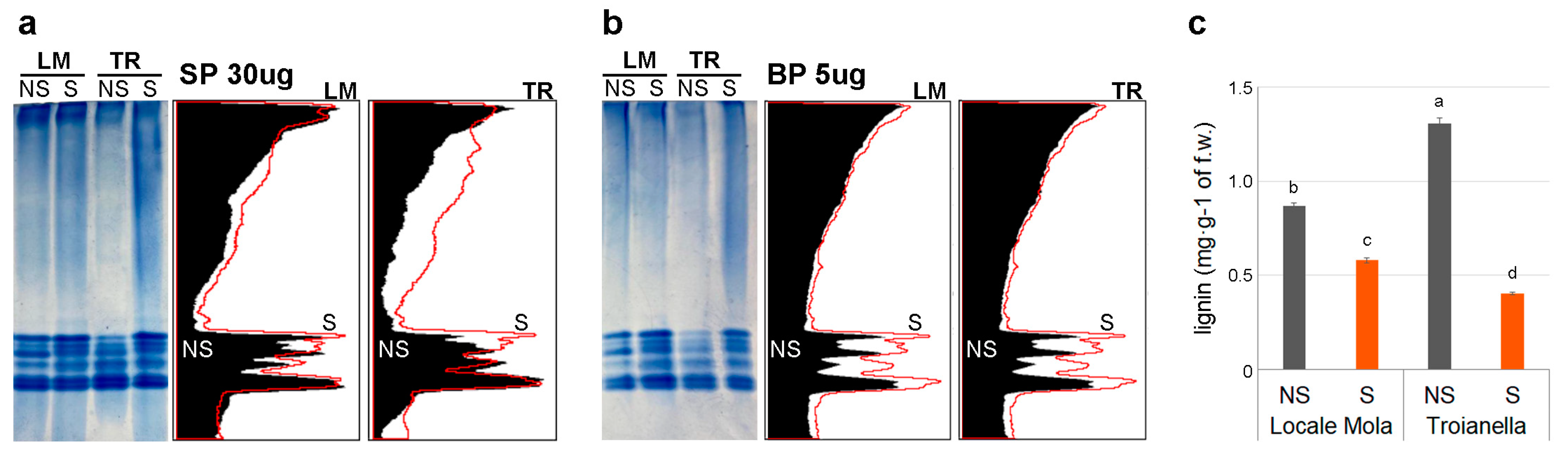
2.5. Peroxidase Activity and Lignin Content
3. Discussion
3.1. Ecotypes’ Adaptation to Environments and Phytosanitary Status
3.2. Modulation of Artichoke Ecotype Transcriptome in Sanitized Plants
3.3. Virus-Free Status Decreases the Accumulation of Polyphenols
3.4. Correlation of High POD Activity with the Accumulation of Developmental Lignin in Sanitized Plants
4. Materials and Methods
4.1. Plant Materials and Assessment of the Sanitary Status
4.2. Total RNA Extraction, cDNA Preparation, and High-Throughput mRNA Sequencing
4.3. Mapping of Sequence Reads
4.4. Validation of the RNA-Sequencing Results by Quantitative Real-Time PCR
4.5. Artichoke Polyphenol Extraction and HPLC Analysis
4.6. Determination of Artichoke Peroxidase Activity and Lignin Content
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lattanzio, V.; Kroon, P.A.; Linsalata, V.; Cardinali, A. Globe artichoke: A functional food and source of nutraceutical ingredients. J. Funct. Foods 2009, 1, 131–144. [Google Scholar] [CrossRef]
- Gaafar, A.A.; Salama, Z.A. Phenolic Compounds from Artichoke (Cynara scolymus L.) By-products and their Antimicrobial Activities. J. Biol. Agric. Healthc. 2013, 3, 1–6. [Google Scholar]
- Noriega-Rodríguez, D.; Soto-Maldonado, C.; Torres-Alarcón, C.; Pastrana-Castro, L.; Weinstein-Oppenheimer, C.; Zúñiga-Hansen, M.E. Valorization of Globe Artichoke (Cynara scolymus) Agro-Industrial Discards, Obtaining an Extract with a Selective Effect on Viability of Cancer Cell Lines. Processes 2020, 8, 715. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; Abbas, H.; Zewail, M.; Noureldin, M.H.; Ali, M.M.; Shamaa, M.M.; Khattab, M.A.; Ibrahim, N. Neuroprotective Effect of Artichoke-Based Nanoformulation in Sporadic Alzheimer’s Disease Mouse Model: Focus on Antioxidant, Anti-Inflammatory, and Amyloidogenic Pathways. Pharmaceuticals 2022, 15, 1202. [Google Scholar] [CrossRef]
- Romani, A.; Pinelli, P.; Cantini, C.; Cimato, A.; Heimler, D. Characterization of Violetto di Toscana, a typical Italian variety of artichoke (Cynara scolymus L.). Food Chem. 2006, 95, 221–225. [Google Scholar] [CrossRef]
- Mauro, R.P.; Portis, E.; Acquadro, A.; Lombardo, S.; Mauromicale, G.; Lanteri, S. Genetic diversity of globe artichoke landraces from Sicilian small-holdings: Implications for evolution and domestication of the species. Conserv. Gene 2009, 10, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Mauro, R.P.; Portis, E.; Lanteri, S.; Mauromicale, G. Genotypic and bio-agronomical characterization of an early Sicilian landrace of globe artichoke. Euphytica 2012, 186, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Ciancolini, A.; Rey, N.A.; Pagnotta, M.A.; Crinò, P. Characterization of Italian spring globe artichoke germplasm: Morphological and molecular profiles. Euphytica 2012, 186, 433–443. [Google Scholar] [CrossRef]
- Tavazza, R.; Lucioli, A.; Benelli, C.; Giorgi, D.; D’Aloisio, E.; Papacchioli, V. Cryopreservation in artichoke: Towards a phytosanitary qualified germplasm collection. Ann. Appl. Biol. 2013, 163, 231–241. [Google Scholar] [CrossRef]
