
Simplified Genomic Data Revealing the Decline of Aleuritopteris grevilleoides Population Accompanied by the Uplift of Dry-Hot Valley in Yunnan, China

Abstract
:1. Introduction
2. Results
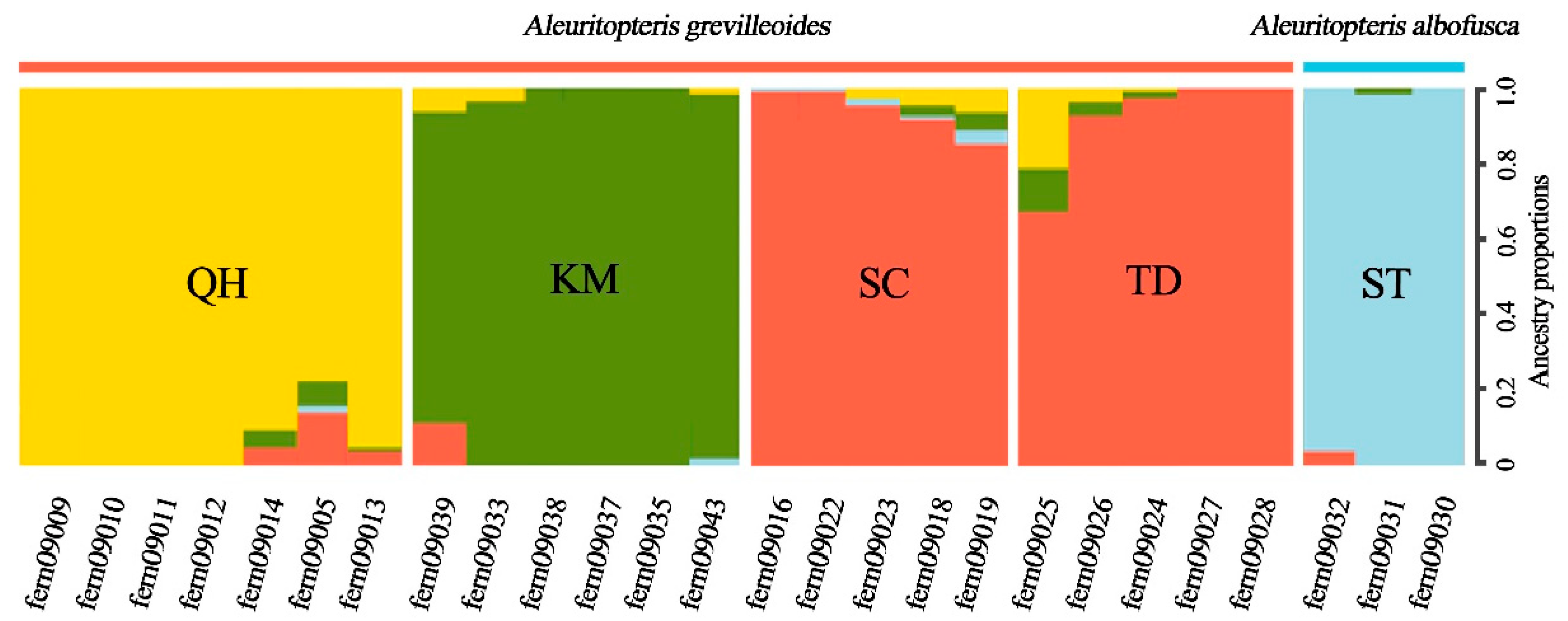
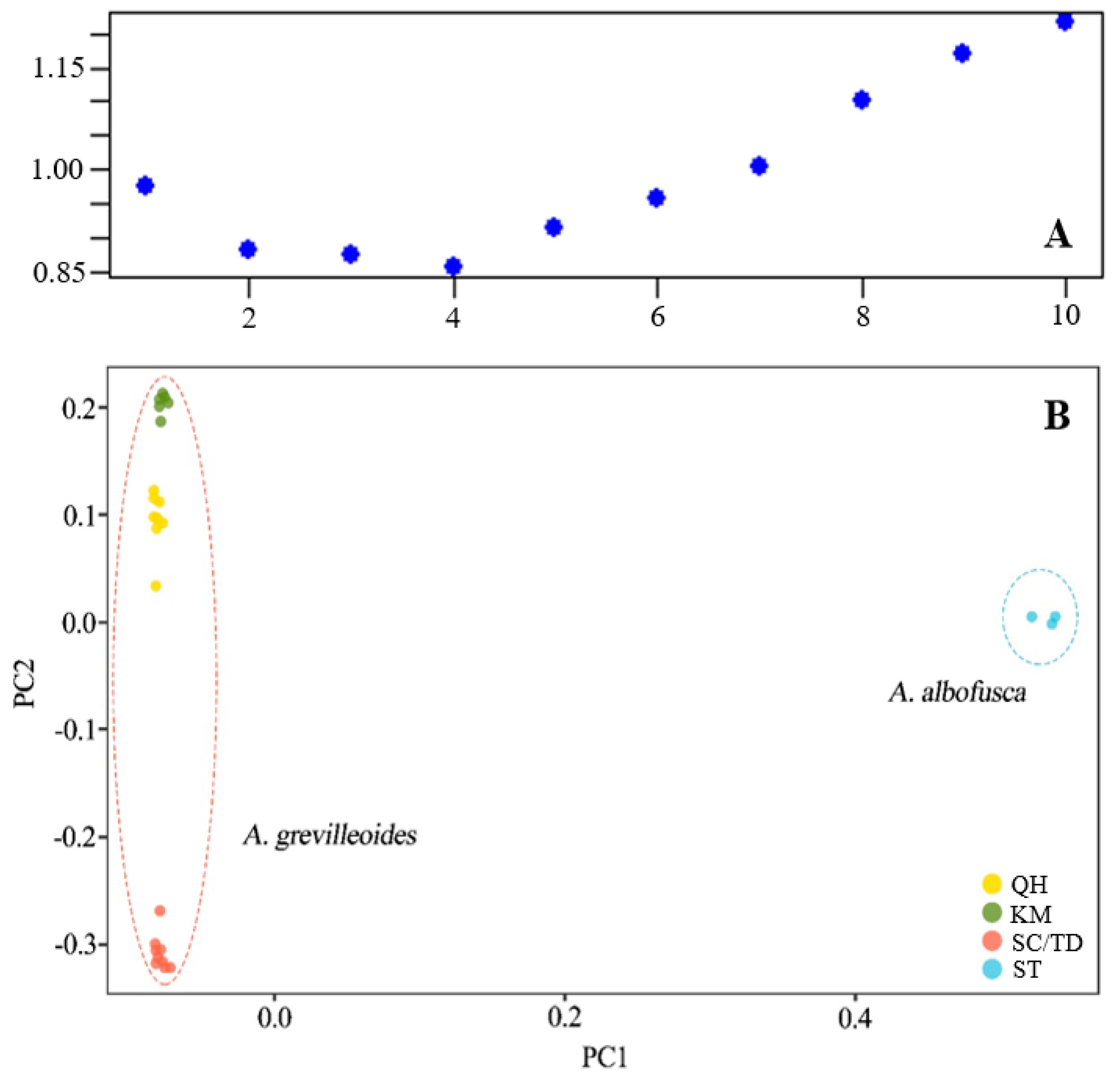
2.1. Genetic Structure
2.2. Genetic Diversity
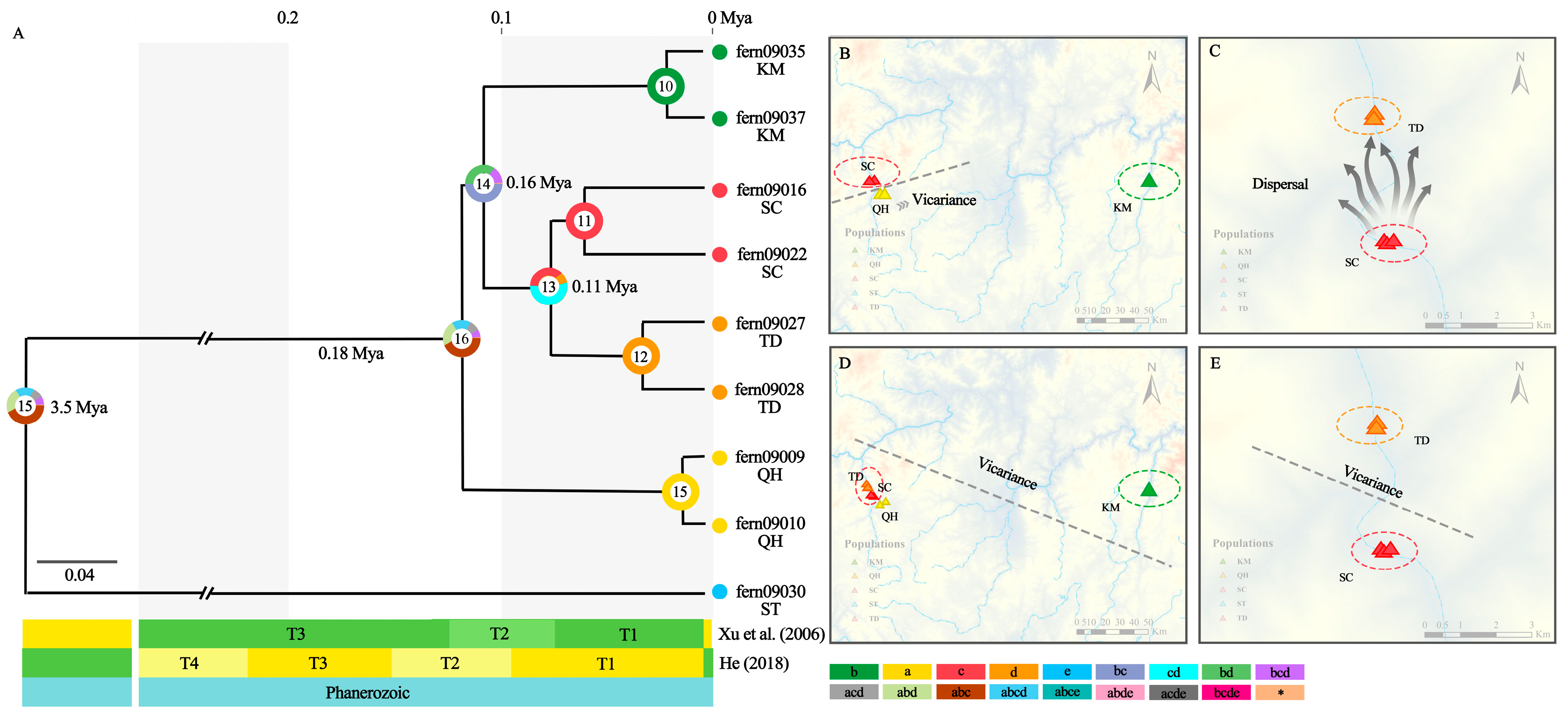
2.3. Divergence Time Estimation and Reconstruction of Ancestral State
2.4. Historical Fluctuation of Effective Population Size
3. Material and Methods
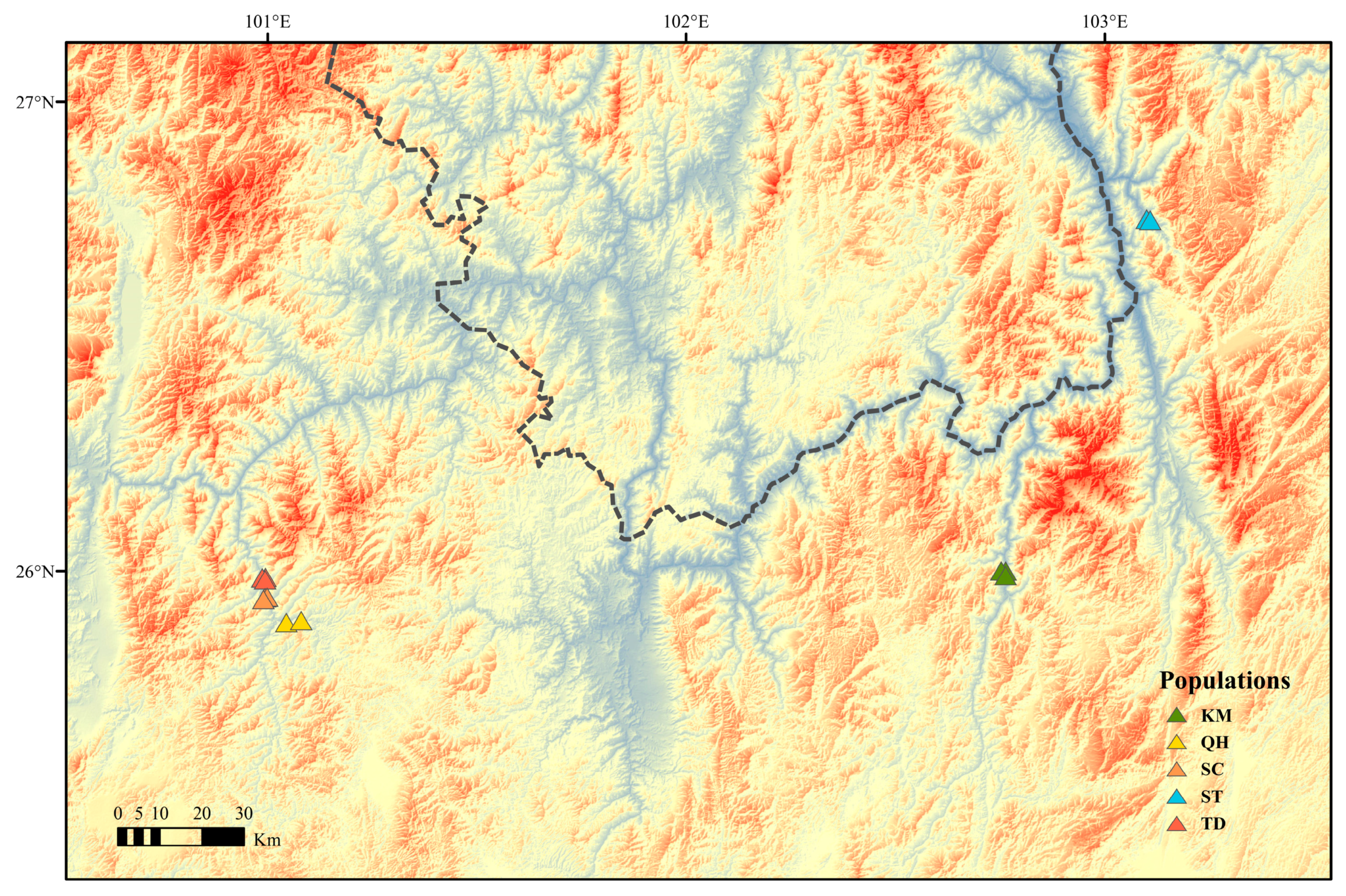
3.1. Plant Material
3.2. DNA Extraction and Genotyping by Sequencing Library Construction
3.3. Determine Single Nucleotide Polymorphisms (SNPs)
3.4. Genetic Diversity and Structure
3.5. Divergence Time and Ancestral State
3.6. Population Demographic History
4. Discussion
4.1. Genetic Diversity of Aleuritopteris Grevilleoides
4.2. Dry-Hot Valleys Are Refuges after A. grevilleoides Population Size Decreases
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antonelli, A.; Kissling, W.D.; Flantua, S.G.A.; Bermúdez, M.A.; Mulch, A.; Muellner-Riehl, A.N.; Kreft, H.; Linder, H.P.; Badgley, C.; Fjeldså, J. Geological and climatic influences on mountain biodiversity. Nat. Geosci. 