
Nuclear DNA Amounts in Chinese Bryophytes Estimated by Flow Cytometry: Variation Patterns and Biological Significances




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. General Aspects
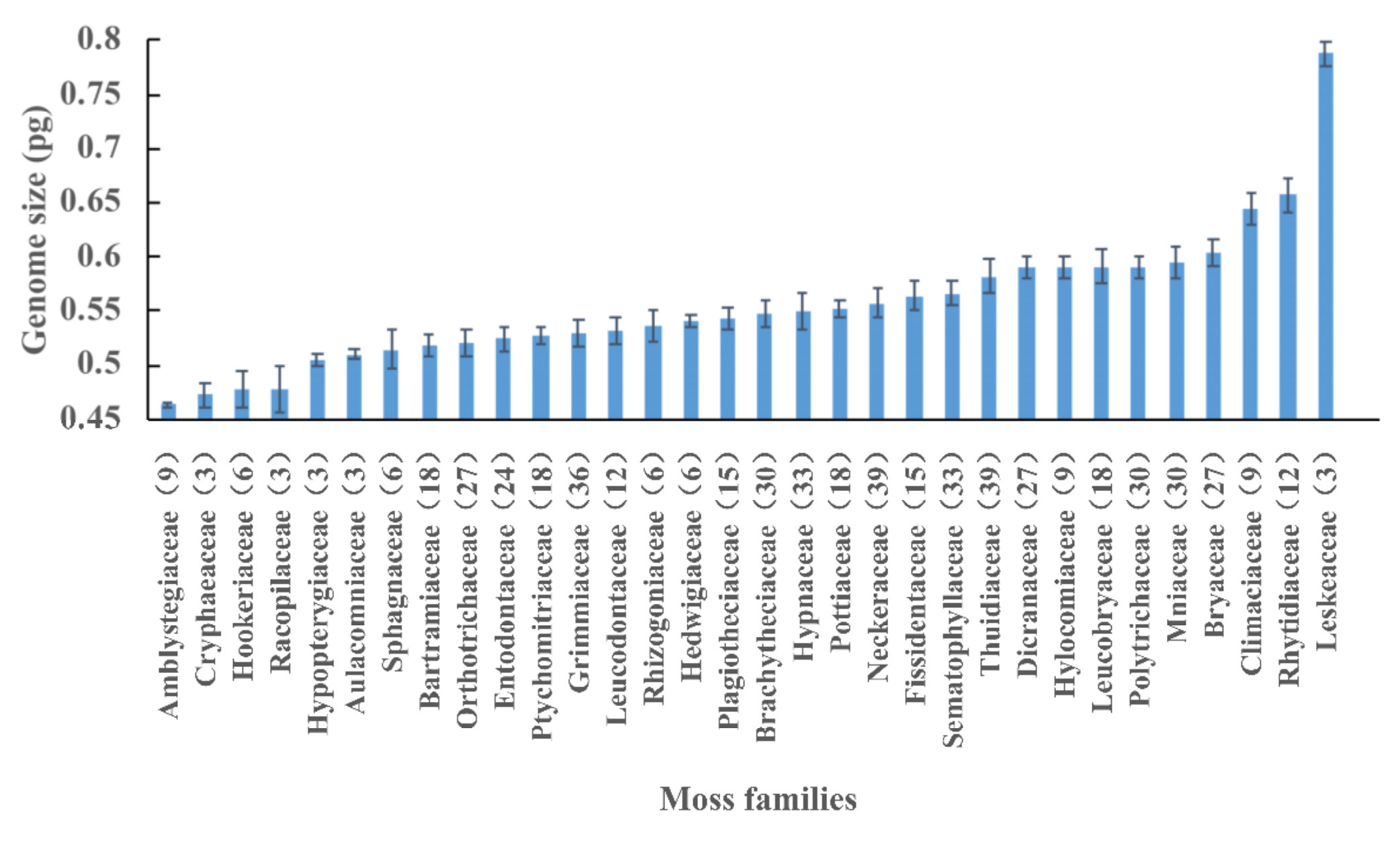
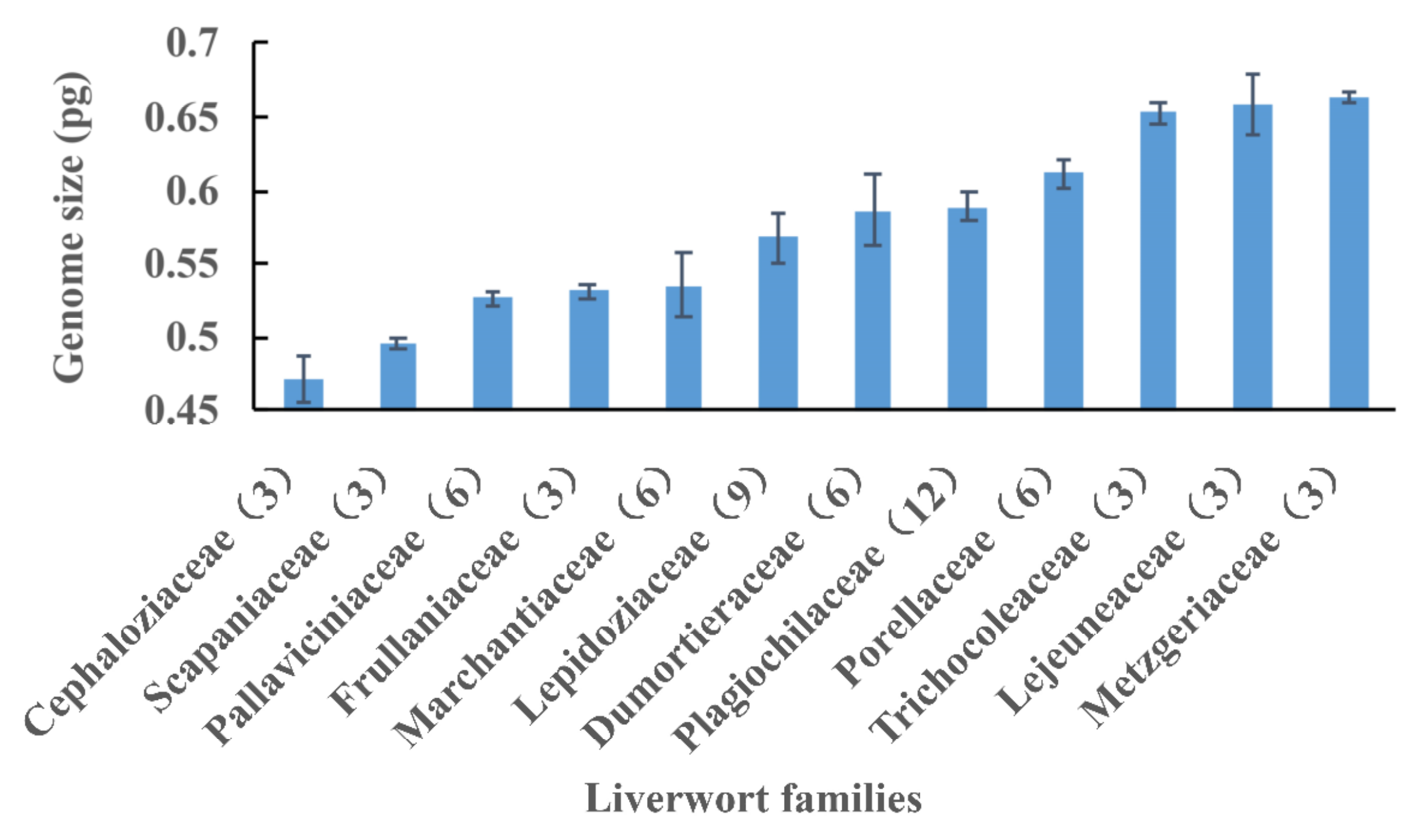
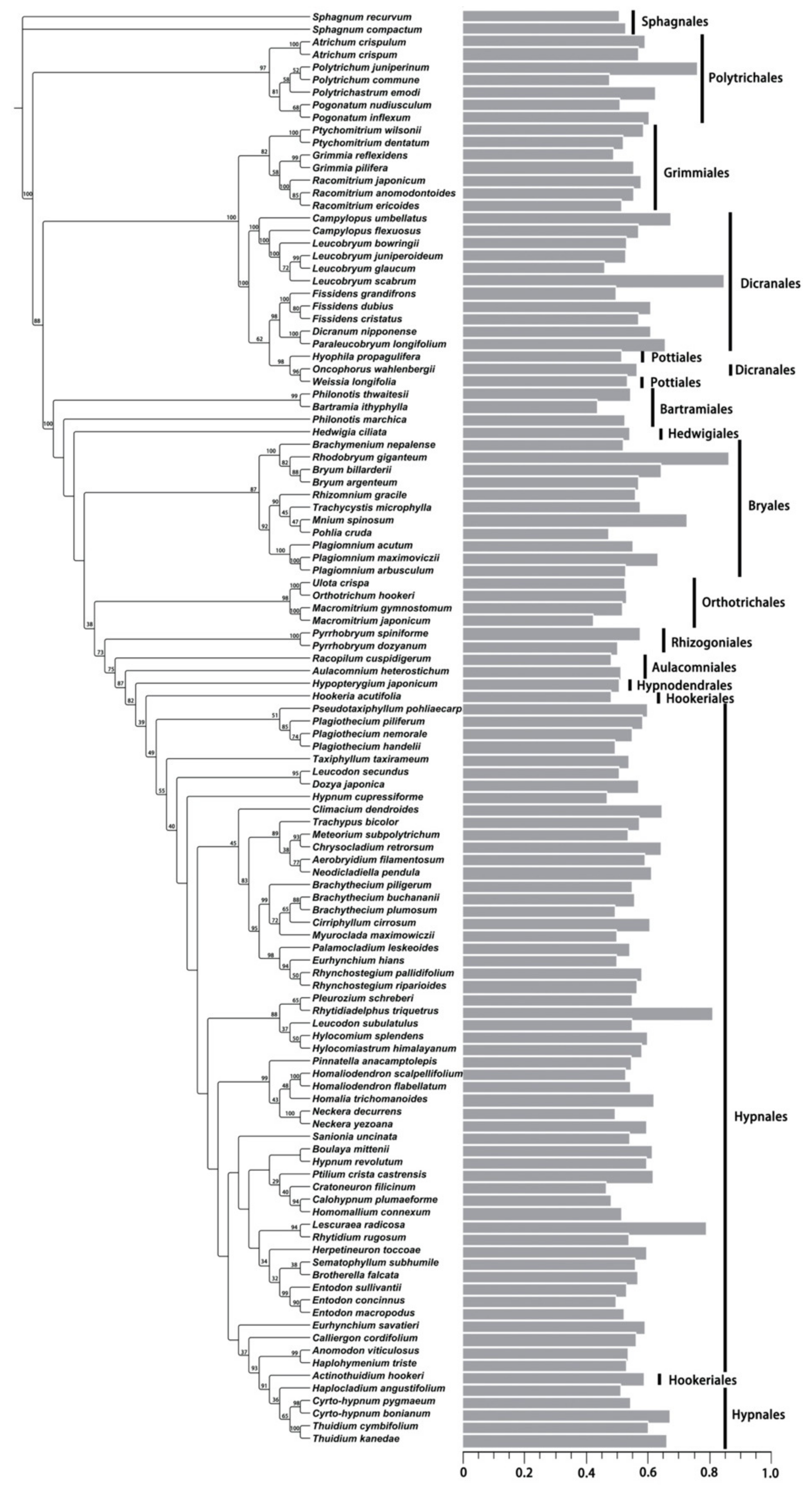
2.2. Variation Pattern of Nuclear DNA Amounts
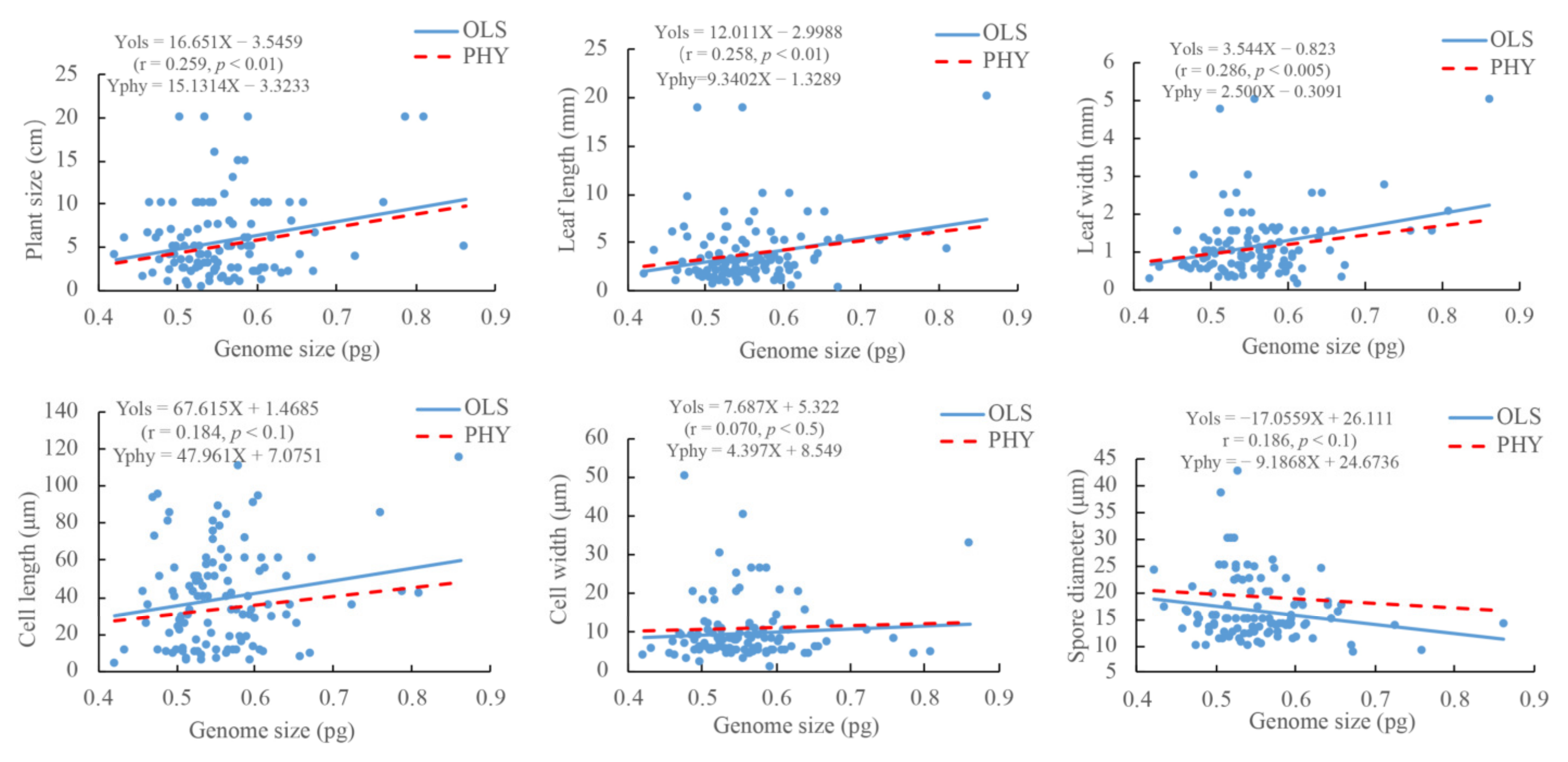
2.3. Relationships between DNA 1C Values and Morphological Traits
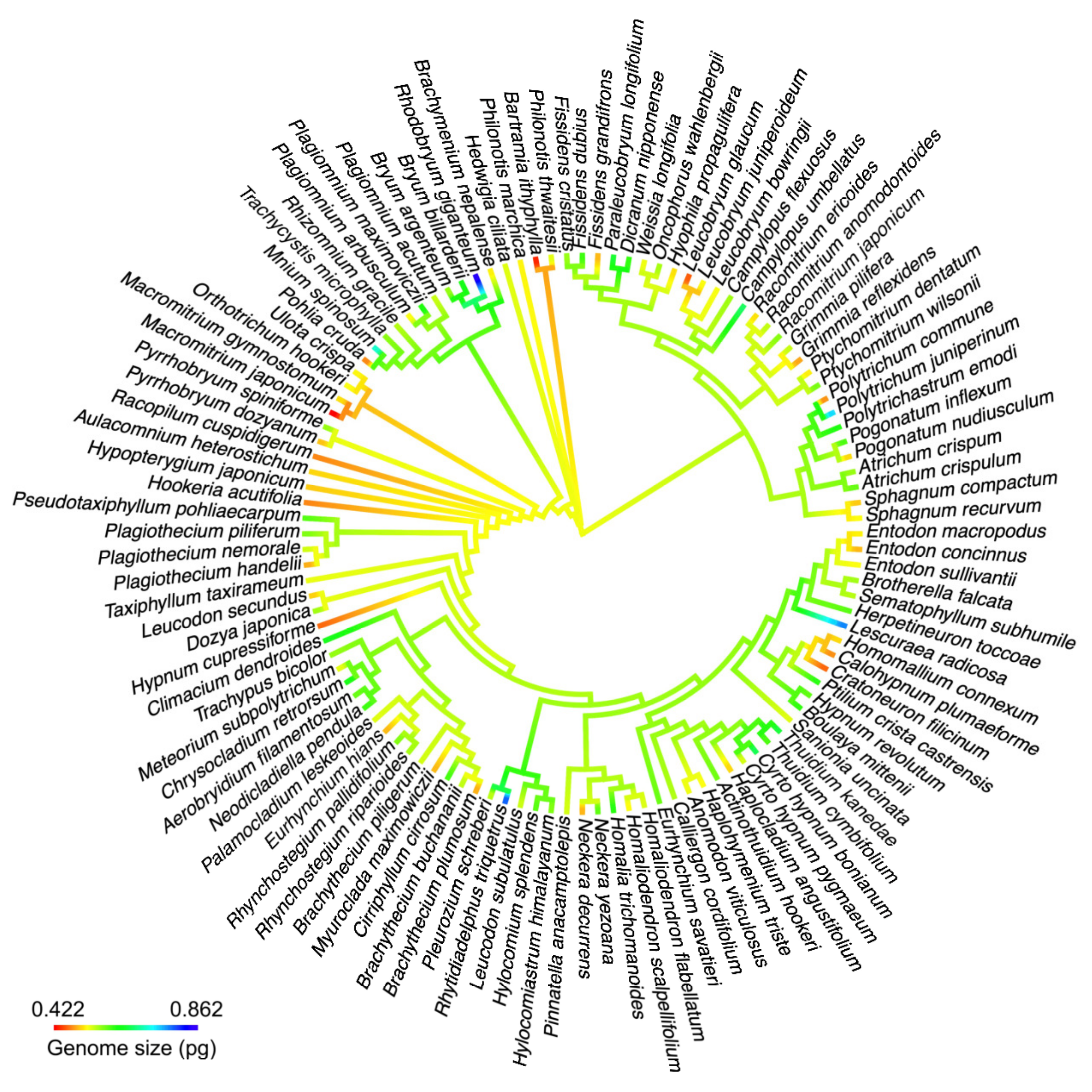
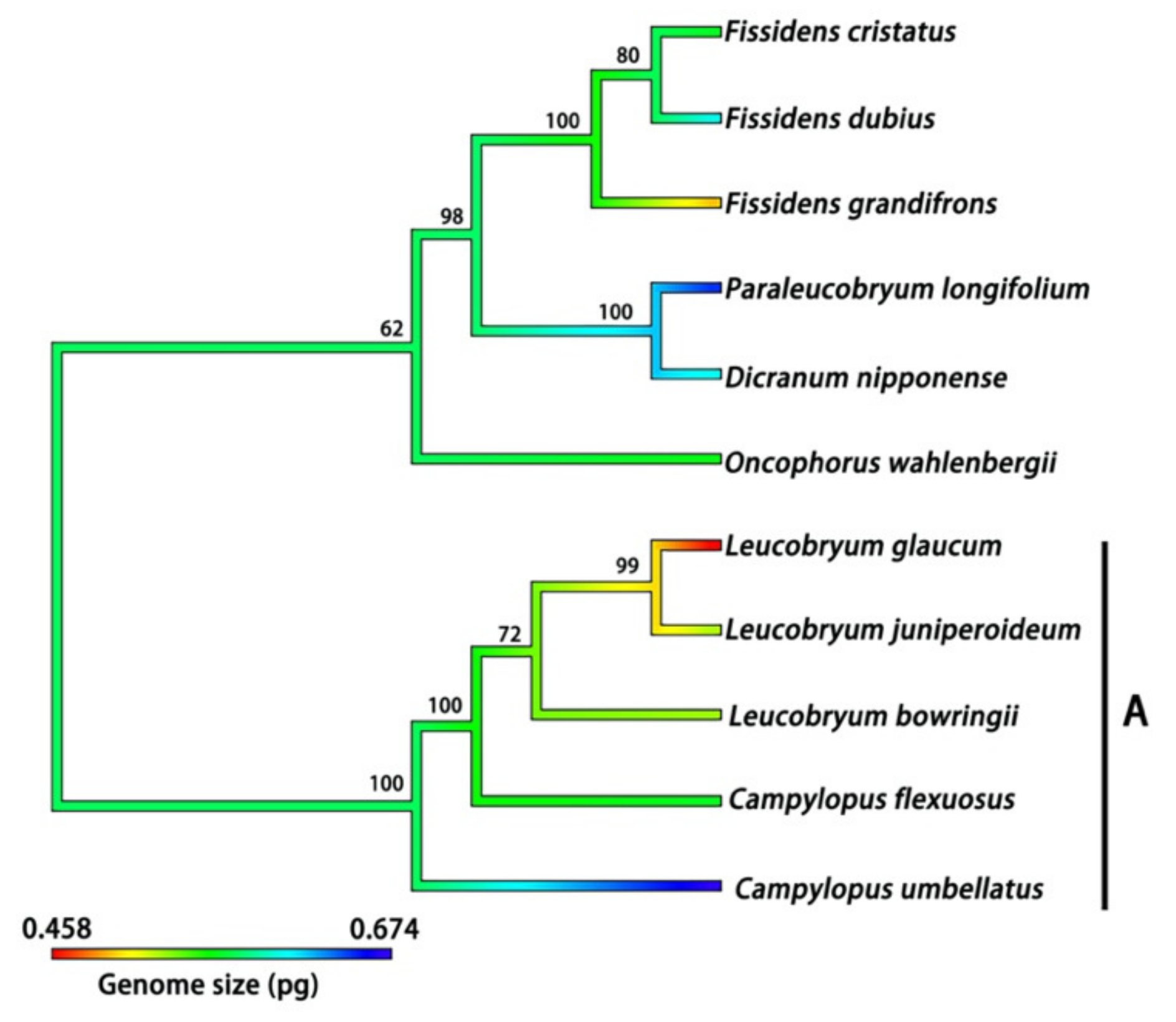
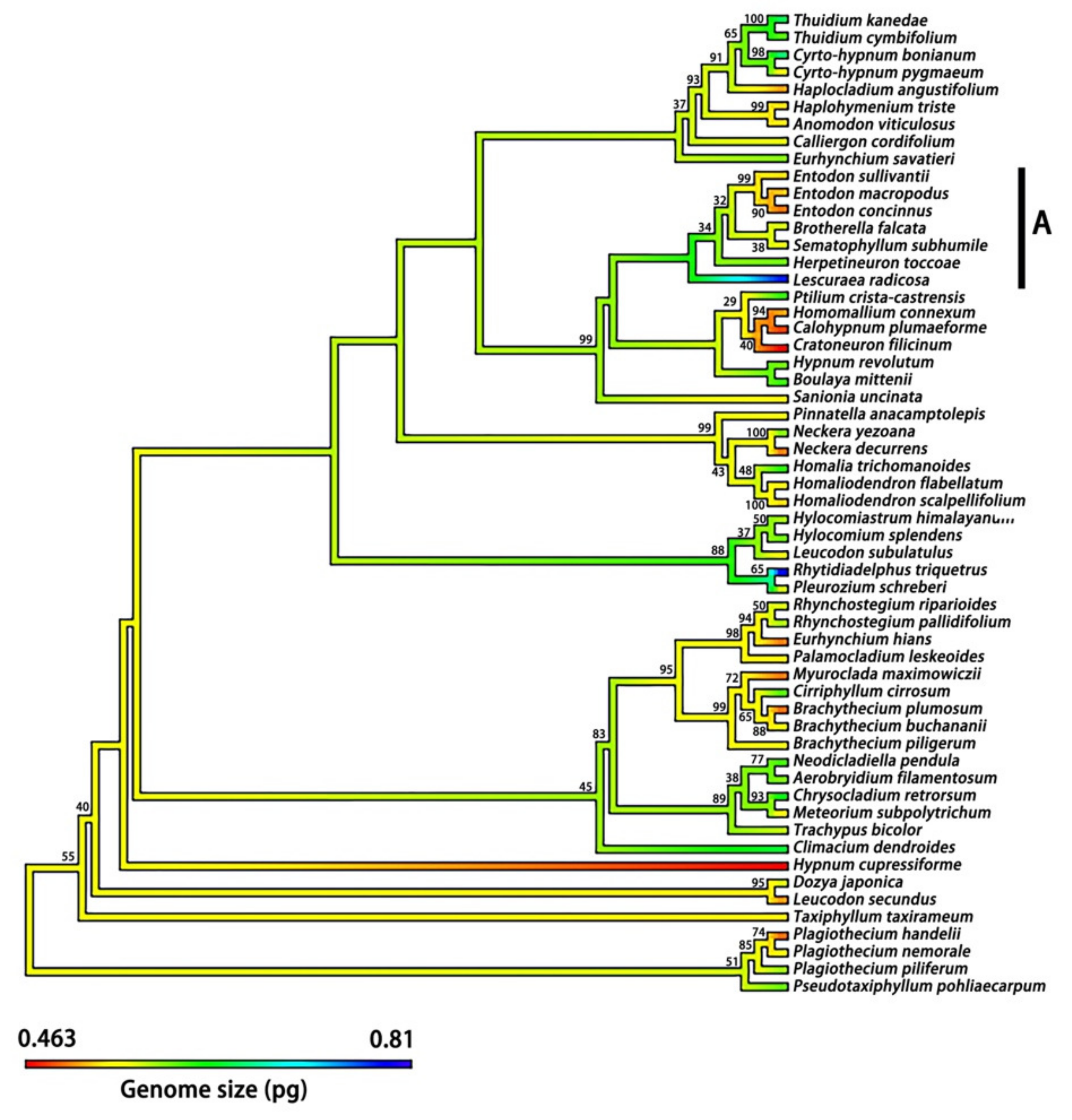
2.4. Phylogenetic Signal for DNA 1C Value among Moss Taxa
3. Discussion
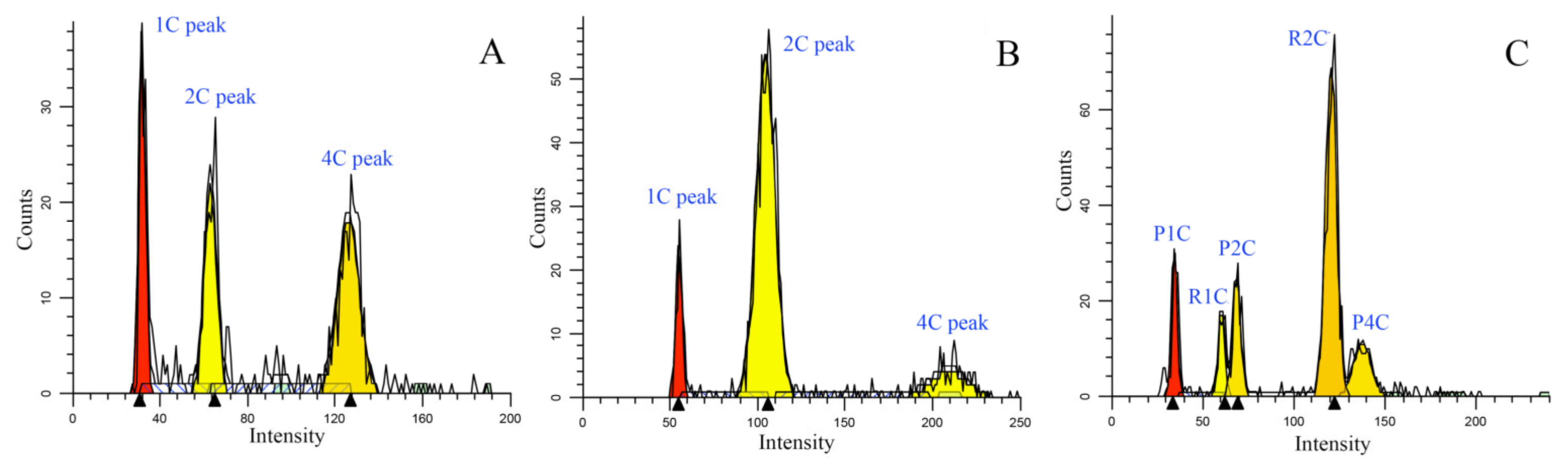
3.1. Credibility of the Nuclear DNA Amounts We Determined
3.2. Variation Patterns of the Nuclear DNA Amounts in Bryophytes
3.3. Nucleotype Effects and Possible Ecological Significance of Nuclear DNA Amounts in Bryophytes
3.4. Phylogenetic Signals of Nuclear DNA Amounts in Bryophytes
4. Materials and Methods
4.1. Materials
4.2. Nuclei Isolation
4.3. Nuclear DNA Amount Measurement
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Plant Materials, Voucher Number, Geographic Sources and DNA 1C Values (Means and Standard Errors)
| Families | Species | Localities | Lat (˚) | Lon (˚) | Ele (m) | Voucher Number | Genome Size Means (pg) | Genome Size SEs (pg) |
| Sphagnaceae | Sphagnum compactum Lam. & DC. | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.669 | 119.48 | 480 | 20170617098 | 0.504 | 0.023 |
| Sphagnaceae | Sphagnum recurvum P. Beauv. | Meiduozegong, Jianziwan, Yajiang, Sichuan Province | 29.348 | 100.113 | 4370 | 20170807033 | 0.526 | 0.014 |
| Polytrichaceae | Atrichum crispulum Schimp. ex Besch. * | Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.428 | 1204 | 20170630022 | 0.59 | 0.006 |
| Polytrichaceae | Atrichum crispum (James) Sull. * | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170503003 | 0.561 | 0.006 |
| Polytrichaceae | Atrichum crispum (James) Sull. * | near Jinhai Heavy Industry, Daishan, Zhejiang Province | 30.244 | 122.263 | 33 | 1-20170530063 | 0.577 | 0.007 |
| Polytrichaceae | Pogonatum inflexum (Lindb.) Sande Lac. * | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.239 | 122.397 | 20 | 2-20170524037 | 0.603 | 0.019 |
| Polytrichaceae | Pogonatum inflexum (Lindb.) Sande Lac. * | Mt. Foding, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.001 | 122.352 | 161 | 1-20170424282 | 0.601 | 0.008 |
| Polytrichaceae | Pogonatum nudiusculum (Müll. Hal.) T.J. Kop. * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.576 | 102.001 | 2965 | 20170802052 | 0.508 | 0.004 |
| Polytrichaceae | Polytrichastrum emodi G.L. Sm. ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.587 | 101.97 | 3505 | 20170801032 | 0.712 | 0.01 |
| Polytrichaceae | Polytrichastrum emodi G.L. Sm. ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.587 | 101.97 | 3505 | 20170801034 | 0.533 | 0.012 |
| Polytrichaceae | Polytrichum commune Hedw. * | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170503002 | 0.474 | 0.005 |
| Polytrichaceae | Polytrichum juniperinum Hedw. | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.67 | 119.482 | 440 | 20170617081 | 0.76 | 0.019 |
| Ptychomitriaceae *** | Ptychomitrium dentatum (Mitt.) A. Jaeger ** | Upstairs, Mt. Xianren, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.028 | 121.906 | 229 | 20170221027 | 0.463 | 0.013 |
| Ptychomitriaceae *** | Ptychomitrium dentatum (Mitt.) A. Jaeger ** | Entrance, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.651 | 102.115 | 1594 | 20170731042 | 0.573 | 0.004 |
| Ptychomitriaceae *** | Ptychomitrium linearifolium Reim. & Sak. ** | Roadside, Taizi Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.325 | 119.45 | 408 | 20170627116 | 0.505 | 0.003 |
| Ptychomitriaceae *** | Ptychomitrium polyphylloides (Müll. Hal.) Paris ** | Camp 1, Hailuogou, Moxi Town, Luding County, Sichuan Province | 29.603 | 102.074 | 1900 | 20170803182 | 0.544 | 0.003 |
| Ptychomitriaceae *** | Ptychomitrium tortula (Harv.) A. Jaeger ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.573 | 102.003 | 2462 | 20170802118 | 0.496 | 0.007 |
| Ptychomitriaceae *** | Ptychomitrium wilsonii Sull. & Lesq. ** | Underbridge, Stele, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.447 | 337 | 20170627118 | 0.585 | 0.017 |
| Grimmiaceae | Grimmia pilifera P. Beauv. ** | Mt. Foding, Mt. Putuo, Zhoushan District, Zhejiang Province | 29.992 | 122.383 | 97 | 1-20170424299 | 0.512 | 0.012 |
| Grimmiaceae | Grimmia pilifera P. Beauv. ** | Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.35 | 119.424 | 1484 | 20170630052 | 0.591 | 0.015 |
| Grimmiaceae | Grimmia reflexidens Müll. Hal. ** | Roadside Daocheng Yading, Sichuan Province | 28.764 | 100.277 | 4030 | 20170805060 | 0.488 | 0.005 |
| Grimmiaceae | Racomitrium anomodontoides Cardot ** | Tiesuo Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.671 | 119.481 | 570 | 20170617171 | 0.554 | 0.027 |
| Grimmiaceae | Racomitrium anomodontoides Cardot ** | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.662 | 119.479 | 630 | 20170617148 | 0.578 | 0.012 |
| Grimmiaceae | Racomitrium anomodontoides Cardot ** | Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.428 | 1204 | 20170630009 | 0.563 | 0.012 |
| Grimmiaceae | Racomitrium anomodontoides Cardot ** | Simian Peak, Mt. Xitianmu, Linan, Zhejiang Province | 30.341 | 119.434 | 1087 | 20170710141 | 0.516 | 0.037 |
| Grimmiaceae | Racomitrium ericoides (Brid.) Brid. ** | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.264 | 122.36 | 45 | 2-20170529120 | 0.509 | 0.019 |
| Grimmiaceae | Racomitrium ericoides (Brid.) Brid. ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.553 | 101.97 | 3505 | 20170801043 | 0.518 | 0.002 |
| Grimmiaceae | Racomitrium japonicum Dozy & Molk. ** | Fayu Temple to Fanyin Cave, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.004 | 122.401 | 56 | 2-20170424085 | 0.577 | 0.005 |
| Grimmiaceae | Schistidium striatum Brid. * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.577 | 102.002 | 2983 | 20170802103 | 0.442 | 0.008 |
| Grimmiaceae | Schistidium subconfertum (Broth.) Deguchi * | Downhill Telpher Station, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.552 | 101.97 | 3557 | 20170801124 | 0.501 | 0.002 |
| Dicranaceae | Campylopus flexuosus (Hedw.) Brid. ** | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.661 | 119.311 | 690 | 20170617148 | 0.567 | 0.014 |
