
Spatial Distribution Characteristics of Suitable Planting Areas for Pyrus Species under Climate Change in China
Abstract
:1. Introduction
2. Results
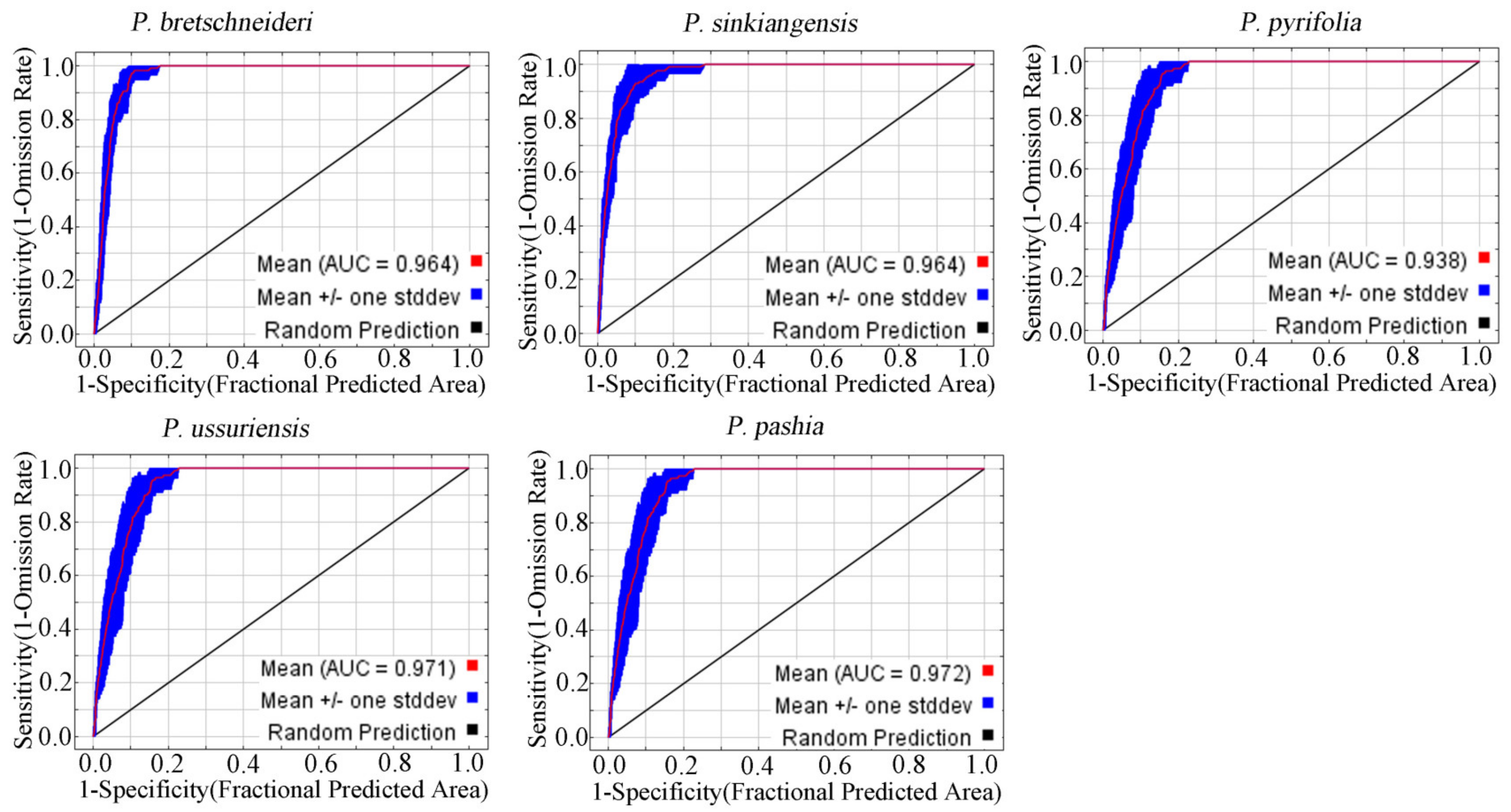
2.1. Model Verification
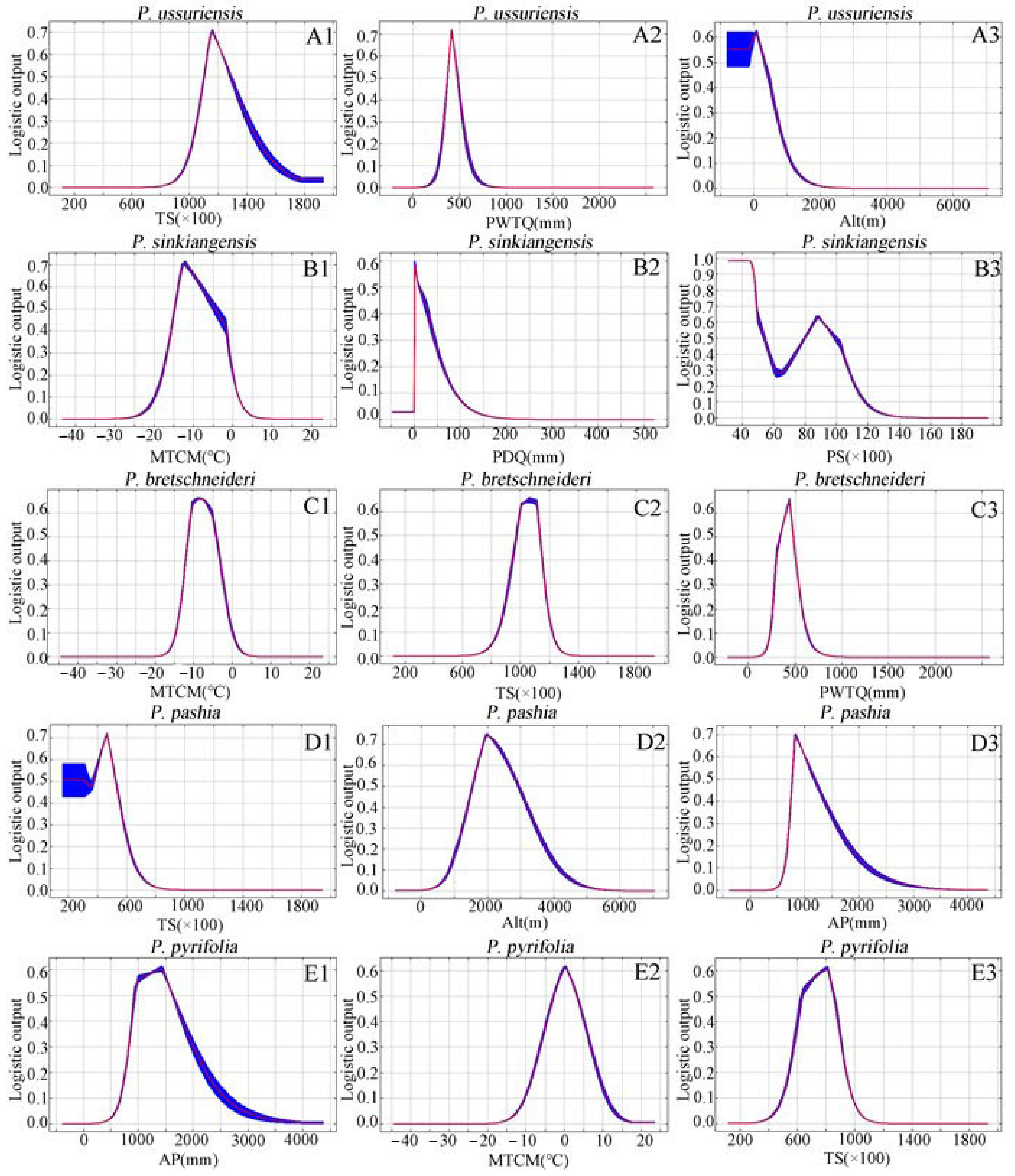
2.2. Percentage Contribution of Environmental Variables and Response Curve
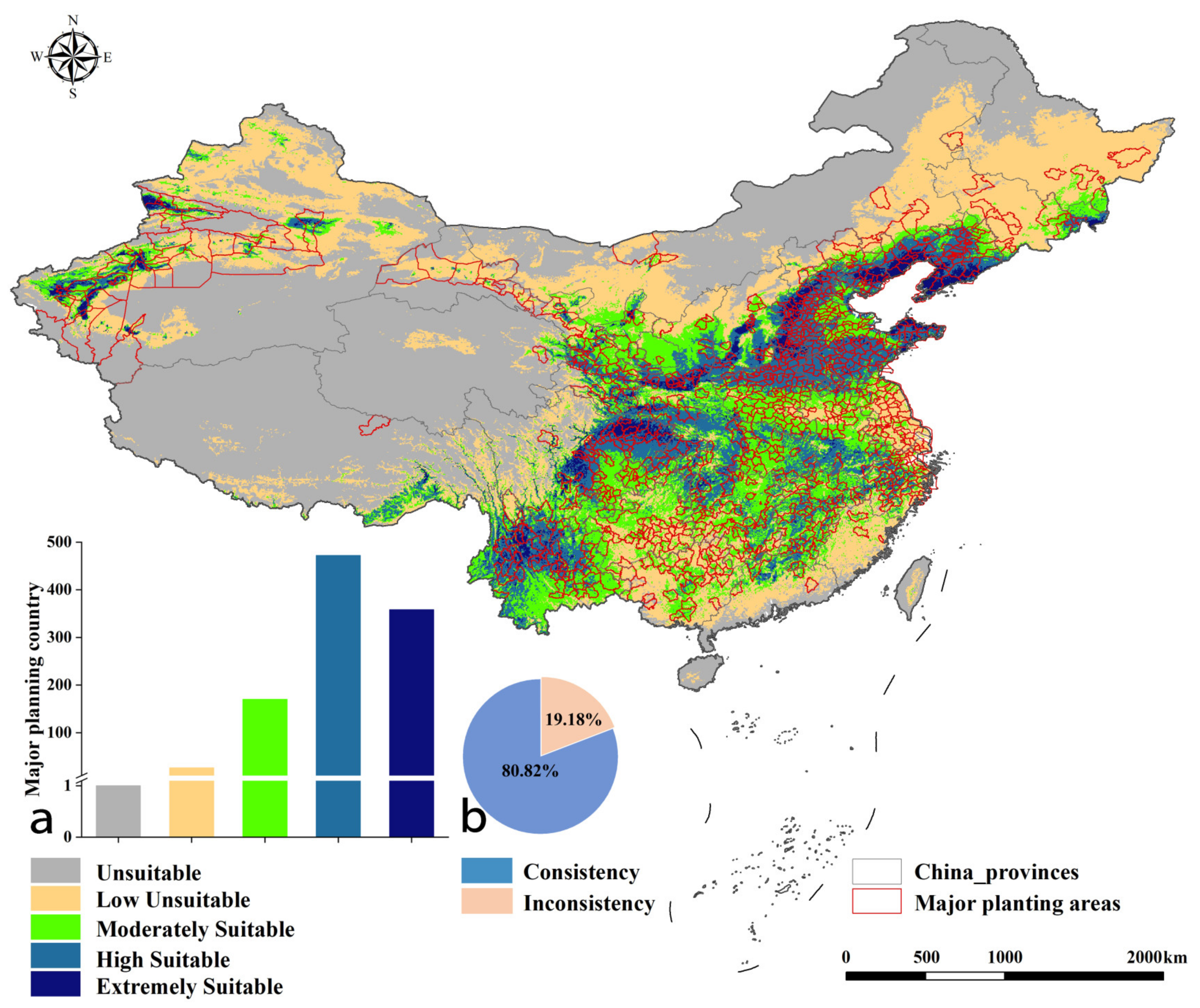
2.3. Suitable Planting Regions of Pyrus under Current Conditions
2.4. Consistency between the Potential Planting Regions and the Actual Main Production Counties under Current Climate Conditions
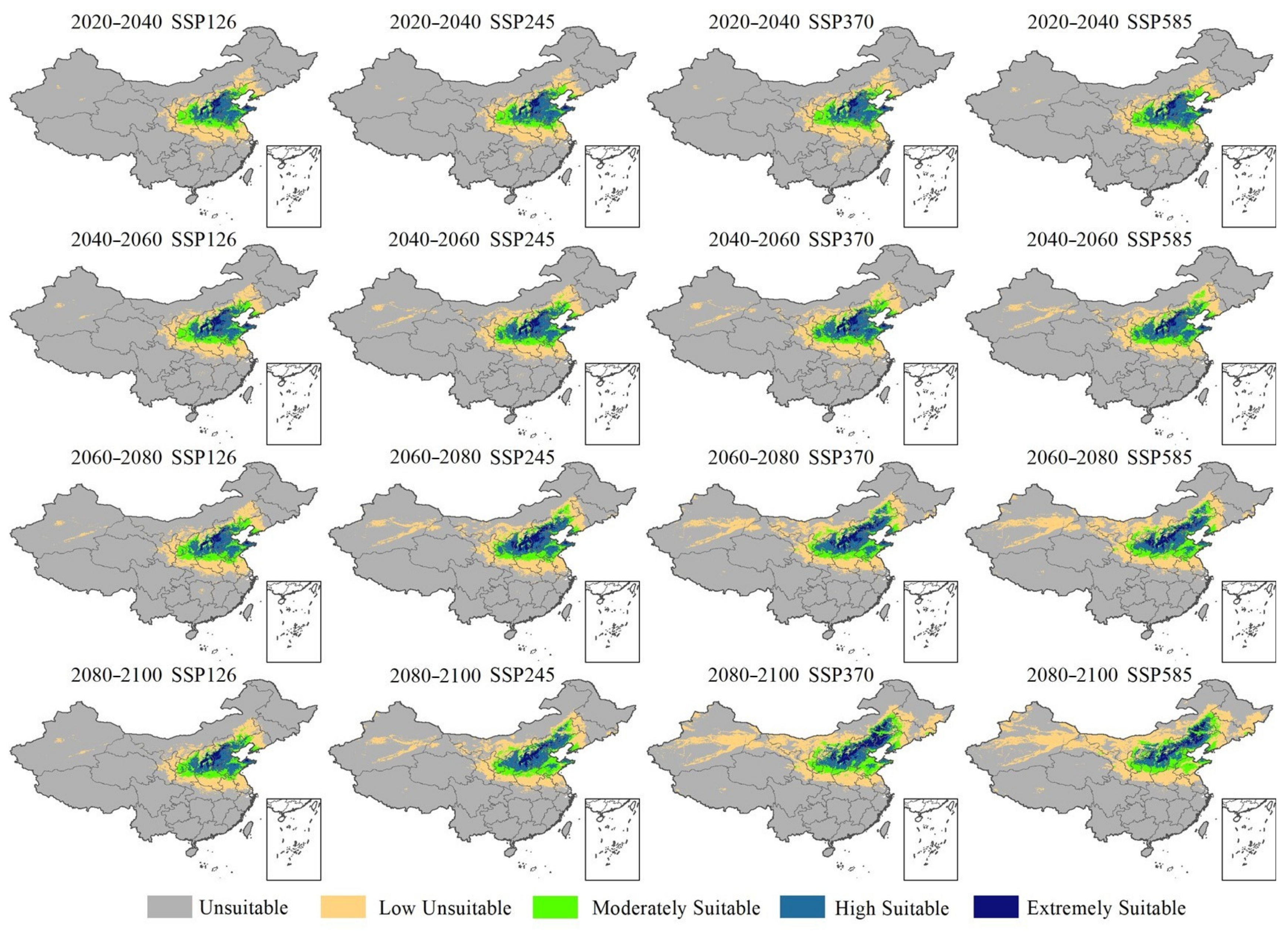
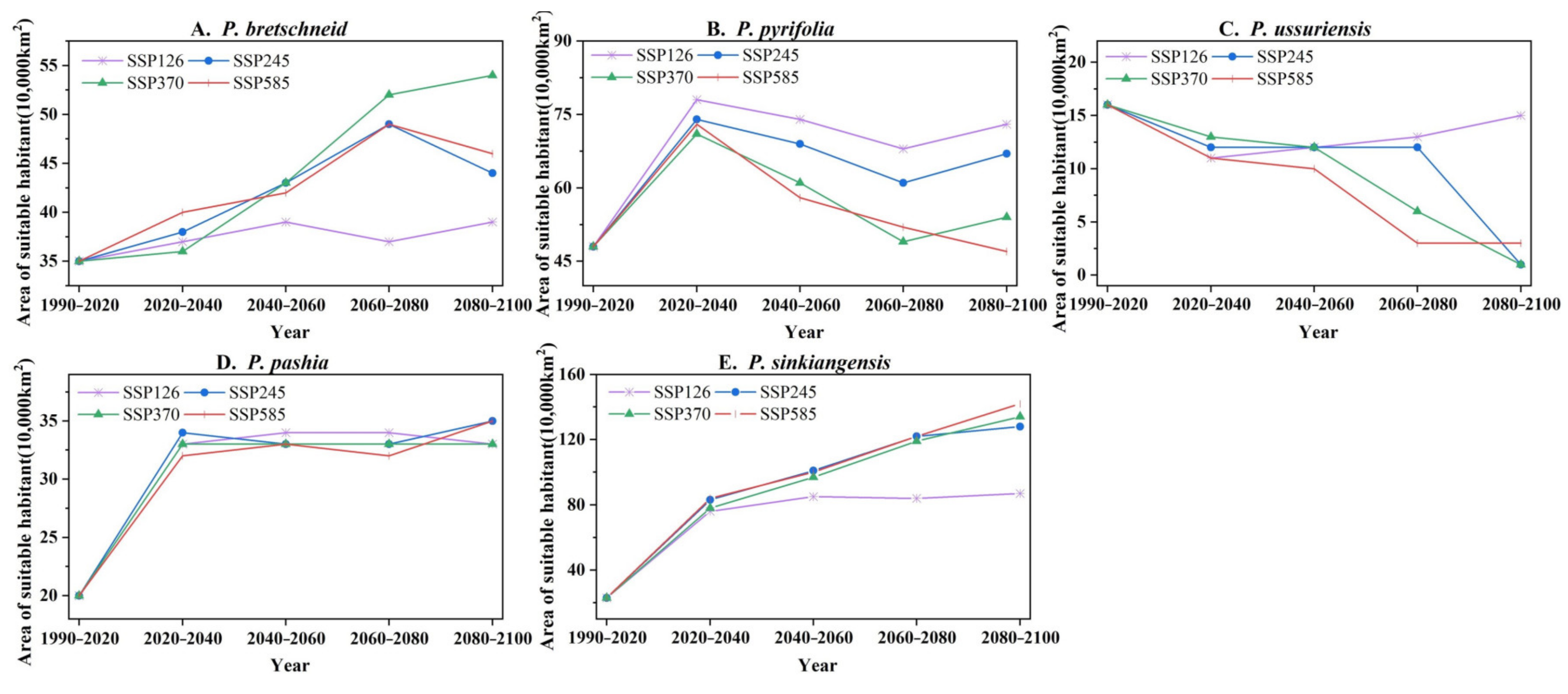
2.5. Future Changes in Potential Planting Areas for Pyrus of Different Species
3. Discussion
3.1. Differences in the Impact of Environmental Variables on Different Species of Pyrus
3.2. Changes in the Distribution Range of Pyrus and Climatic Risks of Planting
3.3. The Guiding Role of Species Distribution Models in Planting Management of Pyrus
3.4. Uncertainty and Limitation
4. Materials and Methods
4.1. Data Description
4.1.1. Species Distribution Data and Preprocessing
4.1.2. Geographical and Environmental Data
4.2. Model Construction
4.2.1. Model Introduction and Parameter Setting
4.2.2. Model Evaluation
4.3. Consistency Evaluation of Potential Planting Regions and Actual Main Production Regions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lenoir, J.; Gégout, J.-C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Vanalli, C.; Casagrandi, R.; Gatto, M.; Bevacqua, D. Shifts in the thermal niche of fruit trees under climate change: The case of peach cultivation in France. Agric. For. Meteorol. 2021, 300, 108327. [Google Scholar] [CrossRef]
- Sugiura, T. Three climate change adaptation strategies for fruit production. In Climate Smart Agriculture for the Small-Scale Farmers in the Asian and Pacific Region; Institute for Agro-Environmental Sciences: Tsukuba, Japan, 2021; p. 1096. [Google Scholar]
