
Seed Priming with the Selenium Nanoparticles Maintains the Redox Status in the Water Stressed Tomato Plants by Modulating the Antioxidant Defense Enzymes

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
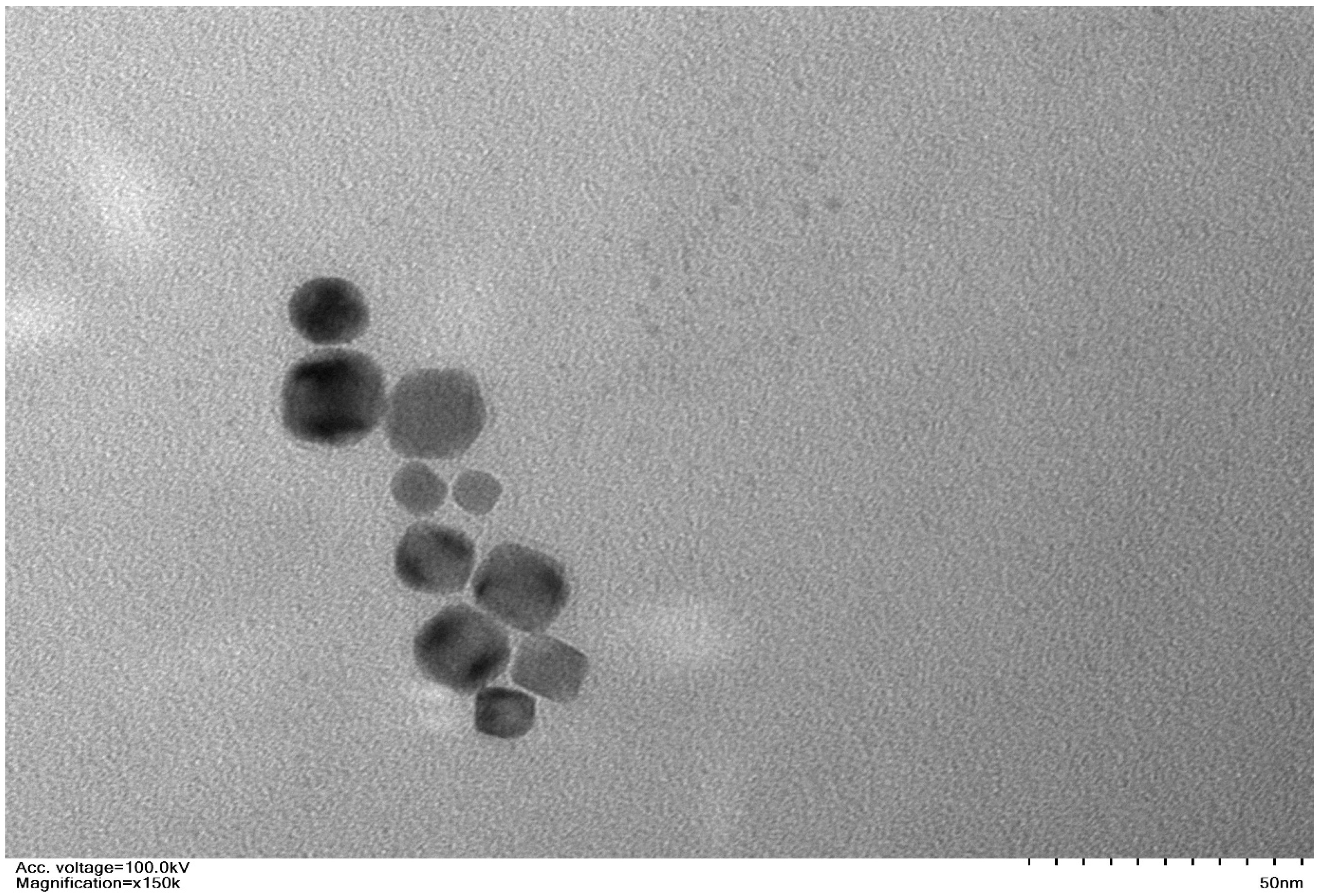
2. Results
3. Discussion
3.1. SeNPs-Mediated Seed Priming Increases Biomass and Photosynthetic Ability
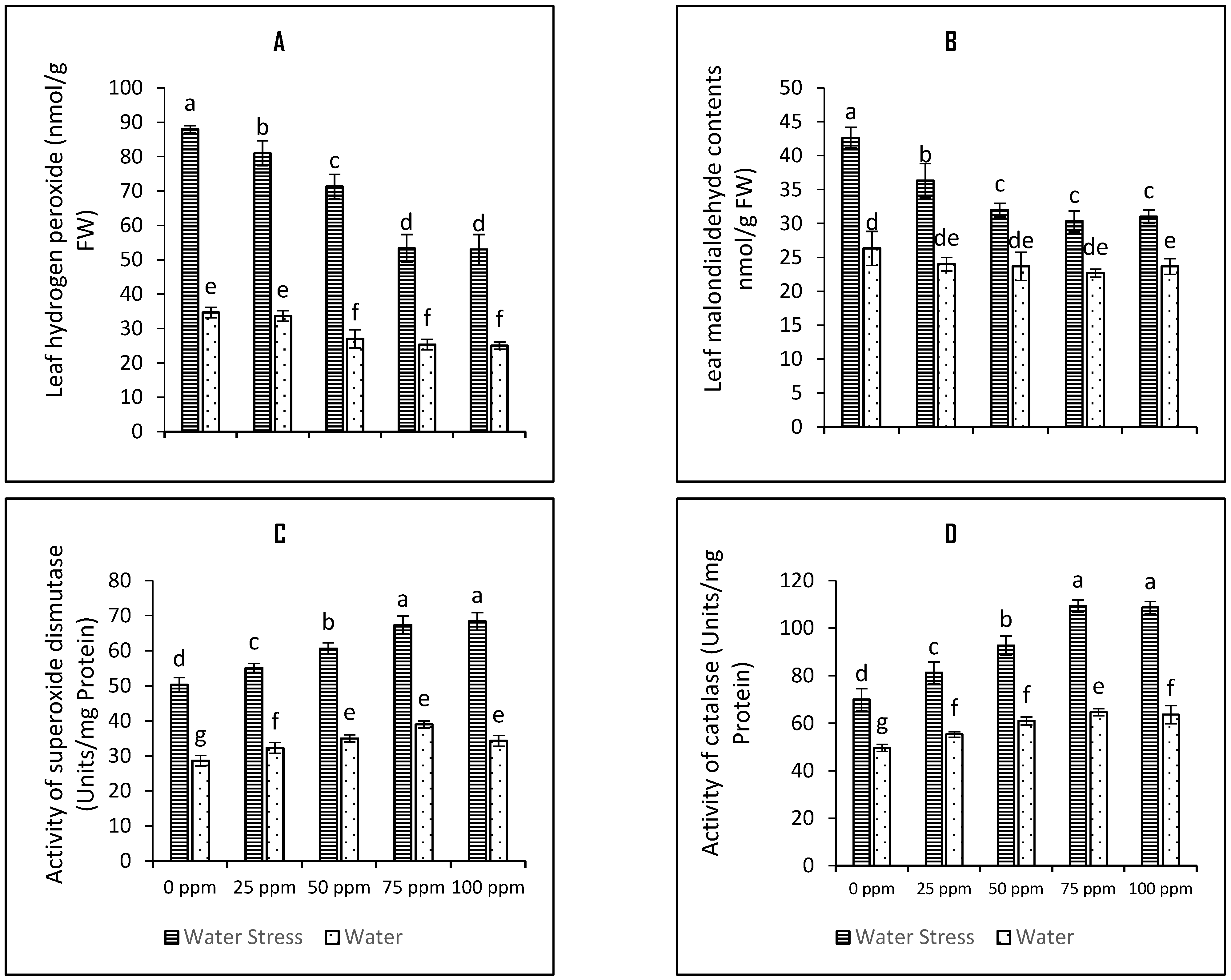
3.2. Impact on Antioxidant Defence Enzymes and Osmotic Stress Markers
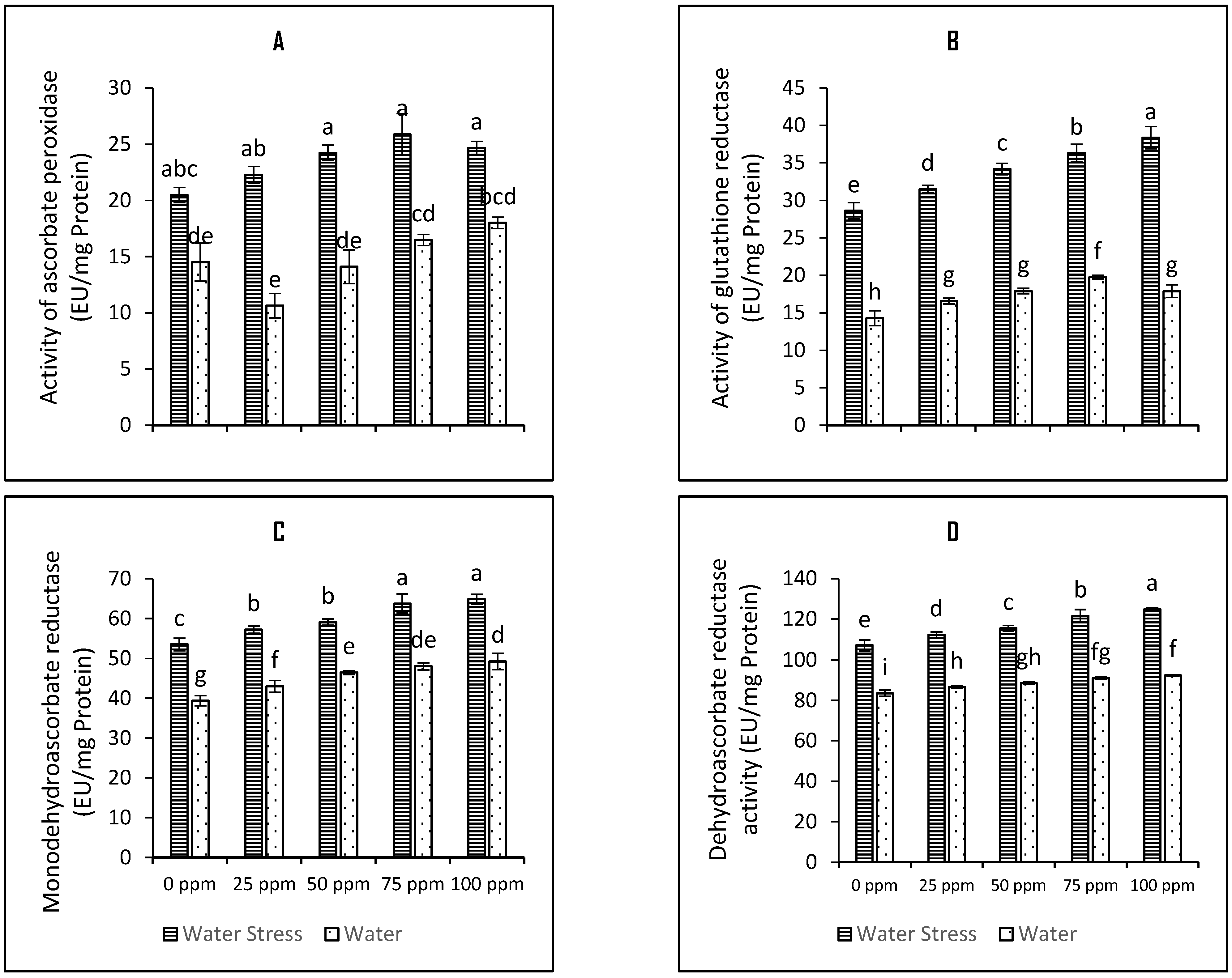
3.3. Modulation of Ascorbate Glutathione Cycle Enzymes
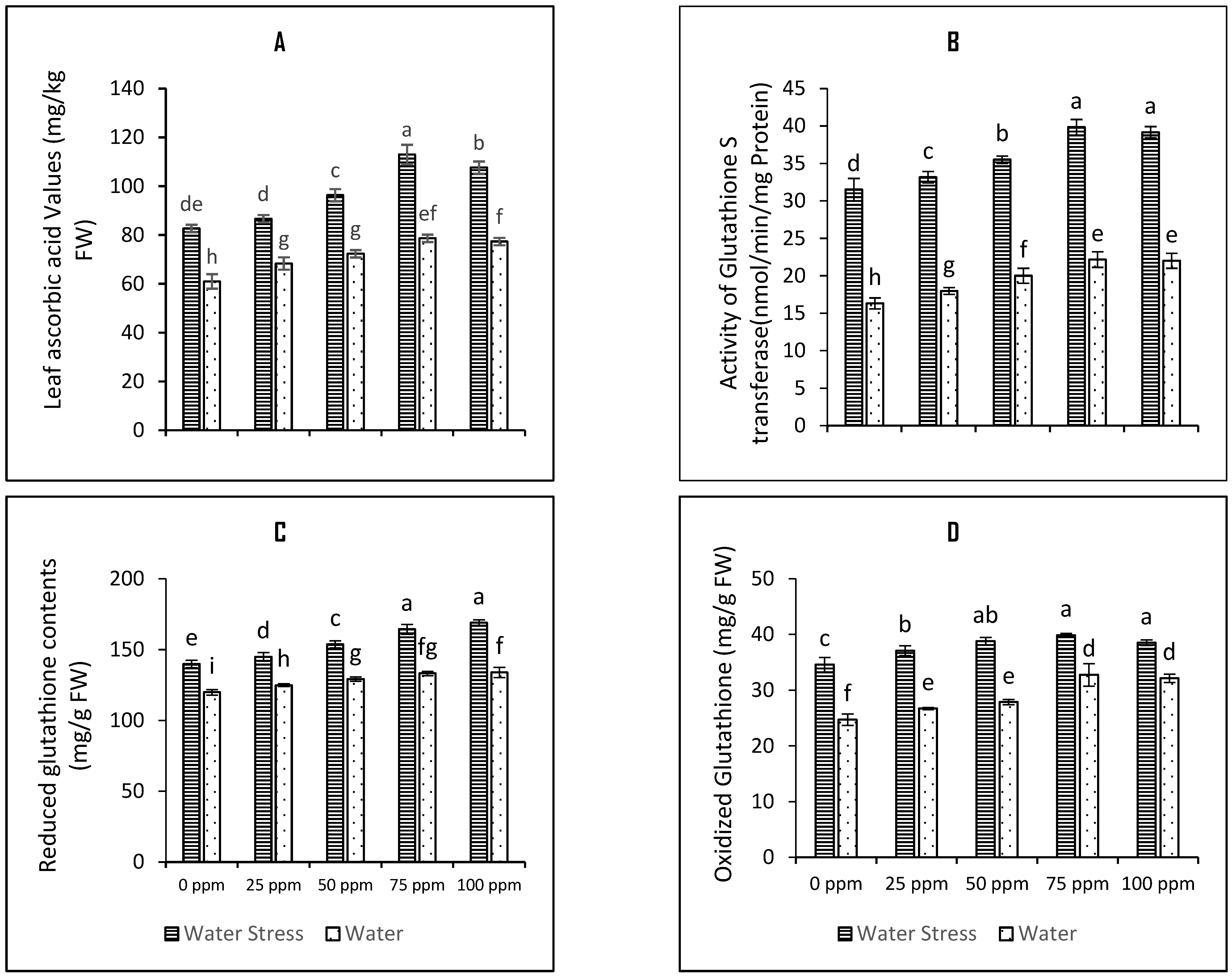
3.4. SeNPs-Mediated Seed Priming Lifts Ascorbate Glutathione Substrate Pool
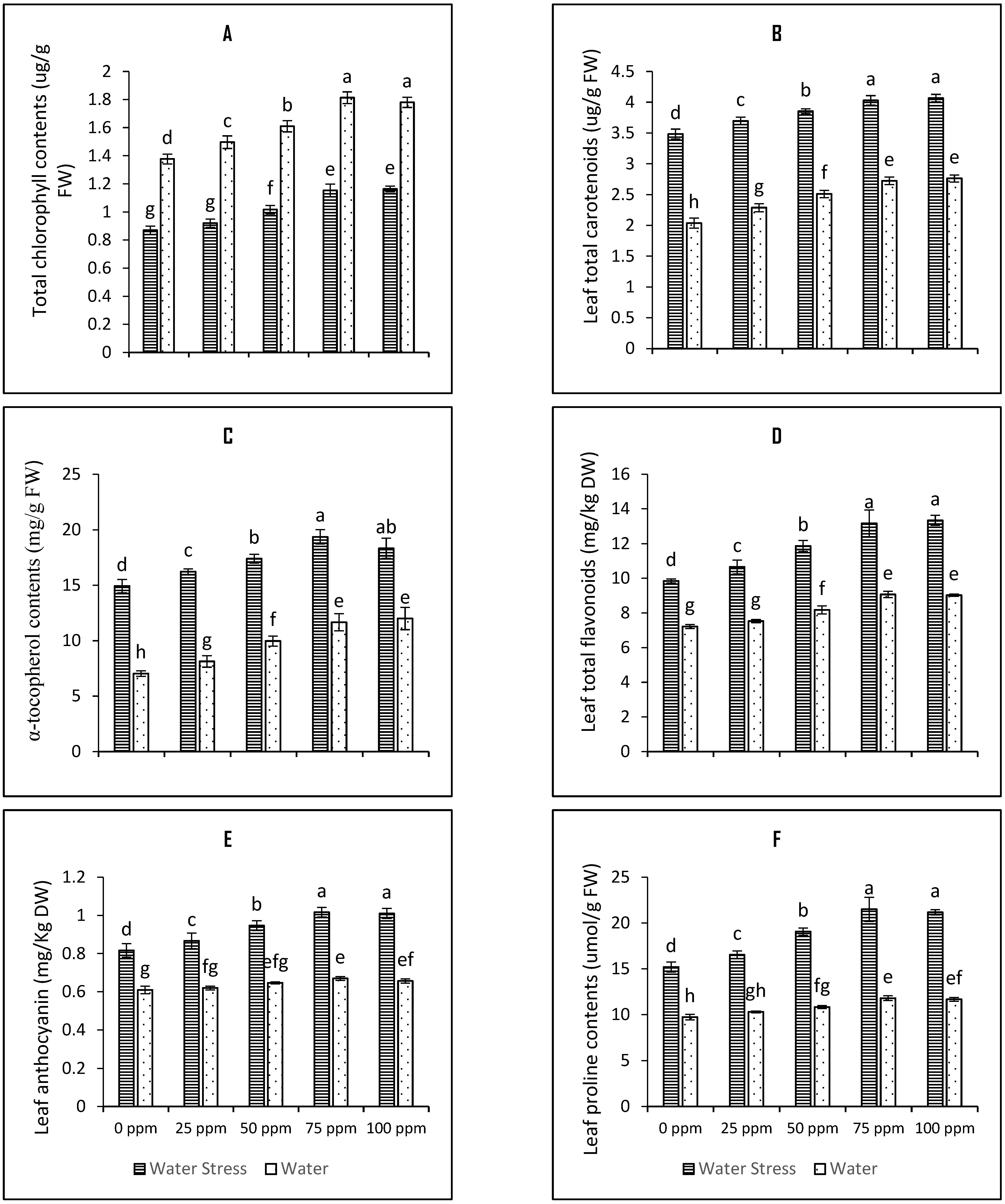
3.5. SeNPs-Mediated Seed Priming Increases Nonenzymatic Antioxidants and Secondary Metabolites Conferring Drought Tolerance
4. Materials and Methods
4.1. Experimental Setup and Treatment Plan
4.2. Observations on Growth Parameters
4.3. Determination of MDA Contents
4.4. Determination of Hydrogen Peroxide Contents
4.5. Activities of Antioxidative Defence Enzymes
4.6. Determination of Leaf Chlorophyll Contents
4.7. Determination of Ascorbic Acid (AsA) Contents
4.8. Determination of Total Anthocyanin Contents
4.9. Total Flavonoid Contents
4.10. Determination of Proline Contents
4.11. Analysis of Glutathione S-Transferase (GST) Functioning
4.12. Determination of Ascorbate-Glutathione Pathway Enzymes Activity
4.13. Determination of Tocopherols
4.14. Measurement of Reduced and Oxidised Glutathione Contents
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, L.; Zhao, X.; Gao, X.; Siddique, K.H. Improving/maintaining water-use efficiency and yield of wheat by deficit irrigation: A global meta-analysis. Agric. Water Manag. 2020, 228, 105906. [Google Scholar] [CrossRef]
- Routson, C.C.; McKay, N.P.; Kaufman, D.S.; Erb, M.P.; Goosse, H.; Shuman, B.N.; Rodysill, J.R.; Ault, T. Mid-latitude net precipitation decreased with Arctic warming during the Holocene. Nature 2019, 568, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increases and its effects on fish stress physiology in the context of global warming. J. Fish Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Yang, B.; Chen, Y.; Shi, J. Reactive oxygen species (ROS)-based nanomedicine. Chem. Rev. 2019, 119, 4881–4985. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Ahmad, P.; Alyemeni, M.N.; Al-Huqail, A.A.; Alqahtani, M.A.; Wijaya, L.; Ashraf, M.; Kaya, C.; Bajguz, A. Zinc oxide nanoparticles application alleviates arsenic (As) toxicity in soybean plants by restricting the uptake of as and modulating key biochemical attributes, antioxidant enzymes, ascorbate-glutathione cycle and glyoxalase system. Plants 2020, 9, 825. [Google Scholar] [CrossRef]
- Sadeghi, F.; Samsampour, D.; Seyahooei, M.A.; Bagheri, A.; Soltani, J. Fungal endophytes alleviate drought-induced oxidative stress in mandarin (Citrus reticulata L.): Toward regulating the ascorbate–glutathione cycle. Sci. Hortic. 2020, 261, 108991. [Google Scholar] [CrossRef]
- Furlan, A.L.; Bianucci, E.; Giordano, W.; Castro, S.; Becker, D.F. Proline metabolic dynamics and implications in drought tolerance of peanut plants. Plant Physiol. Biochem. 2020, 151, 566–578. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Li, H.J.; Zhao, C.F.; Xue, J.Q.; Zhang, R.H. Exogenous melatonin improves drought tolerance in maize seedlings by regulating photosynthesis and the ascorbate–glutathione cycle. Russ. J. Plant Physiol. 2020, 67, 809–821. [Google Scholar] [CrossRef]