| Dicranaceae | Campylopus umbellatus (Arn.) Paris * | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.239 | 122.397 | 45 | 20170529033 | 0.674 | 0.013 |
| Dicranaceae | Dicranodontium tenii Broth. et Herz. * | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 630 | 20170617127 | 0.653 | 0.021 |
| Dicranaceae | Dicranum gymnostomum Mitt. * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.577 | 102.002 | 2983 | 20170802083 | 0.58 | 0.008 |
| Dicranaceae | Dicranum kashmirense Broth. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.668 | 119.481 | 480 | 20170617101 | 0.517 | 0.01 |
| Dicranaceae | Dicranum kashmirense Broth. * | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.671 | 119.485 | 450 | 20170617040 | 0.586 | 0.011 |
| Dicranaceae | Dicranum nipponense Besch. * | Roadside from Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.399 | 1103 | 20170630025 | 0.608 | 0.006 |
| Dicranaceae | Oncophorus wahlenbergii Brid. ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.571 | 101.998 | 2940 | 20170802150 | 0.62 | 0.005 |
| Dicranaceae | Oncophorus wahlenbergii Brid. ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.576 | 102.072 | 2965 | 20170802028 | 0.508 | 0.008 |
| Leucobryaceae *** | Leucobryum bowringiii Mitt. ** | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.674 | 119.5 | 420 | 20170617015 | 0.463 | 0.018 |
| Leucobryaceae *** | Leucobryum bowringiii Mitt. ** | Up Stairs, Chanyuan Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.325 | 119.445 | 377 | 20170627113 | 0.596 | 0.002 |
| Leucobryaceae *** | Leucobryum glaucum (Hedw.) Ångstr. ** | Up Stairs, Chanyuan Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.325 | 119.445 | 377 | 20170627113-1 | 0.458 | 0.019 |
| Leucobryaceae *** | Leucobryum juniperoideum (Brid.) Müll. Hal. ** | Xiaochangtu, Daishan, Zhoushan District, Zhejiang Province | 30.244 | 122.262 | 43 | 1-2017053078 | 0.527 | 0.018 |
| Leucobryaceae *** | Leucobryum scabrum Sande Lac. ** | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.671 | 119.485 | 450 | 20170617037 | 0.846 | 0.025 |
| Leucobryaceae *** | Paraleucobryum longifolium (Ehrh. ex Hedw.) Loeske ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.587 | 101.97 | 3505 | 20170801061 | 0.655 | 0.019 |
| Fissidentaceae | Fissidens cristatus Wilson ex Mitt. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.67 | 119.472 | 460 | 20170617092 | 0.617 | 0.018 |
| Fissidentaceae | Fissidens cristatus Wilson ex Mitt. * | Pavilion, Wannnian Temple, Mt. Emei Scenic Spot, Leshan District, Sichuan Province | 29.593 | 103.376 | 810 | 20170810087 | 0.519 | 0.002 |
| Fissidentaceae | Fissidens dubius P. Beauv. | Mt. Moxing, Daishan, Zhoushan District, Zhejiang Province | 30.267 | 122.191 | 113 | 1-20170528023-2 | 0.607 | 0.018 |
| Fissidentaceae | Fissidens grandifrons Brid. * | Caohaizi to Camp 2, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.586 | 102.029 | 2663 | 20170803076 | 0.496 | 0.011 |
| Fissidentaceae | Fissidens teysmannianus Dozy & Molk. * | Mt. Paotai, Daishan, Zhoushan District, Zhejiang Province | 30.281 | 122.302 | 80 | 1-20170530033 | 0.581 | 0.019 |
| Pottiaceae | Barbula rigidula (Hedw.) Mitt. ** | Entrance, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.649 | 102.115 | 1611 | 20170731002 | 0.66 | 0.015 |
| Pottiaceae | Hyophila propagulifera Broth. ** | Mt. Foding, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.012 | 122.384 | 303 | 1-20170424221 | 0.508 | 0.004 |
| Pottiaceae | Hyophila propagulifera Broth. ** | Farm Stay, Zhutuo Hill, Mt. Xitianmu, Linan, Zhejiang Province | 30.328 | 119.454 | 453 | 20170627056 | 0.519 | 0.003 |
| Pottiaceae | Trichostomum hattorianum B.C. Tan & Z. Iwats. * | Tianshi Peak, Mt. Niutou, Wuyi, Zhejiang Province | 28.664 | 119.476 | 730 | 20170617158 | 0.597 | 0.017 |
| Pottiaceae | Weissia longifolia DC. ** | Roadside, Mt. Xianren, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.028 | 121.911 | 299 | 20170221113 | 0.532 | 0.012 |
| Pottiaceae | Weissia platyphylloides Cardot ** | Camp 3 to Dongga Temple, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.568 | 101.993 | 2957 | 20170802201 | 0.492 | 0.002 |
| Hedwigiaceae | Hedwigia ciliata (Hedw.) P. Beauv. | Guanyin Cave, Mt. Putuo, Zhoushan District, Zhejiang Province | 29.985 | 122.373 | 124 | 2-20170426098 | 0.599 | 0.008 |
| Hedwigiaceae | Hedwigia ciliata (Hedw.) P. Beauv. | Entrance, Bingchuanshihe Park, Mt. Haizi, Sichuan Province | 29.296 | 100.082 | 4120 | 20170807001 | 0.48 | 0.004 |
| Bartramiaceae *** | Bartramia ithyphylla Brid. ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.577 | 102.002 | 2983 | 20170802086 | 0.434 | 0.001 |
| Bartramiaceae *** | Philonotis marchica (Hedw.) Brid. ** | Gully, North of Fuyu Mountain Villa, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.448 | 340 | 20170627034 | 0.523 | 0.016 |
| Bartramiaceae *** | Philonotis mollis (Dozy & Molk.) Mitt. ** | Chanyuan Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.442 | 390 | 20170629005 | 0.536 | 0.016 |
| Bartramiaceae *** | Philonotis revoluta Bosch & Sande Lac. ** | Caohaizi to Camp 2, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.586 | 102.029 | 2663 | 20170803083 | 0.506 | 0.003 |
| Bartramiaceae *** | Philonotis revoluta Bosch & Sande Lac. ** | Tianshi Peak, Mt. Niutou, Wuyi, Zhejiang Province | 28.664 | 119.476 | 730 | 20170617160 | 0.576 | 0.01 |
| Bartramiaceae *** | Philonotis thwaitesii Mitt. ** | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.235 | 122.396 | 66 | 2-20170529053 | 0.542 | 0.012 |
| Bryaceae | Brachymenium nepalense Hook. ** | Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.35 | 119.424 | 1484 | 20170630055 | 0.506 | 0.017 |
| Bryaceae | Brachymenium nepalense Hook. ** | Laba River, Mt. Erlang, Sichuan Province | 29.981 | 102.443 | 1228 | 20170808054 | 0.531 | 0.013 |
| Bryaceae | Bryum argenteum Hedw. | Shenniu Valley, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.787 | 70 | 20170617176 | 0.567 | 0.023 |
| Bryaceae | Bryum billarderi Schwägr. * | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.266 | 122.359 | 32 | 2-20170529118 | 0.686 | 0.004 |
| Bryaceae | Bryum billarderii Schwägr. * | Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.428 | 1204 | 20170630026 | 0.596 | 0.015 |