- Vitasse, Y.; Schneider, L.; Rixen, C.; Christen, D.; Rebetez, M. Increase in the risk of exposure of forest and fruit trees to spring frosts at higher elevations in Switzerland over the last four decades. Agric. For. Meteorol. 2018, 248, 60–69. [Google Scholar] [CrossRef]
- Fernandez, E.; Whitney, C.; Luedeling, E. The importance of chill model selection—A multi-site analysis. Eur. J. Agron. 2020, 119, 126103. [Google Scholar] [CrossRef]
- Silva, G.; Souza, T.M.; Barbieri, R.L.; Costa de Oliveira, A. Origin, domestication, and dispersing of pear (Pyrus spp.). Adv. Agric. 2014, 2014, 541097. [Google Scholar]
- FAO. Fruit and Vegetables—Your Dietary Essentials; The International Year of Fruits and Vegetables, 2021, Background Paper; FAO: Rome, Italy, 2020. [Google Scholar]
- FAO. FAO Statistic Yearbook 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- China Meteorological Administration. Blue Book on Climate Change in China; Science Press: Beijing, China, 2021. [Google Scholar]
- Li, B.; Chen, Y.; Chen, Z.; Xiong, H.; Lian, L. Why does precipitation in northwest China show a significant increasing trend from 1960 to 2010? Atmos. Res. 2016, 167, 275–284. [Google Scholar] [CrossRef]
- Liang, C.; Yin, S.; Li, M.; Zhang, Y. Variation of Climate and its impact on phonological period and growth of Pyrus bretschneideri Rehd in Binxian of Shaanxi Province in last 53 years. Agric. Res. Arid Areas 2012, 30, 190–194+217. [Google Scholar]
- Liu, C.; Huo, H.; Tian, L.; Dong, X.; Xu, J.; Qi, D.; Cao, Y. Prediction of potential geographical distribution patterns of Pyrus xerophila under different climate scenarios. Chin. J. Appl. Ecol. 2020, 31, 4073–4079. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Pacifici, M.; Visconti, P.; Butchart, S.H.M.; Watson, J.E.M.; Cassola, F.M.; Rondinini, C. Species’ traits influenced their response to recent climate change. Nat. Clim. Chang. 