- Alharby, H.F.; Al-Zahrani, H.S.; Alzahrani, Y.M.; Alsamadany, H.; Hakeem, K.R.; Rady, M.M. Maize grain extract enriched with polyamines alleviates drought stress in Triticum aestivum through up-regulation of the ascorbate–glutathione cycle, glyoxalase system, and polyamine gene expression. Agronomy 2020, 11, 949. [Google Scholar] [CrossRef]
- Kour, D.; Khan, S.S.; Kaur, T.; Kour, H.; Singh, G.; Yadav, A.; Yadav, A.N. Drought adaptive microbes as bioinoculants for the horticultural crops. Heliyon 2022, 8, e09493. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease management of tomato through PGPB: Current trends and future perspective. Biotech 2017, 7, 255. [Google Scholar] [CrossRef]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [Green Version]
- Badar, H.; Rasool, A.; Boye, M. Consumer segments and value preferences for tomatoes in Pakistan. J. Anim. Plant Sci 2021, 31, 246–253. [Google Scholar]
- Zhang, Z.; Cao, B.; Gao, S.; Xu, K. Grafting improves tomato drought tolerance through enhancing photosynthetic capacity and reducing ROS accumulation. Protoplasma 2019, 256, 1013–1024. [Google Scholar] [CrossRef]
- Krishna, R.; Ansari, W.A.; Soumia, P.S.; Yadav, A.; Jaiswal, D.K.; Kumar, S.; Singh, A.K.; Singh, M.; Verma, J.P. Biotechnological Interventions in Tomato (Solanum lycopersicum) for Drought Stress Tolerance: Achievements and Future Prospects. BioTech 2022, 11, 48. [Google Scholar] [CrossRef]
- Ors, S.; Ekinci, M.; Yildirim, E.; Sahin, U.; Turan, M.; Dursun, A. Interactive effects of salinity and drought stress on photosynthetic characteristics and physiology of tomato (Lycopersicon esculentum L.) seedlings. South Afr. J. Bot. 2021, 137, 335–339. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qi, M.; Huang, Z.; Xu, X.; Begum, N.; Qin, C.; Zhang, C.; Ahmad, N.; Mustafa, N.S.; Ashraf, M.; et al. Improving growth and photosynthetic performance of drought stressed tomato by application of nano-organic fertilizer involves up-regulation of nitrogen, antioxidant and osmolyte metabolism. Ecotoxicol. Environ. Saf. 2021, 216, 112195. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin improves drought stress tolerance of tomato by modulating plant growth, root architecture, photosynthesis, and antioxidant defense system. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef]
- Rai, G.K.; Parveen, A.; Jamwal, G.; Basu, U.; Kumar, R.R.; Rai, P.K.; Sharma, J.P.; Alalawy, A.I.; Al-Duais, M.A.; Hossain, M.A.; et al. Leaf proteome response to drought stress and antioxidant potential in tomato (Solanum lycopersicum L.). Atmosphere 2021, 12, 1021. [Google Scholar] [CrossRef]
- Mazhar, M.W.; Ishtiaq, M.; Maqbool, M.; Akram, R. Seed priming with Calcium oxide nanoparticles improves germination, biomass, antioxidant defence and yield traits of canola plants under drought stress. South Afr. J. Bot. 2022, 151, 889–899. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Rauf, M.A.; Jolly, J.; Ahmad, Z. Synthesis and Characterization of Nano-Selenium Using Plant Biomolecules and Their Potential Applications. In Selenium and Nano-Selenium in Environmental Stress Management and Crop Quality Improvement; Springer International Publishing: Cham, Switzerland, 2022; pp. 25–40. [Google Scholar]