2018, 11, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Cody, S.; Richardson, J.E.; Rull, V.; Ellis, C.; Pennington, R.T. The great American biotic interchange revisited. Ecography 2010, 33, 326–332. [Google Scholar] [CrossRef]
- Vermeij, G.J. When biotas meet: Understanding biotic interchange. Science 1991, 253, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhou, W.W.; Zhao, H.T.; Yuan, Z.Y.; Wang, Y.Y.; Jiang, K.; Jin, J.Q.; Murphy, R.W.; Che, J.; Zhang, Y.P. Geological events play a larger role than pleistocene climatic fluctuations in driving the genetic structure of Quasipaa boulengeri (Anura: Dicroglossidae). Mol. Ecol. 2013, 22, 1120–1133. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Milne, R.I.; Liu, J.; Slik, F.; Yu, Y.; Luo, Y.H.; Monro, A.K.; Wang, W.T.; Wang, H.; Kessler, P.J.A.; et al. Phylogenomics and evolutionary history of Oreocnide (Urticaceae) shed light on recent geological and climatic events in SE Asia. Mol. Phylogenet. Evol. 2022, 175, 107555. [Google Scholar] [CrossRef]
- Wu, C.Y.; Wu, S.G. A proposal for a new floristic kingdom (realm). In Floristic Characteristics and Diversity of East Asian Plants; Zhang, A.L., Wu, S.G., Eds.; China Higher Education Press: Hong Kong, China; Springer: Berlin/Heidelberg, Germany, 1996; pp. 3–42. [Google Scholar]
- Wen, J.; Zhang, J.Q.; Nie, Z.L.; Zhong, Y.; Sun, H. Evolutionary diversification of plants on the Qinghai-Tibetan Plateau. Front. Genet. 2014, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.B.; Lu, P.Z.; Zhu, T. Causes for the Formation of Dry-Hot Valleys in Hengduan Mountain Yunan Plateau. Resour. Sci. 2000, 22, 69–73. [Google Scholar]
- Zhu, H.; Tan, Y.H.; Yan, L.C.; Liu, F.Y. Flora of the Savanna-like Vegetation in Hot Dry Valleys, Southwestern China with Implications to their Origin and Evolution. Bot. Rev. 2020, 3, 281–297. [Google Scholar] [CrossRef]
- Yang, J.D.; Zhang, Z.M.; Shen, Z.H.; Ou, X.K.; Geng, Y.P.; Yang, M.Y. Review of research on the vegetation and environment of dry-hot valleys in Yunnan. Biodivers. Sci. 2016, 24, 462–474. [Google Scholar] [CrossRef]
- Jin, Z.Z.; Ou, X.K.; Qu, P.D.; Chen, J. Preliminary study on the floristic characteristics of seed plants in the dry-hot valley of Jinsha River. Acta Bot. Yunnanica 1994, 16, 1–16. [Google Scholar]