| Bryaceae | Pohlia cruda (Hedw.) Lindb. * | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.552 | 101.97 | 3554 | 20170801087 | 0.471 | 0.005 |
| Bryaceae | Pohlia timmioides Broth. * | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.553 | 101.969 | 3541 | 20170801074 | 0.661 | 0.004 |
| Bryaceae | Rhodobryum giganteum (Schwägr.) Paris * | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.662 | 119.478 | 660 | 20170617140 | 0.862 | 0.006 |
| Mniaceae | Mnium spinosum (Voit) Schwägr. * | Downhill Telpher Station to Heisonglin, Moxi Town, Luding County, Sichuan Province | 29.574 | 101.993 | 3030 | 20170801160 | 0.725 | 0.013 |
| Mniaceae | Plagiomnium acutum (Lindb.) T.J. Kop. * | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170503004 | 0.561 | 0.006 |
| Mniaceae | Plagiomnium acutum (Lindb.) T.J. Kop. * | near Jinhai Heavy Industry, Daishan, Zhejiang Province | 30.244 | 122.263 | 33 | 1-20170530065 | 0.536 | 0.013 |
| Mniaceae | Plagiomnium arbusculum (Müll. Hal.) T.J. Kop. * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.577 | 102.002 | 2983 | 20170802099 | 0.526 | 0.002 |
| Mniaceae | Plagiomnium maximovicgii (Lindb) T. Kop. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 480 | 20170617114 | 0.599 | 0.019 |
| Mniaceae | Plagiomnium maximovicgii (Lindb) T. Kop. * | Gully, North of Fuyu Mountain Villa, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.448 | 340 | 20170629053 | 0.64 | 0.02 |
| Mniaceae | Plagiomnium maximovicgiiv (Lindb) T. Kop. * | Downstairs, Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.446 | 369 | 20170627138 | 0.657 | 0.029 |
| Mniaceae | Rhizomnium gracile T.J. Kop. * | Camp 3 to Dongga Temple, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.568 | 101.993 | 2957 | 20170802195 | 0.558 | 0.012 |
| Mniaceae | Trachycystis microphylla (Dozy & Molk.) Lindb. * | near Jinhai Heavy Industry, Daishan, Zhejiang Province | 30.244 | 122.263 | 33 | 1-20170530062 | 0.619 | 0.01 |
| Mniaceae | Trachycystis microphylla (Dozy & Molk.) Lindb. ** | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170503001 | 0.528 | 0.016 |
| Orthotrichaceae | Macromitrium gymnostomum Sull. & Lesq. ** | Jiufeng Park, Taizhou, Zhejiang Pronvice | 28.906 | 112.464 | 45 | 20160619011 | 0.459 | 0.023 |
| Orthotrichaceae | Macromitrium gymnostomum Sull. & Lesq. ** | Tiesuo Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.664 | 119.478 | 510 | 20170617121 | 0.572 | 0.014 |
| Orthotrichaceae | Macromitrium japonicum Dozy & Molk. ** | Roadside of Fushan Temple, Shengsi, Zhoushan, Zhejiang Province | 30.725 | 122.821 | 20 | 20160405204 | 0.422 | 0.007 |
| Orthotrichaceae | Orthotrichum brassii E.B. Bartram * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.57 | 101.998 | 2930 | 20170802175 | 0.602 | 0.013 |
| Orthotrichaceae | Orthotrichum erubescens Müll. Hal. * | Fayu Temple to Fanyin Cave, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.004 | 122.403 | 40 | 2-20170424092 | 0.504 | 0.007 |
| Orthotrichaceae | Orthotrichum hookeri Wilson ex Mitt. * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.573 | 102.003 | 2927 | 20170802137 | 0.528 | 0.003 |
| Orthotrichaceae | Schlotheimia pungens E.B. Bartram ** | Tianshi Peak, Mt. Niutou, Wuyi, Zhejiang Province | 28.664 | 119.476 | 730 | 20170617159 | 0.558 | 0.025 |
| Orthotrichaceae | Ulota crispa (Hedw.) Brid. ** | Heisonglin, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.574 | 101.991 | 3043 | 20170801164 | 0.524 | 0.005 |
| Aulacomniaceae | Aulacomnium heterostichum (Hedw.) Bruch & Schimp. | Jieyin Temple to Leidongping, Mt. Emei Scenic Spot, Leshan District, Sichuan Province | 29.536 | 103.327 | 2452 | 20170810040 | 0.51 | 0.005 |
| Rhizogoniaceae *** | Pyrrhobryum dozyanum (Sande Lac.) Manuel ** | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 640 | 20170617133 | 0.5 | 0.014 |
| Rhizogoniaceae *** | Pyrrhobryum spiniforme (Hedw.) Mitt. ** | Tianshi Palace, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 480 | 20170617115 | 0.572 | 0.016 |
| Racopilaceae *** | Racopilum cuspidigerum (Schwägr.) Ångstr. ** | Downstairs, Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.446 | 369 | 20170627135 | 0.478 | 0.021 |
| Hypopterygiaceae *** | Hypopterygium japonicum Mitt. ** | Gully, Fuyu Mountain Villa to Zhonglie Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.447 | 385 | 20170629011 | 0.505 | 0.006 |
| Hookeriaceae | Hookeria acutifolia Hook. & Grev. * | Water side, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.088 | 122.509 | 106 | 1-20170216272 | 0.458 | 0.018 |
| Hookeriaceae | Hookeria acutifolia Hook. & Grev. * | near Laodian, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.435 | 1129 | 20170630065 | 0.497 | 0.016 |
| Plagiotheciaceae | Plagiothecium handelii Broth. * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.571 | 101.914 | 2940 | 20170802157 | 0.491 | 0.006 |
| Plagiotheciaceae | Plagiothecium nemorale (Mitt.) A. Jaeger * | Huiji Temple, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.013 | 122.352 | 290 | 1-20170424188 | 0.545 | 0.007 |
| Plagiotheciaceae | Plagiothecium nemorale (Mitt.) A. Jaeger * | Tianshi Palace, Mt. Niutou, Wuyi, Zhejiang Province | 28.67 | 119.472 | 460 | 20170617094 | 0.562 | 0.01 |
| Plagiotheciaceae | Plagiothecium nemorale (Mitt.) A. Jaeger * | Laodian, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.434 | 1099 | 20170710054 | 0.536 | 0.016 |
| Plagiotheciaceae | Plagiothecium piliferum (Sw.) Schimp. * | West of Hong Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.324 | 119.439 | 447 | 20170627020 | 0.581 | 0.012 |
| Climaciaceae | Climacium dendroides (Hedw.) F. Weber & D. Mohr * | Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.433 | 1098 | 20170630006 | 0.571 | 0.01 |
| Climaciaceae | Climacium dendroides (Hedw.) F. Weber & D. Mohr * | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.587 | 101.97 | 3505 | 20170801031 | 0.737 | 0.003 |
| Climaciaceae | Climacium dendroides (Hedw.) F. Weber & D. Mohr * | Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.552 | 101.97 | 3538 | 20170801012 | 0.626 | 0.017 |
| Amblystegiaceae | Calliergon cordifolium (Hedw.) Kindb. * | Gully, Fuyu Mountain Villa to Zhonglie Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.447 | 385 | 20170629048 | 0.56 | 0.016 |
| Amblystegiaceae | Cratoneuron filicinum (Hedw.) Spruce | Kangding, Sichuan Province | 30.009 | 101.952 | 2812 | 20170808039 | 0.463 | 0.003 |