- Garza-García, J.J.; Hernández-Díaz, J.A.; Zamudio-Ojeda, A.; León-Morales, J.M.; Guerrero-Guzmán, A.; Sánchez-Chiprés, D.R.; López-Velázquez, J.C.; García-Morales, S. The role of selenium nanoparticles in agriculture and food technology. Biol. Trace Elem. Res. 2021, 200, 2528–2548. [Google Scholar] [CrossRef] [PubMed]
- Abedi, S.; Iranbakhsh, A.; Oraghi Ardebili, Z.; Ebadi, M. Nitric oxide and selenium nanoparticles confer changes in growth, metabolism, antioxidant machinery, gene expression, and flowering in chicory (Cichorium intybus L.): Potential benefits and risk assessment. Environ. Sci. Pollut. Res. 2021, 28, 3136–3148. [Google Scholar] [CrossRef] [PubMed]
- Alvi, G.B.; Iqbal, M.S.; Ghaith, M.M.S.; Haseeb, A.; Ahmed, B.; Qadir, M.I. Biogenic selenium nanoparticles (SeNPs) from citrus fruit have anti-bacterial activities. Sci. Rep. 2021, 11, 4811. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, M.; Akram, A.; Raja, N.I.; Mukhtar, T.; Mehak, A.; Fatima, N.; Ajmal, M.; Ali, K.; Mustafa, N.; Abasi, F. Antifungal activity of green synthesized selenium nanoparticles and their effect on physiological, biochemical, and antioxidant defense system of mango under mango malformation disease. PLoS ONE 2023, 18, e0274679. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V. High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 2018, 3, 2479–2491. [Google Scholar] [CrossRef] [Green Version]
- El-Gazzar, N.; Ismail, A.M. The potential use of Titanium, Silver and Selenium nanoparticles in controlling leaf blight of tomato caused by Alternaria alternata. Biocatal. Agric. Biotechnol. 2020, 27, 101708. [Google Scholar] [CrossRef]
- Morales-Espinoza, M.C.; Cadenas-Pliego, G.; Pérez-Alvarez, M.; Hernández-Fuentes, A.D.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Valdés-Reyna, J.; Juárez-Maldonado, A. Se nanoparticles induce changes in the growth, antioxidant responses, and fruit quality of tomato developed under NaCl stress. Molecules 2019, 24, 3030. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Feng, X.; Wei, L.; Chen, L.; Song, B.; Shao, L. The toxicology of ion-shedding zinc oxide nanoparticles. Crit. Rev. Toxicol. 2016, 46, 348–384. [Google Scholar] [CrossRef] [PubMed]
- Skalickova, S.; Milosavljevic, V.; Cihalova, K.; Horky, P.; Richtera, L.; Adam, V. Selenium nanoparticles as a nutritional supplement. Nutrition 2017, 33, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Liang, X.; Nong, K.; Gong, Z.; Qin, T.; Qin, X.; Wang, D.; Zhu, Y. Advances in research on the toxicological effects of selenium. Bull. Environ. Contam. Toxicol. 2021, 106, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; EL Sabagh, A. Evaluation of fourteen bread wheat (Triticum aestivum L.) genotypes by observing gas exchange parameters, relative water and chlorophyll content, and yield attributes under drought stress. Sustainability 2021, 13, 4799. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Johnson, J.; Shanmugam, R.; and Lakshmi, T. A review on plant-mediated selenium nanoparticles and its applications. J. Popul. Ther. Clin. Pharmacol. 2022, 28, 29–40. [Google Scholar]