- Jin, Z.Z.; Yang, Y.P.; Tao, G.D. The floristic characteristics, nature and origin of seed plant in the dry-hot river valley of SW China. Acta Bot. Yunnanica 1995, 17, 129–143. [Google Scholar]
- Ji, Z.H.; Pan, Z.X.; Sha, Y.C.; Fang, H.D.; Liao, C.F.; Bai, D.Z.; Yang, Y.X. Model Construction of Ecological Restoration in Arid Hot Valley of Jinsha River. J. Agro-Environ. Sci. 2006, 25, 716–720. [Google Scholar]
- Díaz, F.P.; Latorre, C.; Carrasco-Puga, G.; Wood, J.R.; Wilmshurst, J.M.; Soto, D.C.; Cole, T.L.; Gutiérrez, R.A. Multiscale climate change impacts on plant diversity in the Atacama Desert. Glob. Change Biol. 2019, 25, 1733–1745. [Google Scholar] [CrossRef]
- Mao, K.S.; Milne, R.; Zhang, L.B.; Peng, Y.L.; Liu, J.Q.; Thomas, P.; Mill, R.; Renner, S.S. Distribution of living Cupressaceae reflects the breakup of Pangea. Proc. Natl. Acad. Sci. USA 2012, 109, 7793–7798. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.L.; Xia, Y.M.; Cannon, C.H.; Kress, W.J.; Li, Q.J. Evolutionary diversification of alpine ginger reflects the early uplift of the Himalayan-Tibetan Plateau and rapid extrusion of Indochina. Gondwana Res. 2016, 32, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Sun, H. Phylogeographic structure of Terminalia franchetii (combretaceae) in southwest china and its implications for drainage geological history. J. Plant Res. 2011, 124, 63–73. [Google Scholar] [CrossRef]
- Chen, L.H.; Hu, T.X.; Zhang, F. AFLP analysis on genetic diversity of Juglans populations in dry and dry-hot valleys of Sichuan province. J. Fruit Sci. 2009, 26, 48–54. [Google Scholar]
- Dong, S.Y.; Zuo, Z.Y.; Yan, Y.H.; Xiang, J.Y. Red list assessment of lycophytes and ferns in China. Biodivers. Sci. 2017, 25, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.N.; Yang, Y.; Dong, S.Y.; He, Q. Threatened species list of China’s higher plants. Biodivers. Sci. 2017, 25, 696–744. [Google Scholar] [CrossRef] [Green Version]
- Christensen, C.; Chink, R.C. Sinopteris, a New Fern Genus in China. Bull. Fan Mem. Inst. Bot. 1933, 4, 355–364. [Google Scholar]
- National Forestry Administration and Ministry of Agriculture of the People’s Republic of China. 1999; List of National Key Preserved Wild Plants (Batch 1) [EB/OL]. Available online: http://www.gov.cn/gongbao/content/2000/content_60072.htm (accessed on 20 March 2023).