- Spanò, R.; Bottalico, G.; Corrado, A.; Campanale, A.; Di Franco, A.; Mascia, T. A Protocol for Producing Virus-Free Artichoke Genetic Resources for Conservation, Breeding, and Production. Agriculture 2018, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Gallitelli, D.; Mascia, T.; Martelli, G.P. Viruses in artichoke. Adv. Virus Res. 2012, 84, 289–324. [Google Scholar]
- Rosales Calderon, O.; Arantes, V. A review on commercial-scale high-value products that can be produced alongside cellulosic ethanol. Biotechnol. Biofuels 2019, 12, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, P.; Yang, G.; Hu, L.; Sun, J.; Shi, L.; Zhou, Y.; Wang, Z.; Yang, J. Recent advances in the valorization of plant biomass. Biotechnol. Biofuels 2021, 14, 102. [Google Scholar] [CrossRef]
- Yadav, M.; Goel, G.; Hatton, F.L.; Bhagat, M.; Mehta, S.K.; Mishra, R.K.; Bhojak, N. A review on biomass-derived materials and their applications as corrosion inhibitors, catalysts, food and drug delivery agents. Curr. Opin. Green Sustain. Chem. 2021, 4, 100153. [Google Scholar] [CrossRef]
- Cardinali, A.; Tursi, N.; Ligorio, A.; Giuffrida, M.G.; Napolitano, L.; Caliandro, R.; Sergio, L.; Di Venere, D.; Lattanzio, V.; Sonnante, G. Purification, biochemical characterization and cloning of a new cationic peroxidase isoenzyme from artichoke. Plant Physiol. Biochem. 2011, 49, 395–403. [Google Scholar] [CrossRef]
- Negro, D.; Montesano, V.; Grieco, S.; Crupi, P.; Sarli, G.; De Lisi, A.; Sonnante, G. Polyphenol Compounds in Artichoke Plant Tissues and Varieties. J. Food Sci. 2012, 77, C244–C252. [Google Scholar] [CrossRef]
- Desjardins, Y.; Dubuc, J.F.; Badr, A. In Vitro Culture of Plants: A Stressful Activity! ISHS Acta Horticulturae: Leuven, Belgium, 2009; Volume 812, pp. 29–50. [Google Scholar]
- Magyar-Tábori, K.; Mendler-Drienyovszki, N.; Hanász, A.; Zsombik, L.; Dobránszki, J. Phytotoxicity and Other Adverse Effects on the In Vitro Shoot Cultures Caused by Virus Elimination Treatments: Reasons and Solutions. Plants 2021, 10, 670. [Google Scholar] [CrossRef] [PubMed]
- Faraone, I.; Russo, D.; Ponticelli, M.; Candido, V.; Castronuovo, D.; Cardone, L.; Sinisgalli, C.; Labanca, F.; Milella, L. Preserving Biodiversity as Source of Health Promoting Compounds: Phenolic Profile and Biological Activity of Four Varieties of Solanum lycopersicum L. Plants 2021, 10, 447. [Google Scholar] [CrossRef]
- Martínez-Ispizua, E.; Calatayud, Á.; Marsal, J.I.; Mateos-Fernández, R.; Díez, M.J.; Soler, S.; Valcárcel, J.V.; Martínez-Cuenca, M.R. Phenotyping Local Eggplant Varieties: Commitment to Biodiversity and Nutritional Quality Preservation. Front. Plant Sci. 2021, 12, 696272. [Google Scholar] [CrossRef] [PubMed]
- Cirilli, M.; Caruso, G.; Gennai, C.; Urbani, S.; Frioni, E.; Ruzzi, M.; Servili, M.; Gucci, R.; Poerio, E.; Muleo, R.M. The Role of Polyphenoloxidase, Peroxidase, and Glucosidase in Phenolics Accumulation in Olea europaea L. Fruits under Different Water Regimes. Front. Plant Sci. 2017, 8, 717. [Google Scholar] [CrossRef] [Green Version]
- Cefola, M.; D’Antuono, I.; Pace, B.; Calabrese, N.; Carito, A.; Linsalata, V.; Cardinali, A. Biochemical relationships and browning index for assessing the storage suitability of artichoke genotypes. Food Res. Int. 2012, 48, 397–403. [Google Scholar] [CrossRef]