- Zahedi, S.M.; Hosseini, M.S.; Daneshvar Hakimi Meybodi, N.; Peijnenburg, W. Mitigation of the effect of drought on growth and yield of pomegranates by foliar spraying of different sizes of selenium nanoparticles. J. Sci. Food Agric. 2021, 101, 5202–5213. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, S.; Biswas, B.; Chakraborty, D.; Timsina, J.; Pal, S.; Chandra Tarafdar, J.; Banerjee, S.; Hossain, A.; Roy, S. Seed priming with selenium and zinc nanoparticles modifies germination, growth, and yield of direct-seeded rice (Oryza sativa L.). Sci. Rep. 2022, 12, 7103. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Qu, J.; Pu, Y.; Rao, S.; Xu, F.; Wu, C. Selenium biofortification of crop food by beneficial microorganisms. J. Fungi 2020, 6, 59. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Vangnai, A.S.; Cornejo, P. R hodotorula mucilaginosa CAM4 improved selenium uptake in Spinacia oleracea L.; soil enzymatic activities under abiotic stresses. Environ. Sci. Pollut. Res. 2022, 29, 89943–89953. [Google Scholar] [CrossRef] [PubMed]
- Sotoodehnia-Korani, S.; Iranbakhsh, A.; Ebadi, M.; Majd, A.; Ardebili, Z.O. Selenium nanoparticles induced variations in growth, morphology, anatomy, biochemistry, gene expression, and epigenetic DNA methylation in Capsicum annuum; an in vitro study. Environ. Pollut. 2020, 265, 114727. [Google Scholar] [CrossRef]
- Nasibi, F.; Aminian, F.; Mohammadinejad, G.; Hassanshahian, M. Seed priming with selenium nanoparticle and plant growth promoting rhizobacteria improve seedling development of foxtail millet (Setaria italica) under salinity stress. Research Square 2022. [Google Scholar] [CrossRef]
- Salam, A.; Afridi, M.S.; Javed, M.A.; Saleem, A.; Hafeez, A.; Khan, A.R.; Zeeshan, M.; Ali, B.; Azhar, W.; Ulhassan, Z.; et al. Nano-priming against abiotic stress: A way forward towards sustainable agriculture. Sustainability 2022, 14, 14880. [Google Scholar] [CrossRef]
- Khalofah, A.; Migdadi, H.; El-Harty, E. Antioxidant enzymatic activities and growth response of quinoa (Chenopodium quinoa willd) to exogenous selenium application. Plants 2021, 10, 719. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Cheema, M.A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Nawaz, A.; Abbas, T.; Ali, Q. Physiological and biochemical attributes of bread wheat (Triticum aestivum L.) seedlings are influenced by foliar application of silicon and selenium under water deficit. Acta Physiol. Plant. 2019, 41, 1–11. [Google Scholar] [CrossRef]
- Zakeri, N.; Asbaghi, O.; Naeini, F.; Afsharfar, M.; Mirzadeh, E.; kasra Naserizadeh, S. Selenium supplementation and oxidative stress: A review. PharmaNutrition 2021, 17, 100263. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Mohamed, I.A.; Wang, Z.; Wang, C.; Tabl, K.M.; Khatab, A.; Kuai, J.; Wang, J.; Wang, B.; et al. Mitigation of the salinity stress in rapeseed (Brassica napus L.) productivity by exogenous applications of bio-selenium nanoparticles during the early seedling stage. Environ. Pollut. 2022, 310, 119815. [Google Scholar] [CrossRef]
- Soleymanzadeh, R.; Iranbakhsh, A.; Habibi, G.; Ardebili, Z.O. Selenium nanoparticle protected strawberry against salt stress through modifications in salicylic acid, ion homeostasis, antioxidant machinery, and photosynthesis performance. Acta Biol. Crac. s. Bot. 2020, 62, 33–42. [Google Scholar]