| Amblystegiaceae | Sanionia uncinata (Hedw.) Loeske ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.552 | 101.97 | 3554 | 20170801087 | 0.498 | 0.011 |
| Amblystegiaceae | Sanionia uncinata (Hedw.) Loeske ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.573 | 102.003 | 2462 | 20170802134 | 0.581 | 0.004 |
| Leskeaceae | Lescuraea radicosa (Mitt.) Mönk. ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.577 | 102.001 | 2946 | 20170802016 | 0.788 | 0.011 |
| Thuidiaceae | Actinothuidium hookeri (Mitt.) Broth. ** | Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.574 | 101.993 | 3010 | 20170801143 | 0.502 | 0.003 |
| Thuidiaceae | Actinothuidium hookeri (Mitt.) Broth. ** | Downhill Telpher Station, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.552 | 101.97 | 3557 | 20170801132 | 0.669 | 0.027 |
| Thuidiaceae | Anomodon viticulosus (Hedw.) Hook. & Taylor | Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.445 | 377 | 20170627092 | 0.534 | 0.017 |
| Thuidiaceae | Boulaya mittenii (Broth.) Cardot ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.576 | 102.001 | 2965 | 20170802044 | 0.612 | 0.003 |
| Thuidiaceae | Cyrtohypnum bonianum (Besch.) W.R. Buck & H.A. Crum ** | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.674 | 119.5 | 430 | 20170617026 | 0.671 | 0.01 |
| Thuidiaceae | Cyrtohypnum pygmaeum (Schimp.) Buck et Crum ** | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170509002 | 0.541 | 0.011 |
| Thuidiaceae | Haplocladium angustifolium (Hampe & Müll. Hal.) Broth. * | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170509001 | 0.51 | 0.019 |
| Thuidiaceae | Haplohymenium triste (Ces.) Kindb. * | Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.445 | 377 | 20170629027 | 0.528 | 0.012 |
| Thuidiaceae | Herpetineuron toccoae (Sull. & Lesq.) Cardot ** | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.261 | 122.341 | 117 | 1-20170530013 | 0.619 | 0.025 |
| Thuidiaceae | Herpetineuron toccoae (Sull. & Lesq.) Cardot ** | Pavilion, Wannnian Temple, Mt. Emei Scenic Spot, Leshan District, Sichuan Province | 29.643 | 103.476 | 816 | 20170810082 | 0.571 | 0.003 |
| Thuidiaceae | Pinnatella anacamptolepis (Müll. Hal.) Broth. ** | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.668 | 119.472 | 470 | 20170617097 | 0.545 | 0.014 |
| Thuidiaceae | Thuidium cymbifolium (Dozy & Molk.) Dozy & Molk. * | Entrance, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.649 | 102.115 | 1611 | 20170731018 | 0.599 | 0.015 |
| Thuidiaceae | Thuidium kanedae Sakurai * | Roadside, Taizi Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.324 | 119.439 | 447 | 20170627115 | 0.659 | 0.021 |
| Brachytheciaceae | Brachythecium buchananii (Hook.) A. Jaeger * | Entrance, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.649 | 102.115 | 1611 | 20170731013 | 0.554 | 0.002 |
| Brachytheciaceae | Brachythecium piligerum Cardot * | Jieyin Temple to Leidongping, Mt. Emei Scenic Spot, Leshan District, Sichuan Province | 29.502 | 103.331 | 2091 | 20170810021 | 0.547 | 0.005 |
| Brachytheciaceae | Brachythecium plumosum (Hedw.) Schimp. * | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170517001 | 0.492 | 0.013 |
| Brachytheciaceae | Cirriphyllum cirrosum (Schwägr.) Grout * | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.577 | 102.001 | 2946 | 20170802014 | 0.606 | 0.003 |
| Brachytheciaceae | Eurhynchium hians (Hedw.) Sande Lac. | Lianfeng Road, Mt. Xitianmu, Linan, Zhejiang Province | 30.341 | 119.436 | 1101 | 20170710117 | 0.498 | 0.008 |
| Brachytheciaceae | Eurhynchium savatieri Schimp. ex Besch. * | Mt. Moxing, Daishan, Zhoushan District, Zhejiang Province | 30.264 | 122.192 | 180 | 1-20170528006-2 | 0.589 | 0.019 |
| Brachytheciaceae | Myuroclada maximowiczii (G.G. Borshch.) Steere & W.B. Schofield ** | Mt. Moxing, Daishan, Zhoushan District, Zhejiang Province | 30.269 | 122.36 | 68 | 1-20170528032-3 | 0.498 | 0.015 |
| Brachytheciaceae | Palamocladium leskeoides (Hook.) E. Britton ** | Jinshanmen, Mt. Xitianmu, Linan, Zhejiang Province | 30.326 | 119.441 | 341 | 20170629012 | 0.54 | 0.008 |
| Brachytheciaceae | Rhynchostegium pallidifolium (Mitt.) A. Jaeger * | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.239 | 122.397 | 13 | 2-20170529046 | 0.579 | 0.025 |
| Brachytheciaceae | Rhynchostegium riparioides (Hedw.) Cardot * | Mt. Moxing, Daishan, Zhoushan District, Zhejiang Province | 30.266 | 122.191 | 143 | 1-20170528019-2 | 0.564 | 0.017 |
| Hypnaceae | Homomallium connexum (Cardot) Broth. * | Telpher Station, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.026 | 122.397 | 71 | 1-20170425044 | 0.458 | 0.016 |
| Hypnaceae | Homomallium connexum (Cardot) Broth. * | Farm Stay, Zhutuo Hill, Mt. Xitianmu, Linan, Zhejiang Province | 30.328 | 119.454 | 453 | 20170627052 | 0.507 | 0.017 |
| Hypnaceae | Homomallium connexum (Cardot) Broth. * | Mali Tree, Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.445 | 377 | 20170629045 | 0.572 | 0.027 |
| Hypnaceae | Hypnum cupressiforme Hedw. * | Dachangtu, Daishan, Zhoushan District, Zhejiang Province | 30.235 | 122.396 | 66 | 2-20170529049 | 0.465 | 0.011 |
| Hypnaceae | Hypnum plumaeforme Wilson * | Botanical Garden, Xuhui District, Shanghai Normal University | 31.164 | 121.413 | 20 | 20170509003 | 0.48 | 0.022 |
| Hypnaceae | Hypnum revolutum (Mitt.) Lindb. * | Entrance, Bingchuanshihe Park, Mt. Haizi, Sichuan Province | 29.298 | 100.082 | 4140 | 20170807012 | 0.593 | 0.013 |
| Hypnaceae | Pseudotaxiphyllum pohliaecarpum (Sull. & Lesq.) Z. Iwats. ** | Mt. Mahuang, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.062 | 122.052 | 109 | 2-20170430033 | 0.617 | 0.003 |
| Hypnaceae | Pseudotaxiphyllum pohliaecarpum (Sull. & Lesq.) Z. Iwats. ** | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.671 | 119.484 | 460 | 20170617050 | 0.578 | 0.021 |
| Hypnaceae | Ptilium crista-castrensis (Hedw.) De Not. ** | Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.651 | 102.115 | 1594 | 20170801007 | 0.571 | 0.025 |
| Hypnaceae | Ptilium crista-castrensis (Hedw.) De Not. ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.553 | 101.969 | 3541 | 20170801067 | 0.659 | 0.011 |
| Hypnaceae | Taxiphyllum taxirameum (Mitt.) M. Fleisch. * | Tianshi Palace, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 480 | 20170617116 | 0.536 | 0.024 |
| Meteoriaceae | Aerobryidium filamentosum (Hook.) M. Fleisch. ** | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.673 | 119.485 | 420 | 20170617075 | 0.589 | 0.018 |