- Prabha, T.N.; Patwardhan, M.V. Polyphenol oxidase peroxidase enzyme activities and their isoenzyme pattern in ripening fruits. Acta Aliment. 1986, 15, 199–207. [Google Scholar]
- Sato, Y.; Sugiyama, M.; Gorecki, R.J.; Fukuda, H.; Komamine, A. Interrelationship between lignin deposition and the activities of peroxidase isoenzymes in differentiating tracheary elements of Zinnia. Planta 1993, 189, 584–589. [Google Scholar] [CrossRef]
- Pedreno, M.A.; Ferrer, M.A.; Gaspar, T.H.; Munoz, R.; Ros Barcelo, A. The polyfunctionality of cell wall peroxidases avoids the necessity of an independent H2O2-generating system for phenolic coupling in the cell walls. Plant Peroxidases Newsl. 1995, 5, 3–8. [Google Scholar]
- Dixon, R.A.; Palva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 2004, 7, 1085–1097. [Google Scholar] [CrossRef]
- Técsi, L.I.; Smith, A.M.; Maule, A.J.; Leegood, R.C. A spatial analysis of physiological changes associated with infection of cotyledons of marrow plants with cucumber mosaic virus. Plant Physiol. 1996, 111, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.F.; Guy, P.L.; Burritt, D.J.; Jameson, P.E. Changes in the activities of antioxidant enzymes in response to virus infection and hormone treatment. Physiol. Plant 2002, 114, 157–164. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A.; Mahmoud, S.Y.; Hamad, A.; Lu, G. Salicylic acid alleviates growth inhibition and oxidative stress caused by zucchini yellow mosaic virus infection in Cucurbita pepo leaves. Physiol. Mol. Plant Pathol. 2006, 69, 172–181. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A.; Mahmoud, S.Y.; Hamad, A.; Lu, G. Physiological and metabolic changes of Cucurbita pepo leaves in response to zucchini yellow mosaic virus (ZYMV) infection and salicylic acid treatments. Plant Physiol. Biochem. 2007, 45, 480–489. [Google Scholar] [CrossRef]
- Montesano, V.; Negro, D.; Sonnante, G.; Laghetti, G.; Urbano, M. Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application. Plants 2022, 11, 2067. [Google Scholar] [CrossRef]
- Bianco, V.V. Carciofo (Cynara scolymus L.). In Orticoltura; Bianco, V.V., Pimpini, F., Eds.; Patron: Bologna, Italy, 1990; pp. 209–251. [Google Scholar]
- Lanteri, S.; Portis, E. Globe Artichoke and Cardoon. In Handbook of Plant Breeding: Vegetables I; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; Volume 1, pp. 49–74. [Google Scholar]
- McKey, D.; Elias, M.; Pujol, B.; Duputié, A. The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 2010, 186, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Wagenbreth, D. Evaluation of artichoke cultivars for growing and pharmaceutical use. Beitr. Zucht. 1996, 2, 400–403. [Google Scholar]
- Sevcikova, P.; Glatz, Z.; Slanina, J. Analysis of artichoke (Cynara cardunculus L.) extract by means of micellar electrokinetic capillary chromatography. Electrophoresis 2002, 23, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.F.; Simon, J.E.; Aviles, I.F.; He, K.; Zheng, Q.Y.; Tadmor, Y. Analysis of antioxidative phenolic compounds in artichoke (Cynara scolymus L.). J. Agric. Food Chem. 2003, 51, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.; Perez, F.; Fernandez, J.; Lezaun, J.A.; Rodriguez, D.; Perea, F.; Romero, C.; Ochoa, M.J.; Garcia, M. Study of Cynara cardunculus L: Lignocellulosic biomass production in dry conditions. Acta Hort. 2004, 221, 7. [Google Scholar]