- Kiumarzi, F.; Morshedloo, M.R.; Zahedi, S.M.; Mumivand, H.; Behtash, F.; Hano, C.; Chen, J.T.; Lorenzo, J.M. Selenium Nanoparticles (Se-NPs) Alleviates Salinity Damages and Improves Phytochemical Characteristics of Pineapple Mint (Mentha suaveolens Ehrh.). Plants 2022, 11, 1384. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.J.; Blasco, B.; Cervilla, L.M.; Rosales, M.A.; Sanchez-Rodriguez, E.; Romero, L.; Ruiz, J.M. Production and detoxification of H2O2 in lettuce plants exposed to selenium. Ann. Appl. Biol. 2009, 154, 107–116. [Google Scholar] [CrossRef]
- Lanza, M.G.D.B.; Dos Reis, A.R. Roles of selenium in mineral plant nutrition: ROS scavenging responses against abiotic stresses. Plant Physiol. Biochem. 2021, 164, 27–43. [Google Scholar] [CrossRef]
- Kaur, A.; Tyagi, S.; Singh, K.; Upadhyay, S.K. Exploration of glutathione reductase for abiotic stress response in bread wheat (Triticum aestivum L.). Plant Cell Rep. 2022, 41, 639–654. [Google Scholar]
- Ha, H.Y.; Alfulaij, N.; Berry, M.J.; Seale, L.A. From selenium absorption to selenoprotein degradation. Biol. Trace Elem. Res. 2019, 192, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Wang, B.; Han, Y.; Li, S. The pivotal function of dehydroascorbate reductase in glutathione homeostasis in plants. J. Exp. Bot. 2020, 71, 3405–3416. [Google Scholar] [CrossRef] [PubMed]
- Khalil, R.; Yusuf, M.; Bassuony, F.; Haroun, S.; Gamal, A. Alpha-tocopherol reinforce selenium efficiency to ameliorates salt stress in maize plants through carbon metabolism, enhanced photosynthetic pigments and ion uptake. South Afr. J. Bot. 2022, 144, 1–9. [Google Scholar] [CrossRef]
- Sun, X.; Han, G.; Ye, S.; Luo, Y.; Zhou, X. Effects of selenium on serotonin synthesis and the glutathione redox cycle in plum leaves. J. Soil Sci. Plant Nutr. 2020, 20, 2212–2221. [Google Scholar] [CrossRef]
- Moursi, Y.S.; Dawood, M.F.; Sallam, A.; Thabet, S.G.; Alqudah, A.M. Antioxidant enzymes and their genetic mechanism in alleviating drought stress in plants. In Organic Solutes, Oxidative Stress, and Antioxidant Enzymes Under Abiotic Stressors; CRC Press: Boca Raton, FL, USA, 2021; pp. 233–262. [Google Scholar]
- Bagherzadeh Homaee, M.; and Ehsanpour, A.A. Silver nanoparticles and silver ions: Oxidative stress responses and toxicity in potato (Solanum tuberosum L.) grown in vitro. Hortic. Environ. Biotechnol. 2016, 57, 544–553. [Google Scholar] [CrossRef]
- Mittler, R.; Zilinskas, B.A. Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. Plant J. 1994, 5, 397–405. [Google Scholar] [CrossRef]
- Koramutla, M.K.; Negi, M.; Ayele, B.T. Roles of glutathione in mediating abscisic acid signaling and its regulation of seed dormancy and drought tolerance. Genes 2021, 12, 1620. [Google Scholar] [CrossRef]
- Zechmann, B. Subcellular roles of glutathione in mediating plant defense during biotic stress. Plants 2020, 9, 1067. [Google Scholar] [CrossRef] [PubMed]
- Estévez, I.H.; Hernández, M.R. Plant glutathione S-transferases: An overview. Plant Gene 2020, 23, 100233. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Belal Chowdhury, M.; Afrin, S.; Burritt, D.J.; Murata, Y.; Hossain, M.A.; Afzal Hossain, M. Exogenous glutathione-mediated drought stress tolerance in rice (Oryza sativa L.) is associated with lower oxidative damage and favorable ionic homeostasis. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 955–971. [Google Scholar] [CrossRef]