2017, 7, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Dos Santos, L.A.; Mendes, M.F.; Kruger, A.P.; Blauth, M.L.; Gottschalk, M.S.; Garcia, F.R. Global potential distribution of Drosophila suzukii (Diptera, Drosophilidae). PLoS ONE 2017, 12, e0174318. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Cabral, N.Y.; Kumar, L.; Shabani, F. Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX). Sci. Rep. 2017, 7, 5910. [Google Scholar] [CrossRef] [Green Version]
- Mi, C.; Huettmann, F.; Guo, Y.; Han, X.; Wen, L. Why choose Random Forest to predict rare species distribution with few samples in large undersampled areas? Three Asian crane species models provide supporting evidence. PeerJ 2017, 5, e2849. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 90, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Mukul, S.A.; Alamgir, M.; Sohel, M.S.I.; Pert, P.L.; Herbohn, J.; Turton, S.M.; Laurance, W.F. Combined effects of climate change and sea-level rise project dramatic habitat loss of the globally endangered Bengal tiger in the Bangladesh Sundarbans. Sci. Total Environ. 2019, 663, 830–840. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755 Pt 1, 142548. [Google Scholar] [CrossRef] [PubMed]
- Goberville, E.; Beaugrand, G.; Hautekeete, N.C.; Piquot, Y.; Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Gueguen7, M.; Renaud, J.; Karger, D.N.; Zimmermann, N.E. Uncertainty in ensembles of global biodiversity scenarios. Nat. Commun. 2019, 10, 1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Li, X.; Zhao, Z.; Nawaz, Z. Predicting the impacts of climate change, soils and vegetation types on the geographic distribution of Polyporus umbellatus in China. Sci. Total Environ. 2019, 648, 1–11. [Google Scholar] [CrossRef]
- Etherington, T.R. Python based GIS tools for landscape genetics: Visualising genetic relatedness and measuring landscape connectivity. Methods Ecol. Evol. 2011, 2, 52–55. [Google Scholar] [CrossRef]