- Quilho, T.; Gominho, J.; Pereira, H. Anatomical characterisation and variability of the thistle Cynara cardunculus in view of pulping potential. IAWA J. 2004, 25, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Sonnante, G.; De Paolis, A.; Pignone, D. Relationships among artichoke cultivars and somerelated wild taxa based on AFLP markers. Plant Genet. Resour. Characterisation Util. 2003, 1, 125–133. [Google Scholar] [CrossRef]
- Minutillo, S.A.; Marais, A.; Mascia, T.; Faure, C.; Svanella-Dumas, L.; Theil, S.; Payet, A.; Perennec, S.; Schoen, L.; Gallitelli, D.; et al. Complete Nucleotide Sequence of Artichoke latent virus Shows it to be a Member of the Genus Macluravirus in the Family Potyviridae. Phytopathology 2015, 105, 1155–1160. [Google Scholar] [CrossRef] [Green Version]
- Sanfaçon, H.; Iwanami, T.; Karasev, A.V.; Van der Vlugt, R.; Wellink, J.; Wetzel, T.; Yoshikawa, N. Family Secoviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses; King, A.M.Q., Adams, M.J., Eric, B., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2011; pp. 881–899. [Google Scholar]
- Gallitelli, D.; Rana, G.L.; Vovlas, C.; Martelli, G.P. Viruses of Globe Artichoke: An Overview. J. Plant Pathol. 2004, 86, 267–281. [Google Scholar]
- Testa, M.; Cadinu, M.; Pilia, R.; Pintire, R.; Baghia, L. Virus Elimination in Artichoke: Comparison between Different Methods of Extraction and Detection Methods; ISHS Acta Horticulturae: Leuven, Belgium, 2013; Volume 283, pp. 295–299. [Google Scholar]
- Kyriakopoulou, P.E. Artichoke Italian latent virus causes artichoke patchy chlorotic stunting disease. Ann. Appl. Biol. 1995, 127, 489–497. [Google Scholar] [CrossRef]
- Salleh, W.; Minutillo, S.A.; Spanò, R.; Zammouri, S.; Gallitelli, D.; Mnari-Hattab, M. Occurrence of artichoke infecting viruses in Tunisia. EPPO Bull. 2017, 47, 48–56. [Google Scholar] [CrossRef]
- Barba, M.; Di Lernia, G.; Babes, G.; Citrulli, F. Produzione e conservazione di germoplasma di carciofo di tipo ‘Romanesco’ esente da virus. Italus Hortus 2004, 11, 5–10. [Google Scholar]
- Papanice, M.A.; Campanale, A.; Bottalico, G.; Sumerano, P.; Gallitelli, G. Produzione di germoplama risanato di carciofo brindisino. Italus Hortus 2004, 11, 11–15. [Google Scholar]
- Saccardo, F.; Micozzi, F.; Di Lernia, G.; Piccioni, C.; Barba, M.; Pagnotta, M.A. Virus free artichoke germplasm, quali-quantitative response of globe artichoke. Acta Hortic. 2007, 730, 375–379. [Google Scholar] [CrossRef]
- Olsen, K.M.; Lea, U.S.; Slimestad, R.; Verheul, M.; Lillo, C. Differential expression of the four Arabidopsis PAL genes –PAL1 and PAL2 have functional specialization in abiotic environmental triggered flavonoid synthesis. J. Plant Physiol. 2008, 165, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Vashisth, D.; Misra, A.; Akhtar, M.Q.; Jalil, S.U.; Shanker, K.; Gupta, M.M.; Rout, P.K.; Gupta, A.K.; Shasany, A.K. RNAi down-regulation of cinnamate-4-hydroxylase increases artemisinin biosynthesis in Artemisia annua. Sci. Rep. 2016, 6, 26458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilmiller, A.L.; Stout, J.; Weng, J.K.; Humphreys, J.; Ruegger, M.O.; Chapple, C. Mutations in the cinnamate 4-hydroxylase gene impact metabolism, growth and development in Arabidopsis. Plant J. 2009, 60, 771–782. [Google Scholar] [CrossRef]