- Sahay, S.; Khan, E.; Gupta, M. Nitric oxide and abscisic acid protects against PEG-induced drought stress differentially in Brassica genotypes by combining the role of stress modulators, markers and antioxidants. Nitric Oxide 2019, 89, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çelik, Ö.; Ayan, A.; Atak, Ç. Enzymatic and non-enzymatic comparison of two different industrial tomato (Solanum lycopersicum) varieties against drought stress. Bot. Stud. 2017, 58, 32. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, H.; Tadayon, M.R.; Nadeem, M.; Cheema, M.; Razmjoo, J. Proline-mediated changes in antioxidant enzymatic activities and the physiology of sugar beet under drought stress. Acta Physiol. Plant. 2019, 41, 23. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Hu, H.; Hu, J.; Xiang, M.; Yang, Q. Comparative proteomics analysis of the responses to selenium in selenium-enriched alfalfa (Medicago sativa L.) leaves. Plant Physiol. Biochem. 2021, 165, 265–273. [Google Scholar] [CrossRef]
- Cunha, M.L.O.; de Oliveira, L.C.A.; Silva, V.M.; Montanha, G.S.; Dos Reis, A.R. Selenium increases photosynthetic capacity, daidzein biosynthesis, nodulation and yield of peanuts plants (Arachis hypogaea L.). Plant Physiol. Biochem. 2022, 190, 231–239. [Google Scholar] [CrossRef]
- Rady, M.O.; Semida, W.M.; Abd El-Mageed, T.A.; Howladar, S.M.; Shaaban, A. Foliage applied selenium improves photosynthetic efficiency, antioxidant potential and wheat productivity under drought stress. Int. J. Agric. Biol 2020, 24, 1293–1300. [Google Scholar]
- Jalil, S.U.; Ansari, M.I. Nanoparticles and abiotic stress tolerance in plants: Synthesis, action, and signaling mechanisms. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 549–561. [Google Scholar]
- Sharma, S.; Singh, V.K.; Kumar, A.; Mallubhotla, S. Effect of nanoparticles on oxidative damage and antioxidant defense system in plants. In Molecular Plant Abiotic Stress: Biology and Biotechnology; Wiley Online Library: Hoboken, NJ, USA, 2019; pp. 315–333. [Google Scholar]
- Naing, A.H.; Kim, C.K. Abiotic stress-induced anthocyanins in plants: Their role in tolerance to abiotic stresses. Physiol. Plant. 2021, 172, 1711–1723. [Google Scholar] [CrossRef]
- Sardar, R.; Ahmed, S.; Shah, A.A.; Yasin, N.A. Selenium nanoparticles reduced cadmium uptake, regulated nutritional homeostasis and antioxidative system in Coriandrum sativum grown in cadmium toxic conditions. Chemosphere 2022, 287, 132332. [Google Scholar] [CrossRef]
- Moaaz Ali, M.; Javed, T.; Mauro, R.P.; Shabbir, R.; Afzal, I.; Yousef, A.F. Effect of seed priming with potassium nitrate on the performance of tomato. Agriculture 2020, 10, 498. [Google Scholar] [CrossRef]
- Dewis, J.; Freitas, F. Physical and Chemical Methods of Soil and Water Analysis; FAO Soils Bulletins series 10; Food and Agriculture Organization of the United Nations: Rome, Italy, 1970. [Google Scholar]
- Capra, A.; Consoli, S.; Scicolone, B. Deficit irrigation: Theory and Practice. In Agricultural Irrigation Research Progress; Nova Science Publishers: New York, NY, USA, 2008; pp. 53–82. [Google Scholar]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A.J.P.S. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of Catalases and Peroxidases; Elsevier: Amsterdam, The Netherlands, 1955. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [Green Version]