- Chao, L.; Hong-liang, H.; Lu-ming, T.; Xing-guang, D.; Dan, Q.; Ying, Z.; Yu-fen, C. Potential geographical distribution of Pyrus calleryana under different climate change scenarios based on the MaxEnt mode. Chin. J. Appl. Ecol. 2018, 29, 3696–3704. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, H.t.; Song, H.w.; Zhang, Y.b.; Tao, R. Progress in Studies and Utilization of Hardy Fruit Germplasm Resources. J. Plant Genet. Resour. 2006, 1, 123–128. [Google Scholar] [CrossRef]
- Zhang, H. Analysis on the influence of climatic conditions on the growth of Korla Fragrant Pear. Meteorol. Mon. 2005, 31, 84–86. [Google Scholar]
- Li, X.; Yang, H.; Li, L.; Zhang, S.; Zhang, W. Resources and utilization evaluation of Pyrus in Yunnan. South China Fruits 2002, 3, 50–51. [Google Scholar]
- Thurm, E.A.; Hernandez, L.; Baltensweiler, A.; Ayan, S.; Rasztovits, E.; Bielak, K.; Falk, W. Alternative tree species under climate warming in managed European forests. Forest Ecol. Manag. 2018, 430, 485–497. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, Y.; Huang, D.; Wang, H.; Cao, Q.; Fan, P.; Wang, R. The effect of climate change on the richness distribution pattern of oaks (Quercus L.) in China. Sci. Total Environ. 2020, 744, 140786. [Google Scholar] [CrossRef]
- Fitchett, J.M.; Grab, S.W.; Thompson, D.; Roshan, G. Spatio-temporal variation in phenological response of citrus to climate change in Iran: 1960–2010. Agric. Forest Meteorol. 2014, 198, 285–293. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, D.; Chen, Y.; Kang, G.; Cheng, C. Empirical analysis of pear spatial distribution variation in China. China Fruits 2021, 4, 92–97. [Google Scholar] [CrossRef]
- CSY. China Statistical Yearbook; China Statistical Publishing House: Beijing, China, 2021. [Google Scholar]