- National Forestry and Grassland Administration of the People’s Republic of China. 2021; List of National Key Preserved Wild Plants, EB/OL. Available online: http://www.gov.cn/zhengce/zhengceku/2021-09/09/content_5636409.htm (accessed on 9 September 2021).
- Wang, W.Q.; Zhu, H.; Peng, Z.J.; Sun, C.S.; Cheng, X. Resource Distribution, Protection Status and Reasonable Utilization of Sinopteris. In China Botanical Garden; China Forestry Publishing House: Beijing, China, 2010; pp. 158–163. [Google Scholar]
- Wang, W.Q.; Cheng, X.; Jiao, Y.; Chen, G.J. Substratum Affects the Gametophyte Development and Reproduction of Sinopteris grevilleoides (Sinopteridaceae) an Endangered Fern Endemic to China. Acta Bot. Yunnanica 2010, 32, 25–31. [Google Scholar] [CrossRef]
- Yuan, Q.; Liu, Y.B.; Jin, D.M.; Song, Y.G.; Morigengowa; Gu, Y.F.; Wang, A.H.; Xiang, J.Y.; Dai, X.L.; Yan, Y.H. Three endangered Pteridaceae species in China: Current population and conservation suggestion. Chin. J. Ecol. 2021, 42, e02396. [Google Scholar]
- Zhong, X.H. Ecosystem degradation, restoration and reconstruction approaches in hot-dry valley. Resour. Environ. Yang Tze Basin 2000, 9, 1–4. [Google Scholar]
- Xu, Q.M.; Yang, D.Y.; Ge, Z.S.; Chen, D.J.; Xu, G.F. Terraces along Sanduizi-Wudongde section of Jinsha Rive. Sci. Geogr. Sin. 2006, 26, 609–615. [Google Scholar]
- He, R. Study on the Formation Age of the Dry Hot Valley Based on River Terrace Records in the Qiaojia Section of Jinsha River. Master’s Thesis, Yunnan Normal University, Yunnan, China, 2018. [Google Scholar]
- Qin, R.C.; Xing, G.X. Flora Reipublicae Popularis Sinicae. Typis Acad. Sci. Sin. 1990, 3, 180. [Google Scholar]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Le´gare, G.; Boyle, B.; Normandeau, E.; Laroche, J.; Larose, S.; Jean, M.; et al. An improved genotyping by sequencing (gbs) approach offering increased versatility and efficiency of snp discovery and genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef] [Green Version]
- Rochette, N.; Rivera-Colón, A.; Catchen, J. Stacks 2: Analytical methods for paired-end sequencing improve RADseq-based population genomics. Mol. Ecol. 2019, 28, 4737–4754. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GIGASCIENCE 2015, 4, 7. [Google Scholar] [CrossRef]
- Frichot, E.; Francois, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Galinsky, K.J.; Bhatia, G.; Loh, P.R.; Georgiev, S.; Mukherjee, S.; Patterson, N.J.; Price, A.L. Fast Principal-Component Analysis Reveals Convergent Evolution of ADH1B in Europe and East Asia. Am. J. Hum. Genet. 2016, 98, 456–472. [Google Scholar] [CrossRef] [Green Version]
- Catchen, J.; Hohenlohe, P.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.M.; Gammerdinger, J.W.; Bartie, L.K.; Penman, D.J.; Kocher, T.D. A high quality assembly of the Nile Tilapia (Oreochromis niloticus) genome reveals the structure of two sex determination regions. BMC Genom. 2017, 18, 341. [Google Scholar] [CrossRef]
- Stange, M.; Sanchez-Villagra, M.R.; Salzburger, W.; Matschiner, M. Bayesian divergence-time estimation with genome-wide single-nucleotide polymorphism data of sea catfishes (ariidae) supports miocene closure of the panamanian isthmus. Syst. Biol. 2018, 4, 681–699. [Google Scholar] [CrossRef] [Green Version]
- Nitta, J.H.; Schuettpelz, E.; Ramírez-Barahona, S.; Iwasaki, W. An open and continuously updated fern tree of life. Front. Plant Sci. 2022, 13, 909768. [Google Scholar] [CrossRef]