- Kirk, J.T.O.; Allen, R.L. Dependence of chloroplast pigment synthesis on protein synthesis: Effect of actidione. Biochem. Biophys. Res. Commun. 1965, 21, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Solomon, A.; Golubowicz, S.; Yablowicz, Z.; Grossman, S.; Bergman, M.; Gottlieb, H.E.; Altman, A.; Kerem, Z.; Flaishman, M.A. Antioxidant activities and anthocyanin content of fresh fruits of common fig (Ficus carica L.). J. Agric. Food Chem. 2006, 54, 7717–7723. [Google Scholar] [CrossRef]
- Mattila, P.; Astola, J.; Kumpulainen, J. Determination of flavonoids in plant material by HPLC with diode-array and electro-array detections. J. Agric. Food Chem. 2000, 48, 5834–5841. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Foster, J.G.; Hess, J.L. Responses of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol. 1980, 66, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Miyake, C.; Asada, K. Thylakoid-bound ascorbate peroxidase in spinach chloroplasts and photoreduction of its primary oxidation product monodehydroascorbate radicals in thylakoids. Plant Cell Physiol. 1992, 33, 541–553. [Google Scholar]
- Baker, J.L., Jr. The effectiveness of alpha-tocopherol (vitamin E) in reducing the incidence of spherical contracture around breast implants. Plast. Reconstr. Surg. 1981, 68, 696–698. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variation Source | a df | SFW | RFW | PH | TC | Carotenoids | Ascorbic Acid | α-Tocopherols |
|---|---|---|---|---|---|---|---|---|
| Water Stress (WS) | 1 | 4788.033 b*** (0.000) | 375.948 *** (0.000) | 533.408 *** (0.000) | 2.616 *** (0.000) | 13.912 *** (0.000) | 4966.345 *** (0.000) | 421.12 *** (0.000) |
| Se-NPs Seed Priming (SSP) | 4 | 421.033 *** (0.000) | 37.052 *** (0.000) | 64.122 *** (0.000) | 0.151 *** (0.000) | 0.449 *** (0.000) | 602.8 *** (0.000) | 22.418 *** (0.000) |
| WS X SSP | 4 | 10.033 ns (0.120) | 0.692 ns (0.464) | 0.464 ns (0.537) | 0.004 * (0.021) | 0.006 ns (0.263) | 63.553 *** (0.000) | 0.722 ns (0.162) |
| Error | 20 | 4.800 | 0.742 | 0.577 | 0.001 | 0.004 | 5.566 | 0.394 |
| Variation Source | df | Flavonoids | H2O2 | MDA | SOD | CAT | GST | APX |
| Water Stress (WS) | 1 | 95.551 *** (0.000) | 12,120.3 *** (0.000) | 811.2 *** (0.000) | 8433.633 *** (0.000) | 5266.871 *** (0.000) | 1954.392 *** (0.000) | 574.918 *** (0.000) |
| Se-NPs Seed Priming (SSP) | 4 | 8.425 *** (0.000) | 622.133 *** (0.000) | 64.466 *** (0.000) | 816.783 *** (0.000) | 186.366 *** (0.000) | 57.153 *** (0.000) | 28.238 ns (0.057) |
| WS X SSP | 4 | 0.717 ** (0.013) | 201.8 ***(0.000) | 22.533 *** (0.000) | 183.716 *** (0.000) | 36.416 *** (0.000) | 2.099 ns (0.079) | 8.535 ns (0.522) |
| Error | 20 | 0.105 | 7.666 | 2.633 | 9.366 | 2.991 | 0.855 | 0.001 |
| Variation Source | df | GR | MDHAR | DHAR | GSH | GSSG | Proline | Anthocyanins |
| Water Stress (WS) | 1 | 2043.030 *** (0.000) | 1560.984 *** (0.000) | 5826.920 *** (0.000) | 5096.33 *** (0.000) | 594.965 *** (0.000) | 459.425 *** (0.000) | 0.633 *** (0.000) |
| Se-NPs Seed Priming (SSP) | 4 | 48.010 *** (0.000) | 11.872 *** (0.000) | 169.671 *** (0.000) | 492.283 ***(0.000) | 42.492 ***(0.000) | 20.122 *** (0.000) | 0.019 *** (0.000) |
| WS X SSP | 4 | 8.617 *** (0.000) | 2.417 ns (0.339) | 20.401 *** (0.000) | 67.783 *** (0.000) | 6.362 ** (0.001) | 5.468 *** (0.000) | 0.006 *** (0.000) |