- Peng, Y.; Gao, H.; Yuan, Z. Impact of Global Climate Change on Temperate Fruit Tree. J. Agric. Sci. Technol. 2018, 20, 1–10. [Google Scholar] [CrossRef]
- Perrigo, A.; Hoorn, C.; Antonelli, A. Why mountains matter for biodiversity. J. Biogeogr. 2019, 47, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant pathogen infection risk tracks global crop yields under climate change. Nat. Clim. Chang. 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Jin, Z.; Müller, C.; Pugh, T.A.M.; Chen, A.; Piao, S. Occurrence of crop pests and diseases has largely increased in China since 1970. Nat. Food 2022, 3, 57–65. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Zhao, B. Effects of climate change on agricultural meteorological disaster and crop insects diseases. Trans. CSAE 2010, 26, 263–271. [Google Scholar]
- Wang, Y.; Ji, X.; Wu, Y.; Mao, Z.; Jiang, Y.; Peng, F.; Chen, X. Research progress of cover crop in Chinese orchard. Chin. J. Appl. Ecol. 2015, 26, 1892–1900. [Google Scholar]
- Zheng, J.Y.; Bian, J.; Ge, Q.S.; Hao, Z.X.; Yin, Y.H.; Liao, Y.M. The climate regionalization in China for 1981–2010. Chin. Sci. Bull. 2013, 58, 3088–3099. [Google Scholar]
- Jiang, Z.; Tang, F.; Huang, H.; Hu, H.; Chen, Q. Assessment of genetic diversity of Chinese sand pear landraces (Pyrus pyrifolia Nakai) using simple sequence repeat markers. HortScience 2009, 44, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Shen, T. Pears in China. HortScience 1980, 15, 13–17. [Google Scholar] [CrossRef]
- Cao, S.; Li, T.; Song, H.; Liu, J. Map of Chinese Pear Local Varieties; China Forestry Publishing House: Beijing, China, 2018. [Google Scholar]
- Hortal, J.; Jiménez-Valverde, A.; Gómez, J.F.; Lobo, J.M.; Baselga, A. Historical bias in biodiversity inventories affects the observed environmental niche of the species. Oikos 2008, 117, 847–858. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Iglesias, T.L. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Peng, S.; Ding, Y.; Liu, W.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar]