| Meteoriaceae | Chrysocladium retrorsum (Mitt.) M. Fleisch. ** | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.668 | 119.481 | 480 | 20170617107 | 0.642 | 0.003 |
| Meteoriaceae | Meteorium subpolytrichum (Besch.) Broth. * | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.663 | 119.312 | 670 | 20170617161 | 0.535 | 0.008 |
| Meteoriaceae | Neodicladiella pendula (Sull.) W.R. Buck ** | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 490 | 20170617112 | 0.598 | 0.023 |
| Meteoriaceae | Neodicladiella pendula (Sull.) W.R. Buck ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi, Luding County, Sichuan Province | 29.576 | 102.072 | 2965 | 20170802030 | 0.654 | 0.01 |
| Meteoriaceae | Neodicladiella pendula (Sull.) W.R. Buck ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi, Luding County, Sichuan Province | 29.573 | 102.003 | 2462 | 20170802118 | 0.576 | 0.018 |
| Sematophyllaceae | Barbella spiculata (Mitt.) Broth. ** | Downhill Telpher Station to Heisonglin, Hailuo Gully, Moxi, Luding County, Sichuan Province | 29.575 | 101.993 | 2997 | 20170801152 | 0.482 | 0.001 |
| Sematophyllaceae | Brotherella falcata (Dozy & Molk.) M. Fleisch. ** | Downhill Telpher Station, Hailuo Gully, Moxi, Luding County, Sichuan Province | 29.574 | 101.993 | 3010 | 20170801137 | 0.565 | 0.005 |
| Sematophyllaceae | Sematophyllum subhumile (Müll. Hal.) M. Fleisch. ** | Mt. Moxing, Daishan, Zhoushan District, Zhejiang Province | 30.266 | 122.191 | 120 | 2-20170726015 | 0.557 | 0.01 |
| Sematophyllaceae | Trichosteleum lutschianum (Broth. & Paris) Broth. ** | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 630 | 20170617130 | 0.498 | 0.013 |
| Trachypodaceae *** | Trachypus bicolor Reinw. & Hornsch. * | Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.428 | 1204 | 20170630039 | 0.571 | 0.007 |
| Hylocomiaceae | Hylocomiastrum himalayanum (Mitt.) Broth. ** | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.553 | 101.969 | 3541 | 20170801064 | 0.578 | 0.008 |
| Hylocomiaceae | Hylocomium splendens (Hedw.) Schimp. | Downhill Telpher Station, Hailuo Gully, Moxi, Luding County, Sichuan Province | 29.552 | 101.97 | 3557 | 20170801133 | 0.548 | 0.012 |
| Hylocomiaceae | Hylocomium splendens (Hedw.) Schimp. | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi, Luding County, Sichuan Province | 29.571 | 101.914 | 2940 | 20170802160 | 0.644 | 0.01 |
| Rhytidiaceae | Rhytidiadelphus triquetrus (Hedw.) Warnst. | Glaciers 1, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.552 | 101.97 | 3557 | 20170801091 | 0.81 | 0.022 |
| Rhytidiaceae | Rhytidiadelphus triquetrus (Hedw.) Warnst. | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.57 | 101.998 | 2930 | 20170802176 | 0.749 | 0.011 |
| Rhytidiaceae | Rhytidium rugosum (Hedw.) Kindb. | Entrance, Bingchuanshihe Park, Mt. Haizi, Sichuan Province | 29.298 | 100.082 | 4140 | 20170807009 | 0.509 | 0.012 |
| Rhytidiaceae | Rhytidium rugosum (Hedw.) Kindb. | Foot of Mt. Bowa, Daocheng, Sichuan Province | 29.593 | 100.289 | 4080 | 20170805017 | 0.563 | 0.016 |
| Entodontaceae | Entodon concinnus (De Not.) Paris * | Shenniu Valley, Mt. Niutou, Wuyi, Zhejiang Province | 28.956 | 119.794 | 70 | 20170617173 | 0.496 | 0.023 |
| Entodontaceae | Entodon divergens Broth. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.668 | 119.481 | 480 | 20170617106 | 0.544 | 0.012 |
| Entodontaceae | Entodon macropodus (Hedw.) Müll. Hal. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.671 | 119.484 | 460 | 20170617055 | 0.522 | 0.005 |
| Entodontaceae | Entodon prorepens (Mitt.) A. Jaeger * | Entrance, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.65 | 102.115 | 1611 | 20170731023 | 0.459 | 0.006 |
| Entodontaceae | Entodon sullivantii (Müll. Hal.) Lindb. * | Yinxiu Nunnery, Mt. Putuo, Zhoushan District, Zhejiang Province | 29.978 | 122.377 | 37 | 2-20170426064 | 0.528 | 0.01 |
| Entodontaceae | Pleurozium schreberi (Brid.) Mitt. | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 650 | 20170617135 | 0.549 | 0.008 |
| Entodontaceae | Pleurozium schreberi (Brid.) Mitt. | Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.35 | 119.424 | 1484 | 20170630049 | 0.625 | 0.012 |
| Entodontaceae | Pleurozium schreberi (Brid.) Mitt. | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.571 | 101.914 | 2940 | 20170802161 | 0.469 | 0.008 |
| Leucodontaceae | Dozya japonica Sande Lac. ** | Farm Stay, Linzhongyuan, Mt. Xitianmu, Linan, Zhejiang Province | 30.328 | 119.454 | 452 | 20170627063 | 0.567 | 0.025 |
| Leucodontaceae | Leucodon secundus (Harv.) Mitt. ** | Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.575 | 101.993 | 2997 | 20170801150 | 0.516 | 0.015 |
| Leucodontaceae | Leucodon secundus (Harv.) Mitt. ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.571 | 101.914 | 2940 | 20170802155 | 0.496 | 0.002 |
| Leucodontaceae | Leucodon subulatulus Broth. ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.574 | 101.992 | 3020 | 20170802003 | 0.548 | 0.01 |
| Cryphaeaceae *** | Pilotrichopsis robusta P.C. Chen ** | Camp 3 to Chengmen Cave, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.571 | 101.914 | 2940 | 20170802163 | 0.472 | 0.011 |
| Neckeraceae | Homalia trichomanoides (Hedw.) Schimp. * | Gully, Fuyu Mountain Villa to Zhonglie Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.447 | 385 | 20170629054 | 0.522 | 0.013 |
| Neckeraceae | Homalia trichomanoides (Hedw.) Schimp. * | Mt. Wutong Scenic Spot, Shenzhen | 22.564 | 114.222 | 160 | 20170726003 | 0.716 | 0.007 |
| Neckeraceae | Homaliodendron flabellatum (Sm.) M. Fleisch. * | Tianshi Palace, Mt. Niutou, Wuyi, Zhejiang Province | 28.668 | 119.481 | 480 | 20170617104 | 0.542 | 0.008 |
| Neckeraceae | Homaliodendron ligulaefolium (Mitt.) M. Fleisch. * | Laodian to Xianrending, Mt. Xitianmu, Linan, Zhejiang Province | 30.342 | 119.428 | 1204 | 20170630011 | 0.601 | 0.017 |
| Neckeraceae | Homaliodendron scalpellifolium (Mitt.) M. Fleisch. * | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 650 | 20170617136 | 0.558 | 0.013 |
| Neckeraceae | Homaliodendron scalpellifolium (Mitt.) M. Fleisch. * | Camp 3 to Dongga Temple, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.568 | 101.993 | 2957 | 20170802202 | 0.492 | 0.013 |
| Neckeraceae | Neckera decurrens Broth. * | Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.651 | 102.115 | 1594 | 20170801004 | 0.493 | 0.016 |