| Error | 20 | 0.763 | 2.033 | 2.528 | 6.133 | 0.929 | 0.272 | 0.001 |
| Variables | TC | Carotenoids | Ascorbic Acid | Toco | Flavonoids | H2O2 | MDA | SOD | CAT | GST | APX | GR | MDHAR | DHAR | GSH | GSSG | Proline | Ant |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TC | 1 | |||||||||||||||||
| Carotenoids | −0.517 * | 1 | ||||||||||||||||
| Ascorbic acid | −0.512 * | 0.989 * | 1 | |||||||||||||||
| Toco | −0.52 * | 0.989 * | 0.993 * | 1 | ||||||||||||||
| Flavonoids | −0.517 * | 0.995 * | 0.991 * | 0.985 * | 1 | |||||||||||||
| H2O2 | −0.983 * | 0.532 * | 0.532 * | 0.537 * | 0.531 * | 1 | ||||||||||||
| MDA | −0.936 * | 0.591 * | 0.573 * | 0.577 * | 0.59 * | 0.924 * | 1 | |||||||||||
| SOD | −0.492 * | 0.966 * | 0.972 * | 0.966 * | 0.973 * | 0.511 * | 0.539 * | 1 | ||||||||||
| CAT | −0.52 * | 0.97 * | 0.98 * | 0.97 * | 0.978 * | 0.54 * | 0.57 * | 0.972 * | 1 | |||||||||
| GST | −0.518 * | 0.989 * | 0.995 * | 0.991 * | 0.988 * | 0.537 * | 0.581 * | 0.97 * | 0.976 * | 1 | ||||||||
| APX | −0.522 * | 0.812 * | 0.794 * | 0.798 * | 0.799 * | 0.508 * | 0.595 * | 0.788 * | 0.769 * | 0.793 * | 1 | |||||||
| GR | −0.528 * | 0.973 * | 0.972 * | 0.955 * | 0.982 * | 0.547 * | 0.575 * | 0.973 * | 0.978 * | 0.967 * | 0.774 * | 1 | ||||||
| MDHAR | −0.513 * | 0.995 * | 0.99 * | 0.988 * | 0.989 * | 0.524 * | 0.581 * | 0.964 * | 0.969 * | 0.989 * | 0.799 * | 0.965 * | 1 | |||||
| DHAR | −0.521 * | 0.989 * | 0.977 * | 0.974 * | 0.988 * | 0.528 * | 0.589 * | 0.972 * | 0.957 * | 0.979 * | 0.808 * | 0.972 * | 0.986 * | 1 | ||||
| GSH | −0.519 * | 0.989 * | 0.982 * | 0.977 * | 0.988 * | 0.525 * | 0.589 * | 0.962 * | 0.968 * | 0.985 * | 0.795 * | 0.967 * | 0.99 * | 0.986 * | 1 | |||
| GSSG | −0.51 * | 0.967 * | 0.985 * | 0.983 * | 0.969 * | 0.534 * | 0.571 * | 0.951 * | 0.957 * | 0.976 * | 0.782 * | 0.942 * | 0.969 * | 0.952 * | 0.955 * | 1 | ||
| Proline | −0.513 * | 0.985 * | 0.992 * | 0.981 * | 0.99 * | 0.531 * | 0.573 * | 0.982 * | 0.976 * | 0.987 * | 0.799 * | 0.979 * | 0.986 * | 0.987 * | 0.98 * | 0.973 * | 1 | |
| Ant | −0.521 * | 0.969 * | 0.978 * | 0.965 * | 0.977 * | 0.525 * | 0.574 * | 0.961 * | 0.971 * | 0.971 * | 0.839 * | 0.969 * | 0.975 * | 0.97 * | 0.974 * | 0.954 * | 0.981 * | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishtiaq, M.; Mazhar, M.W.; Maqbool, M.; Hussain, T.; Hussain, S.A.; Casini, R.; Abd-ElGawad, A.M.; Elansary, H.O. Seed Priming with the Selenium Nanoparticles Maintains the Redox Status in the Water Stressed Tomato Plants by Modulating the Antioxidant Defense Enzymes. Plants 2023, 12, 1556. https://doi.org/10.3390/plants12071556
Ishtiaq M, Mazhar MW, Maqbool M, Hussain T, Hussain SA, Casini R, Abd-ElGawad AM, Elansary HO. Seed Priming with the Selenium Nanoparticles Maintains the Redox Status in the Water Stressed Tomato Plants by Modulating the Antioxidant Defense Enzymes. Plants. 2023; 12(7):1556. https://doi.org/10.3390/plants12071556
Chicago/Turabian StyleIshtiaq, Muhammad, Muhammad Waqas Mazhar, Mehwish Maqbool, Tanveer Hussain, Syed Atiq Hussain, Ryan Casini, Ahmed M. Abd-ElGawad, and Hosam O. Elansary. 2023. "Seed Priming with the Selenium Nanoparticles Maintains the Redox Status in the Water Stressed Tomato Plants by Modulating the Antioxidant Defense Enzymes" Plants 12, no. 7: 1556. https://doi.org/10.3390/plants12071556