- Robert, J.H.; Steven, P.; John, L.; Jane, E. dismo: Species Distribution Modeling, R package version 1.3-3; The R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Hijmans, R.J. terra: Spatial Data Analysis, R package version 1.3-4; The R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 28 March 2023).
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- Kriegler, E.; O’Neill, B.C.; Hallegatte, S.; Kram, T.; Lempert, R.J.; Moss, R.H.; Wilbanks, T. The need for and use of socio-economic scenarios for climate change analysis: A new approach based on shared socio-economic pathways. Glob. Environ. Chang. 2012, 22, 807–822. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Stehfest, E.; Gernaat, D.E.; Doelman, J.C.; van den Berg, M.; Harmsen, M.; de Boer, H.S.; Bouwman, L.F.; Daioglou, V.; Edelenbosch, O.Y.; et al. Edelenbosch. Energy, land-use and greenhouse gas emissions trajectories under a green growth paradigm. Glob. Environ. Chang. 2017, 42, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Riahi, K.; van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’Neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Chang. 2017, 42, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Fischer, G.; Nachtergaele, F.; Prieler, S.; Van Velthuizen, H.; Verelst, L.; Wiberg, D. Global Agro-Ecological Zones Assessment for Agriculture (GAEZ 2008); IIASA: Laxenburg, Austria; FAO: Rome, Italy, 2008. [Google Scholar]
- Yichen, Z.; Zengxin, Z.; Bin, Z.; Xuefei, C.; Liu, Y.; Mingkun, G.; Rui, K. MaxEnt Modeling Based on CMIP6 Models to Project Potential Suitable Zones for Cunninghamia lanceolata in China. Forests 2021, 12, 752. [Google Scholar]
- Phillips, S.J. Transferability, sample selection bias and background data in presence-only modelling: A response to Peterson et al. (2007). Ecography 2008, 31, 272–278. [Google Scholar] [CrossRef]