- Di Donato, R.J.; Roberts, L.A.; Sanderson, T.; Eisley, R.B.; Walker, E.L. Arabidopsis Yellow Stripe-Like2 (YSL2), a metal-regulated gene encoding a plasma membrane transporter of nicotianamine-metal complexes. Plant J. 2004, 39, 403–414. [Google Scholar] [CrossRef]
- Zhu, X.F.; Dong, X.Y.; Wu, Q.; Shen, R. Ammonium regulates Fe deficiency responses by enhancing nitric oxide signaling in Arabidopsis thaliana. Planta 2019, 250, 1089–1102. [Google Scholar] [CrossRef]
- Bienfait, H.F.; Vandenbriel, W.; Meslandmul, N.T. Free space iron pools in roots-generation and mobilization. Plant Physiol. 1985, 78, 596–600. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.F.; Wang, B.; Song, W.F.; Zheng, S.J.; Shen, R.F. Putrescine alleviates iron deficiency via NO-dependent reutilization of root cell-wall Fe in Arabidopsis. Plant Physiol. 2016, 170, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Cicco, N.; Linsalata, V. Antioxidant activities of artichoke phenolics. Acta Hortic. 2005, 681, 421–442. [Google Scholar] [CrossRef]
- Stiller, A.; Garrison, K.; Gurdyumov, K.; Kenner, J.; Yasmin, F.; Yates, P.; Song, B.H. From Fighting Critters to Saving Lives, Polyphenols in Plant Defense and Human Health. Int. J. Mol. Sci. 2021, 22, 8995. [Google Scholar] [CrossRef] [PubMed]
- Herms, D.A.; Mattson, W.J. The dilemma of plants, To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]
- Caretto, S.; Linsalata, V.; Colella, G.; Mita, G.; Lattanzio, V. Carbon Fluxes between Primary Metabolism and Phenolic Pathway in Plant Tissues under Stress. Int. J. Mol. Sci. 2015, 16, 26378–26394. [Google Scholar] [CrossRef] [Green Version]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Leyva, A.; Jarillo, J.A.; Salinas, J.; Martinez-Zapater, J.M. Low temperature induces the accumulation of phenylalanine ammonia-lyase and chalcone synthase mRNAs of Arabidopsis thaliana in a light-dependent manner. Plant Physiol. 1995, 108, 39–46. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance, A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Lillo, C.; Lea, U.S.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ. 2008, 31, 587–601. [Google Scholar] [CrossRef]
- Adams-Phillip, L.; Briggs, A.G.; Bent, A.F. Disruption of poly(ADP-ribosyl)ation mechanisms alters responses of Arabidopsis to biotic stress. Plant Physiol. 2010, 152, 267–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, S.; Boudet, A. Defence lignin and hydroxycinnamyl alcohol dehydrogenase activities in wounded Eucalyptus gunnii. For. Pathol. 2003, 33, 339–352. [Google Scholar] [CrossRef]
- Lauvergeat, V.; Lacomme, C.; Lacombe, E.; Lasserre, E.; Roby, D.; Grima-Pettenati, J. Two cinnamoyl-CoA reductase (CCR) genes from Arabidopsis thaliana are differentially expressed during development and in response to infection with pathogenic bacteria. Phytochemistry 2001, 57, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Di Venere, D.; Linsalata, V.; Calabrese, N.; Pieralice, M.; Bianco, V.V. Morphological and biochemical changes during growth and development of artichoke buds. Acta Hortic. 2005, 681, 437–443. [Google Scholar] [CrossRef]