- Testo, W.; Sundue, M. A 4000-species dataset provides new insight into the evolution of ferns. Mol. Phylogenet Evol. 2016, 105, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Blair, C.; He, X.J. RASP 4: Ancestral State Reconstruction Tool for Multiple Genes and Characters. Mol. Biol. Evol. 2020, 37, 604–606. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X.J. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Biol. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Liu, X.; Fu, Y.X. Exploring population size changes using SNP frequency spectra. Nat Genet. 2015, 47, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Fu, Y.X. Stairway Plot 2: Demographic history inference with folded SNP frequency spectra. Genome Biol. 2020, 21, 280. [Google Scholar] [CrossRef]
- Cristofari, R.; Liu, X.; Bonadonna, F.; Cherel, Y.; Pistorius, P.; Maho, Y.L.; Raybaud, V.; Stenseth, N.C.; Bohec, C.L.; Trucchi, E. Climate-driven range shifts of the king penguin in a fragmented ecosystem. Nat. Clim. Change 2018, 8, 245–251. [Google Scholar] [CrossRef]
- Gitzendanner, M.A.; Soltis, P.S. Patterns of genetic variation in rare and widespread plant congeners. Am. J. Bot. 2000, 87, 783. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.Y.; Shu, J.P.; Gu, Y.F.; Morigengaowa. Conservation genomics analysis revealed the endangered mechanism of Adiantum nelumboides. Biodivers. Sci. 2022, 30, 1–12. [Google Scholar] [CrossRef]
- Murray, G.G.R.; Soares, A.E.R.; Novak, B.J.; Schaefer, N.K.; Cahill, J.A.; Baker, A.J.; Demboski, J.R.; Doll, A.; Fonseca, R.R.D.; Fulton, T.L. Natural selection shaped the rise and fall of passenger pigeon genomic diversity. Science 2017, 358, 951–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelke, M.E.; Martenson, J.S.; O’Brien, S.J. The consequences of demographic reduction and genetic depletion in the endangered florida panther. Curr. Biol. 1993, 6, 340–350. [Google Scholar] [CrossRef]
- Willoughby, J.R.; Sundaram, M.; Wijayawardena, B.K.; Kimble, S.J.A.; Ji, Y.Z.; Fernandez, N.B.; Antonides, J.D.; Lamb, M.C.; Marra, N.J.; DeWoody, J.A. The reduction of genetic diversity in threatened vertebrates and new recommendations regarding iucn conservation rankings. Biol. Conserv. 2015, 191, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Hu, Y.; Wu, Q.; Nie, Y.G.; Yan, L.; Wei, F.W. Conservation genetics and genomics of threatened vertebrates in China. J. Genet. Genom. 2018, 11, 593–601. [Google Scholar] [CrossRef]
- Ehlers, E.; Krafft, T. The “Anthropocene”. Earth System Science in the Anthropocene; Springer: Berlin/Heidelberg, Germany, 2006; pp. 13–18. [Google Scholar]
- Su, H.; Ming, Q.Z.; Pan, B.T.; Gao, H.S.; Zhang, W.X.; Dong, M.; Shi, Z.T. Jinsha River Valley-Analysis and Discussion on Chronological Framework of Water System Development. J. Mt. Sci. 2013, 31, 685–692. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QH | SC | TD | KM | |
|---|---|---|---|---|
| SC | 0.225 | |||
| TD | 0.188 | 0.031 | ||
| KM | 0.226 | 0.142 | 0.116 | |
| ST | 0.684 | 0.621 | 0.543 | 0.579 |
| FID | Private | Obs_Het | Obs_Hom | Exp_Het | Exp_Hom | Pi | Fis |
|---|---|---|---|---|---|---|---|
| QH | 1016 | 0.124 | 0.876 | 0.146 | 0.854 | 0.159 | 0.084 |
| SC | 439 | 0.182 | 0.818 | 0.222 | 0.778 | 0.251 | 0.144 |