- Liu, G.; Han, R.; Cao, L. Artificial Cultivation of the Chinese Cordyceps from Injected Ghost Moth Larvae. Environ. Entomol. 2019, 48, 1088–1094. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Anderson, B. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evo. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Jayasinghe, S.L.; Kumar, L. Modeling the climate suitability of tea [Camellia sinensis (L.) O. Kuntze] in Sri Lanka in response to current and future climate change scenarios. Agric. For. Meteorol. 2019, 272–273, 102–117. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Bariotakis, M.; Georgescu, L.; Laina, D.; Oikonomou, I.; Ntagounakis, G.; Koufaki, M.I.; Pirintsos, S.A. From wild harvest towards precision agriculture: Use of Ecological Niche Modelling to direct potential cultivation of wild medicinal plants in Crete. Sci. Total. Environ. 2019, 694, 133681. [Google Scholar] [CrossRef]
- McPherson, J.M.; Jetz, W.; Rogers, D.J. The effects of species’ range sizes on the accuracy of distribution models: Ecological phenomenon or statistical artefact? J. Appl. Ecol. 2004, 41, 811–823. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- National Forestry and Grassland Administration. China Forestry Industry and Forest Products Yearbook 2018; China Forestry Publishing House: Beijing, China, 2018. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Contribution% | ||||
|---|---|---|---|---|---|
| P. bretschneideri | P. sinkiangensis | P. pashia | P. pyrifolia | P. ussuriensis | |
| AMT | 1.2 | 1 | 0.7 | 0.5 | 9.4 |
| MDR | 1.1 | 3.9 | 2.6 | 1.8 | 0.1 |
| TS | 25.5 | 9.5 | 56.6 | 13.5 | 37.5 |
| MTCM | 33.7 | 26 | 3.3 | 20.7 | 6.1 |
| AP | 0.3 | 0.2 | 13.1 | 53.9 | 0.1 |
| PS | 9.5 | 12.8 | 0.2 | 1 | 5.8 |
| PWTQ | 15.5 | 3.8 | 0.1 | 0.2 | 25.1 |
| PDQ | 0.3 | 15.1 | 4.2 | 3.6 | 0.5 |
| ALT | 11.1 | 12.3 | 17.2 | 2.5 | 14.7 |