- García-Martínez, N.; Andreo-Martínez, P.; Almela, L. Characterization of Six Artichoke Cultivars and Their Suitability for Agro-industrial Processing. J. Food Nutr. Res. 2017, 5, 234–242. [Google Scholar]
- De Klerk, G.-J. Stress in plants cultured in vitro. Propag. Ornam. Plants 2007, 7, 129–137. [Google Scholar]
- Lukaszuk, E.; Ciereszko, I. Plant responses to wounding stress. In Biological Diversity—From Cell to Ecosystem; Laska, G., Ed.; Polis Botanical Society: Bialystok, Poland, 2012; pp. 73–85. [Google Scholar]
- Barnett, O. Surveying for plant viruses: Design nd consideration. In Introduction to Plant Disease Epidemiology; Campbell, C.L., Madden, L.V., Eds.; Wiley Interscience: New York, NY, USA, 1986; pp. 147–166. [Google Scholar]
- Cravero, V.; Martin, E.; Cointry, E. Genetic diversity in Cynara cardunculus determined by Sequence-related Amplified Polymorphism Markers. J. Amer. Soc. Hortic. Sci. 2007, 132, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Minutillo, S.A.; Spanò, R.; Gallitelli, D.; Mascia, T. Simultaneous detection of 10 viruses in globe artichoke by a synthetic oligonucleotide-based DNA polyprobe. Eur. J. Plant Pathol. 2021, 160, 991–997. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR, ultrafast universal RNA-seq aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. Feature Counts, an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate, a practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 89–300. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA, KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Meth. 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV), high performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Blanco, E.; Sabetta, W.; Danzi, D.; Negro, D.; Passeri, V.; De Lisi, A.; Paolocci, F.; Sonnante, G. Isolation and Characterization of the Flavonol Regulator CcMYB12 From the Globe Artichoke [Cynara cardunculus var. scolymus (L.) Fiori]. Front. Plant Sci. 2018, 9, 941. [Google Scholar] [CrossRef] [Green Version]
- Spanò, R.; Mascia, T.; Kormelink, R.; Gallitelli, D. Grafting on a non-transgenic tolerant tomato variety confers resistance to the infection of a Sw5-breaking strain of Tomato spotted wilt virus via RNA silencing. PLoS ONE 2015, 10, e0141319. [Google Scholar] [CrossRef] [Green Version]
- Spanò, R.; Gallitelli, D.; Mascia, T. Grafting to manage infections of top stunting and necrogenic strains of Cucumber mosaic virus in tomato. Ann. Appl. Biol. 2017, 171, 393–404. [Google Scholar] [CrossRef]
- Spanò, R.; Ferrara, M.; Montemurro, C.; Mulè, G.; Gallitelli, D.; Mascia, T. Grafting alters tomato transcriptome and enhances tolerance to an airborne virus infection. Sci. Rep. 2020, 10, 2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Antuono, I.; Garbetta, A.; Linsalata, V.; Minervini, F.; Cardinali, A. Polyphenols from artichoke heads (Cynara cardunculus (L.) subsp. scolymus Hayek): In vitro bio-accessibility, intestinal uptake and bioavailability. Food Funct. 2015, 6, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Survila, M.; Davidsson, P.R.; Pennanen, V.; Kariola, T.; Broberg, M.; Sipari, N.; Heino, P.; Palva, E.T. Peroxidase-Generated Apoplastic ROS Impair Cuticle Integrity and Contribute to DAMP-Elicited Defenses. Front. Plant Sci. 2016, 7, 1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UPOV n. 1 | Feature | Artichoke Ecotypes’ Feature Expression 2 | |