| Neckeraceae | Neckera humilis Mitt. * | Huiji Temple, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.013 | 122.385 | 236 | 1-20170424174 | 0.51 | 0.007 |
| Neckeraceae | Neckera humilis Mitt. * | Huiji Temple, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.013 | 122.385 | 236 | 1-20170424174 | 0.587 | 0.003 |
| Neckeraceae | Neckera yezoana Besch. * | Pavilion, Wannnian Temple, Mt. Emei Scenic Spot, Leshan District, Sichuan Province | 29.593 | 103.376 | 810 | 20170810092 | 0.595 | 0.006 |
| Neckeraceae | Thamnobryum subseriatum (Mitt. ex Sande Lac.) B.C. Tan * | Huiji Temple, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.013 | 122.352 | 290 | 1-20170424184 | 0.587 | 0.013 |
| Neckeraceae | Thamnobryum subseriatum (Mitt. ex Sande Lac.) B.C. Tan * | Dashuwang, Mt. Xitianmu, Linan, Zhejiang Province | 30.341 | 119.433 | 1075 | 20170710065 | 0.532 | 0.038 |
| Neckeraceae | Thamnobryum subseriatum (Sw.) Schimp. * | Gully, Left Hand of Stele, Fuyu Mountain Villa, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.448 | 340 | 20170629056 | 0.497 | 0.007 |
| Marchantiaceae | Marchantia polymorpha L. | Shenniu Valley, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.787 | 70 | 20170617181 | 0.518 | 0.018 |
| Marchantiaceae | Marchantia polymorpha L. | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 480 | 20170617118 | 0.552 | 0.027 |
| Dumortieraceae *** | Dumortiera hirsuta (Sw.) Nees ** | Tiesuo Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 660 | 20170617138 | 0.552 | 0.03 |
| Dumortieraceae *** | Dumortiera hirsuta (Sw.) Nees ** | Roadside, Fuyu Mountain Villa to Zhonglie Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.447 | 376 | 20170627007 | 0.621 | 0.019 |
| Pallaviciniaceae | Pallavicinia ambigua (Mitt.) Steph. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.67 | 119.481 | 410 | 20170617088 | 0.52 | 0.005 |
| Pallaviciniaceae | Pallavicinia ambigua (Mitt.) Steph. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.668 | 119.481 | 480 | 20170617102 | 0.534 | 0.005 |
| Cephaloziaceae | Alobiellopsis parvifolius (Steph.) R. M. Schust. ** | Water side, Mt. Putuo, Zhoushan District, Zhejiang Province | 30.028 | 121.906 | 229 | 20170221042 | 0.471 | 0.015 |
| Scapaniaceae | Scapania verrucosa Heeg * | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.671 | 119.484 | 460 | 20170617048 | 0.496 | 0.004 |
| Trichocoleaceae | Trichocolea tomentella (Ehrh.) Dumort. | Camp 3 to Dongga Temple, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.568 | 101.993 | 2957 | 20170802193 | 0.653 | 0.004 |
| Lepidoziaceae | Bazzania denudata (Torr. ex Gottsche, Lindenb. & Nees) Trevis. | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.673 | 119.485 | 410 | 20170617007 | 0.641 | 0.028 |
| Lepidoziaceae | Bazzania praerupta (Reinw.Blume & Nees) Trevis. * | Xiagushanmen, Mt. Niutou, Wuyi, Zhejiang Province | 28.674 | 119.5 | 430 | 20170617018 | 0.551 | 0.006 |
| Lepidoziaceae | Lepidozia subintegra Lindenb. * | Tiesuo Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 640 | 20170617132 | 0.514 | 0.017 |
| Plagiochilaceae | Plagiochila chinensis Steph. * | Camp 3 to Dongga Temple, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.568 | 101.993 | 2957 | 20170802191 | 0.673 | 0.007 |
| Plagiochilaceae | Plagiochila durelii Schiffn. * | Downstairs, Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.322 | 119.446 | 369 | 20170629036 | 0.542 | 0.018 |
| Plagiochilaceae | Plagiochila griffithiana Steph. * | Yixiantian, Mt. Niutou, Wuyi, Zhejiang Province | 28.666 | 119.476 | 640 | 20170617131 | 0.614 | 0.003 |
| Plagiochilaceae | Plagiochila zonata Steph. * | Camp 3 to Dongga Temple, Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.568 | 101.993 | 2957 | 20170802190 | 0.53 | 0.012 |
| Porellaceae | Porella caespitans (Steph.) S. Hatt. * | Hailuo Gully, Moxi Town, Luding County, Sichuan Province | 29.575 | 101.993 | 2997 | 20170801154 | 0.612 | 0.007 |
| Porellaceae | Porella chinensis (Steph.) Hatt. * | Liuchun House, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.445 | 376 | 20170627130 | 0.613 | 0.013 |
| Frullaniaceae | Frullania ericoides (Nees) Mont. * | Lingshi Temple, Mt. Putuo, Zhoushan District, Zhejiang Province | 29.986 | 122.372 | 121 | 3-20170425108 | 0.531 | 0.005 |
| Lejeuneaceae | Trocholejeunea sandvicensis (Gottsche) Mizut. ** | Fuyu Mountain Villa to Zhonglie Temple, Mt. Xitianmu, Linan, Zhejiang Province | 30.323 | 119.447 | 376 | 20170627045 | 0.659 | 0.021 |
| Metzgeriaceae | Metzgeria novicrassipilis Kuwah. * | Qixi Bridge, Mt. Niutou, Wuyi, Zhejiang Province | 28.67 | 119.482 | 440 | 20170617080 | 0.664 | 0.004 |
References
- Greilhuber, J.; Dolezel, J.; Lysak, M.; Bennett, M.D. The origin, evolution and proposed stabilization of the terms ‘genome size’and ‘C-value’ to describe nuclear DNA contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Leitch, I.J.; Johnston, E.; Pellicer, J.; Hidalgo, O.; Bennett, M.D. Plant DNA C-values database (release 4.0, April 2019). 2019. Available online: https://cvalues.science.kew.org/ (accessed on 20 December 2022).
- Bennett, M.D.; Leitch, I.J. Nuclear DNA amounts in angiosperms. Ann. Bot. 1995, 76, 113–176. [Google Scholar] [CrossRef]
- Bennett, M.D.; Leitch, I.J. Nuclear DNA amounts in angiosperms: Progress, problems and prospects. Ann. Bot. 2005, 95, 45–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, M.D.; Smith, J.B. Nuclear DNA amounts in angiosperms. Philos. Trans. R. Soc. B 1976, 274, 227–274. [Google Scholar] [CrossRef]
- Knight, C.A.; Molinari, N.; Petrov, D.A. The large genome constraint hypothesis: Evolution, ecology and phenotype. Ann. Bot. 2005, 95, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Leitch, I.J.; Soltis, D.E.; Soltis, P.S.; Bennett, M.D. Evolution of DNA amounts across land plants (Embryophyta). Ann. Bot. 2005, 95, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Price, H.J.; Dillon, S.L.; Hodnett, G.; Rooney, W.L.; Ross, L.; Johnston, J.S. Genome evolution in the genus Sorghum (Poaceae). Ann. Bot. 2005, 95, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Li, D.D.; Lou, Y.X.; Guo, S.L. Nuclear DNA content variation of herbaceous angiosperm species on 10 global latitudinal transects. J. Torrey Bot. Soc. 2018, 145, 341–352. [Google Scholar] [CrossRef]