| S_SAND | 0.5 | 1 | 0.2 | 0.7 | 0.6 |
| S_OC | 0.4 | 7.7 | 0.1 | 0.7 | 0 |
| S_PH_H2O | 1.1 | 6.6 | 1.5 | 1 | 0.1 |
| Code | Environment Variables | Units |
|---|---|---|
| AMT | Annual mean temperature | ℃ |
| MDR | Mean diurnal range | °C |
| ISO | Isothermality | ×100 |
| TS | Temperature seasonality | ×100 |
| MTWM | Max. temperature for warmest month | °C |
| MTCM | Min. temperature for coldest month | °C |
| TAR | Temperature annual range | ℃ |
| MTWTQ | Mean temperature of wettest quarter | °C |
| MTDQ | Mean temperature of driest quarter | °C |
| MTWRQ | Mean temperature of warmest quarter | °C |
| MTCQ | Mean temperature of coldest quarter | °C |
| AP | Annual precipitation | mm |
| PWM | Precipitation of wettest month | mm |
| PDM | Precipitation of driest month | mm |
| PS | Precipitation seasonality | ×100 |
| PWTQ | Precipitation of wettest quarter | mm |
| PDQ | Precipitation of driest quarter | mm |
| PWRQ | Precipitation of warmest quarter | mm |
| PCQ | Precipitation of coldest quarter | mm |
| Alt | Altitude | m |
| S_SAND | Subsoil sand fraction | % wt |
| S_OC | Subsoil organic carbon | % weight |
| S_PH_H2O | Subsoil PH (H2O) | −log(H+) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Hu, Z.; Wang, Y.; Zhao, W. Spatial Distribution Characteristics of Suitable Planting Areas for Pyrus Species under Climate Change in China. Plants 2023, 12, 1559. https://doi.org/10.3390/plants12071559
Wang M, Hu Z, Wang Y, Zhao W. Spatial Distribution Characteristics of Suitable Planting Areas for Pyrus Species under Climate Change in China. Plants. 2023; 12(7):1559. https://doi.org/10.3390/plants12071559
Chicago/Turabian StyleWang, Mi, Zhuowei Hu, Yongcai Wang, and Wenji Zhao. 2023. "Spatial Distribution Characteristics of Suitable Planting Areas for Pyrus Species under Climate Change in China" Plants 12, no. 7: 1559. https://doi.org/10.3390/plants12071559