| TD | 461 | 0.216 | 0.784 | 0.237 | 0.763 | 0.268 | 0.108 |
| KM | 995 | 0.159 | 0.841 | 0.191 | 0.809 | 0.211 | 0.114 |
| A. grevilleoides | 0 | 0.166 | 0.834 | 0.322 | 0.678 | 0.333 | 0.452 |
| Specie | No. | Specimen Copy | Date | Site | Abbr |
|---|---|---|---|---|---|
| A. albofusca | fern09030 | 1 | 14 August 2019 | Qiaojia County | ST |
| A. albofusca | fern09031 | 1 | 14 August 2019 | Qiaojia County | ST |
| A. albofusca | fern09032 | 1 | 14 August 2019 | Qiaojia County | ST |
| A. grevilleoides | fern09005 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09009 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09010 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09011 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09012 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09013 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09014 | 1 | 13 August 2019 | Qihei Highway of Dayao County | QH |
| A. grevilleoides | fern09016 | 1 | 13 August 2019 | SanCha River of Dayao County | SC |
| A. grevilleoides | fern09018 | 1 | 13 August 2019 | SanCha River of Dayao County | SC |
| A. grevilleoides | fern09019 | 1 | 13 August 2019 | SanCha River of Dayao County | SC |
| A. grevilleoides | fern09022 | 1 | 13 August 2019 | SanCha River of Dayao County | SC |
| A. grevilleoides | fern09023 | 1 | 13 August 2019 | SanCha River of Dayao County | SC |
| A. grevilleoides | fern09024 | 1 | 13 August 2019 | Tademe Bridge of Dayao County | TD |
| A. grevilleoides | fern09025 | 1 | 13 August 2019 | Tademe Bridge of Dayao County | TD |
| A. grevilleoides | fern09026 | 1 | 13 August 2019 | Tademe Bridge of Dayao County | TD |
| A. grevilleoides | fern09027 | 1 | 13 August 2019 | Tademe Bridge of Dayao County | TD |
| A. grevilleoides | fern09028 | 1 | 13 August 2019 | Tademe Bridge of Dayao County | TD |
| A. grevilleoides | fern09033 | 1 | 17 August 2019 | Luquan County of KunMing City | KM |
| A. grevilleoides | fern09035 | 1 | 17 August 2019 | Luquan County of KunMing City | KM |
| A. grevilleoides | fern09037 | 1 | 17 August 2019 | Luquan County of KunMing City | KM |
| A. grevilleoides | fern09038 | 1 | 17 August 2019 | Luquan County of KunMing City | KM |
| A. grevilleoides | fern09039 | 1 | 17 August 2019 | Luquan County of KunMing City | KM |
| A. grevilleoides | fern09043 | 1 | 17 August 2019 | Luquan County of KunMing City | KM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.-Y.; Wang, T.; Zhou, J.; Sun, W.-Y.; Jin, D.-M.; Xiang, J.-Y.; Shao, J.-W.; Yan, Y.-H. Simplified Genomic Data Revealing the Decline of Aleuritopteris grevilleoides Population Accompanied by the Uplift of Dry-Hot Valley in Yunnan, China. Plants 2023, 12, 1579. https://doi.org/10.3390/plants12071579
Wei X-Y, Wang T, Zhou J, Sun W-Y, Jin D-M, Xiang J-Y, Shao J-W, Yan Y-H. Simplified Genomic Data Revealing the Decline of Aleuritopteris grevilleoides Population Accompanied by the Uplift of Dry-Hot Valley in Yunnan, China. Plants. 2023; 12(7):1579. https://doi.org/10.3390/plants12071579
Chicago/Turabian StyleWei, Xue-Ying, Ting Wang, Jin Zhou, Wei-Yue Sun, Dong-Mei Jin, Jian-Ying Xiang, Jian-Wen Shao, and Yue-Hong Yan. 2023. "Simplified Genomic Data Revealing the Decline of Aleuritopteris grevilleoides Population Accompanied by the Uplift of Dry-Hot Valley in Yunnan, China" Plants 12, no. 7: 1579. https://doi.org/10.3390/plants12071579