- Bainard, J.D.; Forrest, L.L.; Goffinet, B.; Newmaster, S.G. Nuclear DNA content variation and evolution in liverworts. Mol. Phylogenet. Evol. 2013, 68, 619–627. [Google Scholar] [CrossRef]
- Bainard, J.D.; Newmaster, S.G.; Budke, J.M. Genome size and endopolyploidy evolution across the moss phylogeny. Ann. Bot. 2020, 125, 543–555. [Google Scholar] [CrossRef]
- Chen, G.Q.; Guo, S.L.; Yin, L.P. Applying DNA C-values to evaluate invasiveness of angiosperms: Validity and limitation. Biol. Invasions 2010, 12, 1335–1348. [Google Scholar] [CrossRef]
- Voglmayr, H. Nuclear DNA amounts in mosses (Musci). Ann. Bot. 2000, 85, 531–546. [Google Scholar] [CrossRef] [Green Version]
- Temsch, E.M.; Greilhuber, J.; Krisai, R. Genome size in liverworts. Preslia 2010, 82, 63–80. [Google Scholar]
- Bainard, J.D. Patterns and biological implications of DNA content variation in land plants. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2011. Chapter 2. [Google Scholar]
- Bainard, J.D.; Villarreal, J.C. Genome size increase in recently diverged hornwort clades. Genome 2013, 56, 431–435. [Google Scholar] [CrossRef]
- Greilhuber, J.; Sastad, S.M.; Flatberg, K.I. Ploidy determination in Sphagnum samples from Svalbard, Arctic Norway, by DNA image cytometry. J. Bryol. 2003, 25, 235–239. [Google Scholar] [CrossRef]
- Goffinet, B.; Shaw, A.J. Bryophyte Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Aranda, S.C.; Gabriel, R.; Borges, P.A.V.; Santos, A.M.C.; Azevedo, E.B.; Patiño, J.; Hortal, J.; Lobo, J.M. Geographical, temporal and environmental determinants of bryophyte species richness in the Macaronesian Islands. PLoS ONE 2014, 9, e101786. [Google Scholar] [CrossRef]
- Pharo, E.J.; Zartman, C.E. Bryophytes in a changing landscape: The hierarchical effects of habitat fragmentation on ecological and evolutionary processes. Biol. Conserv. 2007, 135, 315–325. [Google Scholar] [CrossRef]
- He, X.L.; He, K.S.; Hyvönen, J. Will bryophytes survive in a warming world? Perspect. Plant Ecol. 2016, 19, 49–60. [Google Scholar] [CrossRef]
- Narayan, R.K.J. Constraints upon the organization and evolution of chromosomes in Allium. Theor. Appl. Genet. 1988, 75, 319–329. [Google Scholar] [CrossRef]
- Cavallini, A.; Natali, L. Intraspecific variation of nuclear DNA content in plant species. Caryologia 1991, 44, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Furnta, Y.; Nishikawa, K. Variation in nuclear and individual chromosomal DNA content and its role in evolution of plants. In Chromosome Engineering in Plants: Genetics, Breeding, Evolution. Part A; Gupta, P.K., Tsuchiya, T., Eds.; Elsevier: New York, NY, USA, 1991; pp. 71–85. [Google Scholar]
- Bennett, M.D.; Leitch, I.J. Nuclear DNA amounts in angiosperms: Targets, trends and tomorrow. Ann. Bot. 2011, 107, 467–590. [Google Scholar] [CrossRef] [PubMed]
- Baniaga, A.E.; Arrigo, N.; Barker, M.S. The small nuclear genomes of Selaginella are associated with a low rate of genome size evolution. Genome Biol. Evol. 2016, 8, 1516–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temsch, E.M.; Greilhuber, J.; Krisai, R. Genome size in Sphagnum (peat moss). Plant Biol. 1998, 111, 325–330. [Google Scholar] [CrossRef]
- Renzaglia, K.S.; Rasch, E.M.; Pike, L.M. Estimates of nuclear DNAcontent in bryophyte sperm cells: Phylogenetic considerations. Am. J. Bot. 1995, 82, 18–25. [Google Scholar] [CrossRef]
- Lawrence, M.E.; Senecio, L. (Asteraceae) in Australia: Nuclear DNA amounts. Aust. J. Bot. 1985, 33, 221–232. [Google Scholar] [CrossRef]
- Bennett, M.D. Variation in genomic form in plants and its ecological implications. New Phytol. 1987, 106, 177–200. [Google Scholar] [CrossRef]
- Bennett, M.D.; Heslopharrison, J.S.; Smith, J.B.; Ward, J.P. DNA density in mitotic and meiotic metaphase chromosomes of plants and animals. J. Cell Sci. 1983, 63, 173–179. [Google Scholar] [CrossRef]
- Anderson, L.K.; Stack, S.M.; Fox, M.H.; Zhang, C.S. The relation between genome size and synaptonemal complex length in higher plants. Exp. Cell Res. 1985, 156, 367–377. [Google Scholar] [CrossRef]
- Rees, H.; Cameron, F.M.; Hazarika, M.H.; Jones, G.H. Nuclear Variation between Diploid Angiosperms. Nature 1966, 211, 828–830. [Google Scholar] [CrossRef]
- Edwards, G.A.; Endrizzi, J.E. Cell size, nuclear size and DNA content relationships in Gossypium. Can. J. Genet. Cytol. 1975, 17, 181–186. [Google Scholar] [CrossRef]
- Sugiyama, S.; Yamaguchi, K.; Yamada, T. Intraspecific phenotypic variation associated with nuclear DNA content in Lolium perenne L. Euphytica 2002, 128, 145–151. [Google Scholar] [CrossRef]
- Zonneveld, B.J.M.; Duncan, G.D. Genome size for the species of Nerine Herb. (Amaryllidaceae) and its evident correlation with growth cycle, leaf width and other morphological characters. Plant Syst. Evol. 2005, 257, 251–260. [Google Scholar] [CrossRef]
- Evans, G.M.; Rees, H.; Snell, C.L.; Sun, S. The relationship between nuclear DNA amount and the duration of the mitotic cycle. In Chromosomes Today; Darlington, C.D., Lewis, K.R., Eds.; Hafner: New York, NY, USA, 1972; pp. 24–31. [Google Scholar]
- Bennett, M.D. The time and duration of meiosis. Philos. Trans. R. Soc. B 1977, 277, 201–226. [Google Scholar] [CrossRef]
- Bennett, M.D. Nuclear DNA content and minimum generation time in herbaceous plants. Proc. R. Soc. B 1972, 181, 109–135. [Google Scholar] [CrossRef]
- Cutler, J.M.; Rains, D.M.; Loomis, R.S. The importance of cell size in the water relations of plants. Physiol. Plantarum 1977, 40, 255–260. [Google Scholar] [CrossRef]
- Rejmánek, M. A theory of seed plant invasiveness: The first sketch. Biol. Conserv. 1996, 78, 171–181. [Google Scholar] [CrossRef]
- Bartels, S.F.; James, R.S.; Cancers, R.T.; Macdonald, S.E. Depth-to-water mediates bryophyte response to harvesting in boreal forests. J. Appl. Ecol. 2019, 56, 1256–1266. [Google Scholar] [CrossRef]
- Vekemans, X.; Lefèbvre, C.; Coulaud, J.; Blaise, S.; Gruber, W.; Siljak-Yakovlev, S.; Brown, S.C. Variation in Nuclear DNA Content at the Species Level in Armeria Maritima. Hereditas 1996, 124, 237–242. [Google Scholar] [CrossRef]
- Löbel, S.; Rydin, H. Trade-offs and habitat constraints in the establishment of epiphytic bryophytes. Funct. Ecol. 2010, 24, 887–897. [Google Scholar] [CrossRef]
- Proctor, M.C.F.; Nagy, Z.; Csintalan, Z.; Takacs, Z. Water-content components in bryophytes: Analysis of pressure-volume relationships. J. Exp. Bot. 1998, 49, 1845–1954. [Google Scholar] [CrossRef]