|---|---|---|---|
| Locale di Mola | Troianella | ||
| Transplant | July–September | July–September | |
| 28 | Central flower head production | December–May | February–May |
| Reproductive cycle | 8–10 months | 8–10 months | |
| 1 | Plant height | 100 cm | 80 cm |
| 2 | Number of lateral heads on the main stem | 4 | 4 |
| 5 | Main stem diameter | 2 cm | 2.5 cm |
| 8 | Leaf length | 95 cm | 100 cm |
| Plant diameter | 160 cm | 190 cm * | |
| 23 | Central flower head length | 11.5 cm | 10.5 cm |
| 24 | Central flower head diameter | 8 cm | 9 cm |
| 25 | Central flower head weight | 180 g | 220 g * |
| 26 | Central flower head shape | oval | oval |
| 51 | Shoots production | 3–4 | 3–4 |
| Comparison Factors | n. of DEGs 1 | % of DEGs 2 | n. of DEGs in Common 3 |
|---|---|---|---|
| LM-S vs. LM-NS | 4269 | 16.10 | 103 |
| TR-S vs. TR-NS | 194 | 0.73 | |
| LM-NS vs. TR-NS | 4626 | 17.45 | 56 |
| LM-S vs. TR-S | 75 | 0.28 |
| Gene Name | Gene ID | Transcript ID | KO | Protein Description | EC |
|---|---|---|---|---|---|
| Phenylpropanoid biosynthesis | |||||
| CCoAMT | Ccrd_012618 | KVI08997 | K00588 | caffeoyl-CoA O-methyltransferase | 2.1.1.104 |
| CCR1 | Ccrd_015577 | KVI06073 | K09753 | cinnamoyl-CoA reductase 1 | 1.2.1.44 |
| COMT | Ccrd_017109 | KVI04575 | K13066 | caffeic acid 3-O-methyltransferase | 2.1.1.68 |
| Flavonoid biosynthesis | |||||
| CCoAMT | Ccrd_012618 | KVI08997 | K00588 | caffeoyl-CoA O-methyltransferase | 2.1.1.104 |
| Stilbenoid, diarylheptanoid, and gingerol biosynthesis | |||||
| CCoAMT | Ccrd_012618 | KVI08997 | K00588 | caffeoyl-CoA O-methyltransferase | 2.1.1.104 |
| Carotenoid biosynthesis | |||||
| NCED | Ccrd_017614 | KVI04081 | K09840 | 9-cis-epoxycarotenoid dioxygenase | 1.13.11.51 |
| Biosynthesis of various plant secondary metabolites | |||||
| NAS | Ccrd_025247 | KVH87493 | K05953 | nicotianamine synthase | 2.5.1.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spanò, R.; Fortunato, S.; Linsalata, V.; D’Antuono, I.; Cardinali, A.; de Pinto, M.C.; Mascia, T. Comparative Analysis of Bioactive Compounds in Two Globe Artichoke Ecotypes Sanitized and Non-Sanitized from Viral Infections. Plants 2023, 12, 1600. https://doi.org/10.3390/plants12081600
Spanò R, Fortunato S, Linsalata V, D’Antuono I, Cardinali A, de Pinto MC, Mascia T. Comparative Analysis of Bioactive Compounds in Two Globe Artichoke Ecotypes Sanitized and Non-Sanitized from Viral Infections. Plants. 2023; 12(8):1600. https://doi.org/10.3390/plants12081600
Chicago/Turabian StyleSpanò, Roberta, Stefania Fortunato, Vito Linsalata, Isabella D’Antuono, Angela Cardinali, Maria Concetta de Pinto, and Tiziana Mascia. 2023. "Comparative Analysis of Bioactive Compounds in Two Globe Artichoke Ecotypes Sanitized and Non-Sanitized from Viral Infections" Plants 12, no. 8: 1600. https://doi.org/10.3390/plants12081600