- Price, H.J. DNA content variation among higher plants. Ann. MO. Bot. Gard. 1988, 75, 1248–1257. [Google Scholar] [CrossRef]
- Leitch, I.J.; Chase, M.W.; Bennett, M.D. Phylogenetic analysis of DNA C-values provides evidence for a small ancestral genome size in flowering plants. Ann. Bot. 1998, 82 (Suppl. A), 85–94. [Google Scholar] [CrossRef]
- Soltis, D.E.; Soltis, P.S.; Bennett, M.D.; Leitch, I.J. Evolution of genome size in the angiosperms. Am. J. Bot. 2003, 90, 1596–1603. [Google Scholar] [CrossRef] [PubMed]
- Ohri, D.; Fritsch, R.M.; Hanelt, P. Evolution of genome size in Allium (Alliaceae). Plant Sys. Evol. 1998, 210, 57–86. [Google Scholar] [CrossRef]
- Moscone, E.A.; Baranyi, M.; Ebnert, I.; Greilhuber, J.; Ehrendorfer, F.; Hunziker, A.T. Analysis of nuclear DNA content in Capsicum (Solanaceae) by flow cytometry and feulgen densiometry. Ann. Bot. 2003, 91, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Caetano-Anollés, G. Evolution of genome size in the Grasses. Crop Sci. 2005, 45, 1809–1816. [Google Scholar] [CrossRef]
- Cruz, G.A.; Filho, J.R.; Vasconcelos, S.; Gitai, J.; De Campos, J.M.; Viccini, L.F.; Zizka, G.; Leme, E.M.; Brasilerio-vidal, A.C.; Benko-iseppon, A.M. Genome size evolution and chromosome numbers of species of the cryptanthoid complex (Bromelioideae, Bromeliaceae) in a phylogenetic framework. Bot. J. Linn. Soc. 2020, 192, 887–899. [Google Scholar] [CrossRef]
- Jia, Y.; He, S. Species Catalogue of China: Bryophytes; Science Press: Beijing, China, 2013. [Google Scholar]
- Johnston, J.S.; Bennett, M.D.; Rayburn, A.L.; Galbraith, D.W.; Price, H.J. Reference standards for determination of DNA content of plant nuclei. Am. J. Bot. 1999, 86, 609–613. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef]
- Bainard, J.D.; Fazekas, A.J.; Newmaster, S.G. Methodology significantly affects genome size estimates: Quantitative evidence using bryophytes. Cytometry Part A 2010, 77, 725–732. [Google Scholar] [CrossRef]
- Schween, G.; Gorr, G.; Hohe, A.; Reski, R. Unique tissue-specific cell cycle in Physcomitrella. Plant Biol. 2003, 5, 50–58. [Google Scholar] [CrossRef]
- Rensing, S.A.; Goffinet, B.; Meyberg, R.; Wu, S.Z.; Bezanilla, M. The Moss Physcomitrella patens: A Model Organism for Non-Seed Plants. Plant Cell 2020, 32, 1361–1367. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Q.; Kerstetter, R.A.; Michael, T.P. Evolution of genome size in duckweeds (Lemnaceae). J. Bot. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y. Measuring genome size in silver moss Bryum argenteum Hedw. by Flow Cytometry. Mol. Plant Breed. 2016, 14, 858–863. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, X.Y.; Xue, Q.Z. Comparison on the effects of different isolaton buffer on the DNA resolution of the cell nucleus suspensions from rice. Acta Agric. Zhejiangensis 2007, 19, 93–96. [Google Scholar] [CrossRef]
- Du, L.Y.; Feng, R.Q. Flow Ctyometer; Peking University Press: Bejing, China, 2008. [Google Scholar]
- Bainard, J.D.; Newmaster, S.G. Endopolyploidy in bryophytes: Widespread in mosses and absent in liverworts. J. Bot. 2010, 2010, 316356. [Google Scholar] [CrossRef] [Green Version]
- Murray, B.G.; Veir, I.E.; Ferguson, A.R.; Lange, P.J. Variation in DNA C-value and haploid genome size in New Zealand native grasses. New Zealand J. Bot. 2003, 41, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Pasqual, M.; Bruzi, A.; Pio, L.A.S.; Mendonça, P.M.S.; Soares, J.D.R. Flow cytometry reliability analysis and variations in sugarcane DNA content. Genet. Mol. Res. 2015, 14, 7172–7183. [Google Scholar] [CrossRef]
- Kong, L.Y.; Li, D.M.; Liu, Y.L. Ploidy identification of six Grape Hyacinths. J. Northwest For. Univ. 2018, 32, 100–103. [Google Scholar]
- Tyagi, A.; Sandhya, S.; Sharma, P.; Saxena, S.; Sharma, R.; Mithra, S.V.; Mithra, A.; Solanke, A.U.; Singh, N.K.; Sharma, T.R.; et al. The genome size of clusterbean (Cyamopsis tetragonoloba) is significantly smaller compared to its wild relatives as estimated by flow cytometry. Gene 2019, 707, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.W.; Liu, L.Q.; Pamier, A.N.; Wang, Y.N.; Cheng, G.; Liao, K. Identification of chromosomal ploidy and DNA content in Xinjiang Armeniaca vulgaris by flow cytometry. J. Agric. Biotechnol. 2019, 27, 542–550. [Google Scholar] [CrossRef]
- Yumni, R.M.; Karim, M.F.; Midin, M.R. Genome size determination of cucumber (Cucumis sativus), honeydew (Cucumis melo inodorus) and rock melon (Cucumis melo cantalupensis) via flow cytometry. Sci. Heritage J. 2021, 5, 14–16. [Google Scholar] [CrossRef]
- Al-Qurainy, F.; Tarroum, M.; Khan, S.; Nadeem, M.; Gaafar, A.R.Z.; Alansi, S.; Alfarraj, N.S. Genome estimation and phytochemical compound identification in the leaves and callus of Abrus precatorius: A Locally Endangered Plant from the Flora of Saudi Arabia. Plants 2022, 11, 567. [Google Scholar] [CrossRef]
- Müller, K.; Quandt, D.; Müller, J.; Neinhuis, C. PhyDE: Phylogenetic Data Editor, version 0.9971. 2010. Available online: https://www.phyde.de/ (accessed on 20 December 2022).
- Nylander, J.A.A. MrModeltest 2.3. Program distributed by the author. Evolutionary Biology Centre, Uppsala University. 2009. Available online: http://www.abc.se/~nylander/ (accessed on 20 December 2022).
- Swofford, D.L. PAUP*: Phylogenetic analysis using parsi- mony, version 4.0 b10. Sunderland (MA): Sinauer. 2003. Available online: http://paup.phylosolutions.com/ (accessed on 20 December 2022).
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Stöver, B.C.; Müller, K.F. Treegraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Kembel, S.; Cowan, P.; Helmus, M.R. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Luo, G.; Guo, S.; Huang, R.; Yang, J.; Cao, T.; Yu, J. Nuclear DNA Amounts in Chinese Bryophytes Estimated by Flow Cytometry: Variation Patterns and Biological Significances. Plants 2023, 12, 1564. https://doi.org/10.3390/plants12071564
Li D, Luo G, Guo S, Huang R, Yang J, Cao T, Yu J. Nuclear DNA Amounts in Chinese Bryophytes Estimated by Flow Cytometry: Variation Patterns and Biological Significances. Plants. 2023; 12(7):1564. https://doi.org/10.3390/plants12071564
Chicago/Turabian StyleLi, Dandan, Guangyu Luo, Shuiliang Guo, Ruoling Huang, Jun Yang, Tong Cao, and Jing Yu. 2023. "Nuclear DNA Amounts in Chinese Bryophytes Estimated by Flow Cytometry: Variation Patterns and Biological Significances" Plants 12, no. 7: 1564. https://doi.org/10.3390/plants12071564