
Transcriptional Profile of Soybean Seeds with Contrasting Seed Coat Color
 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
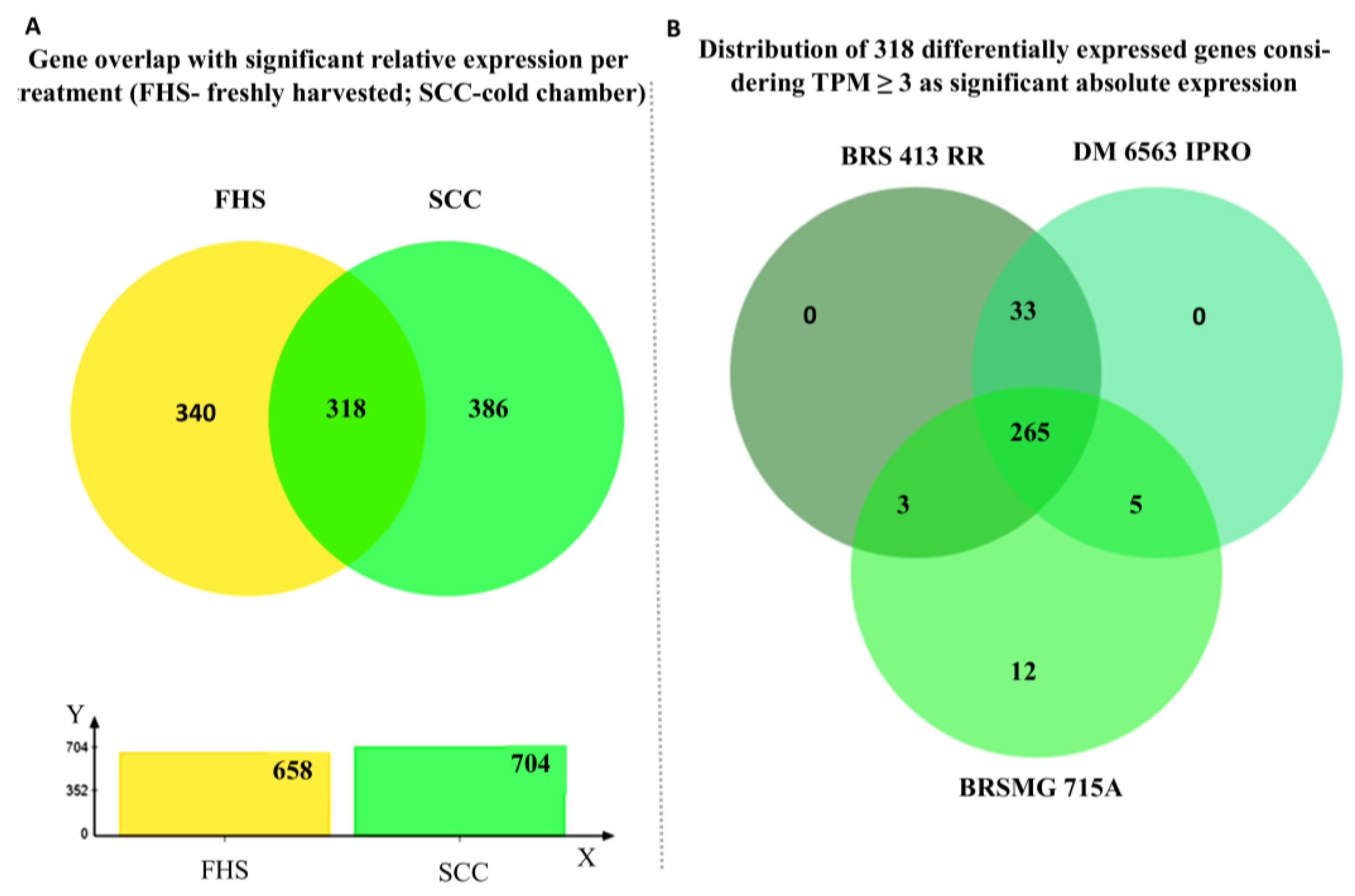
2.1. Differential Expression
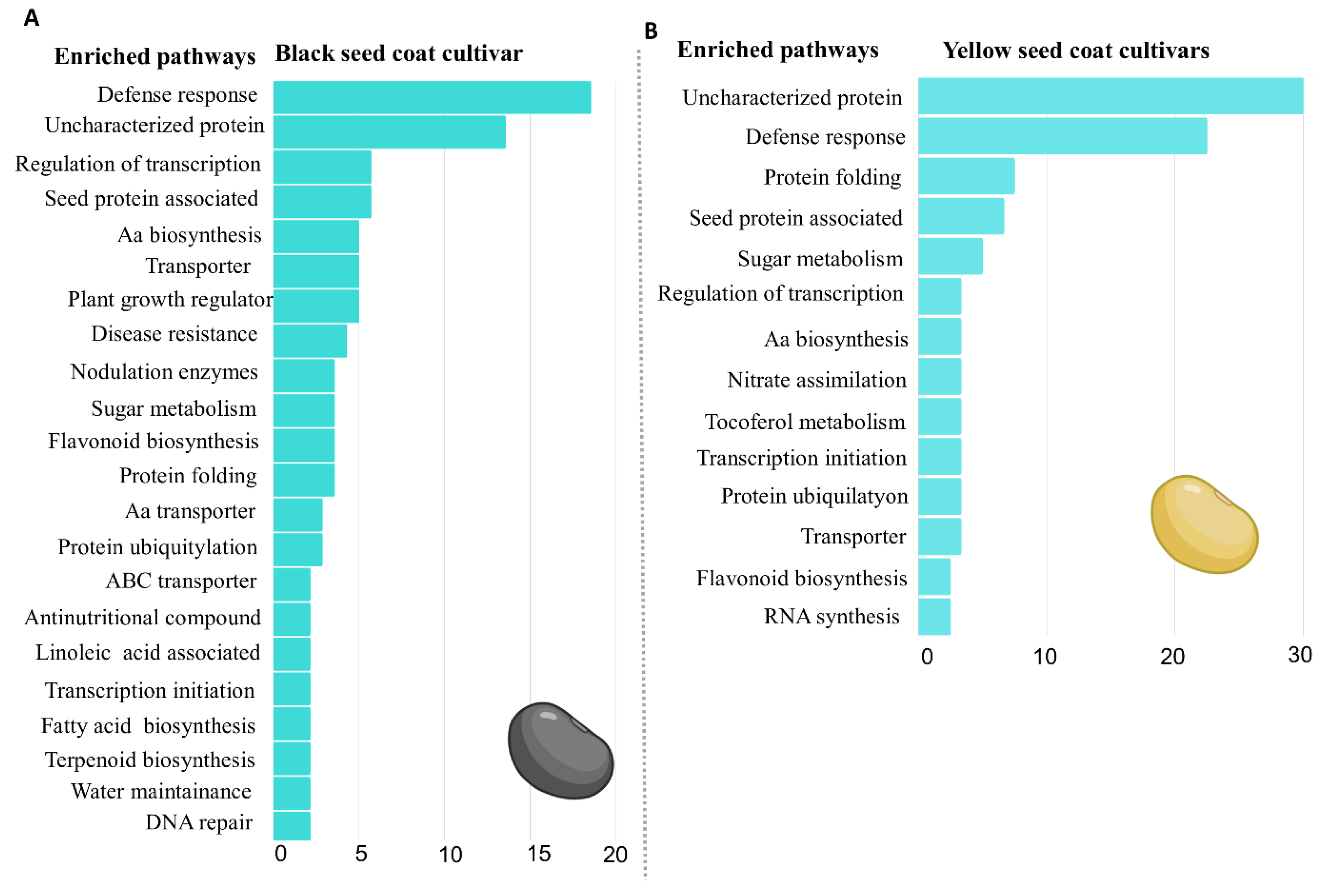
2.2. Enriched Metabolic Pathways
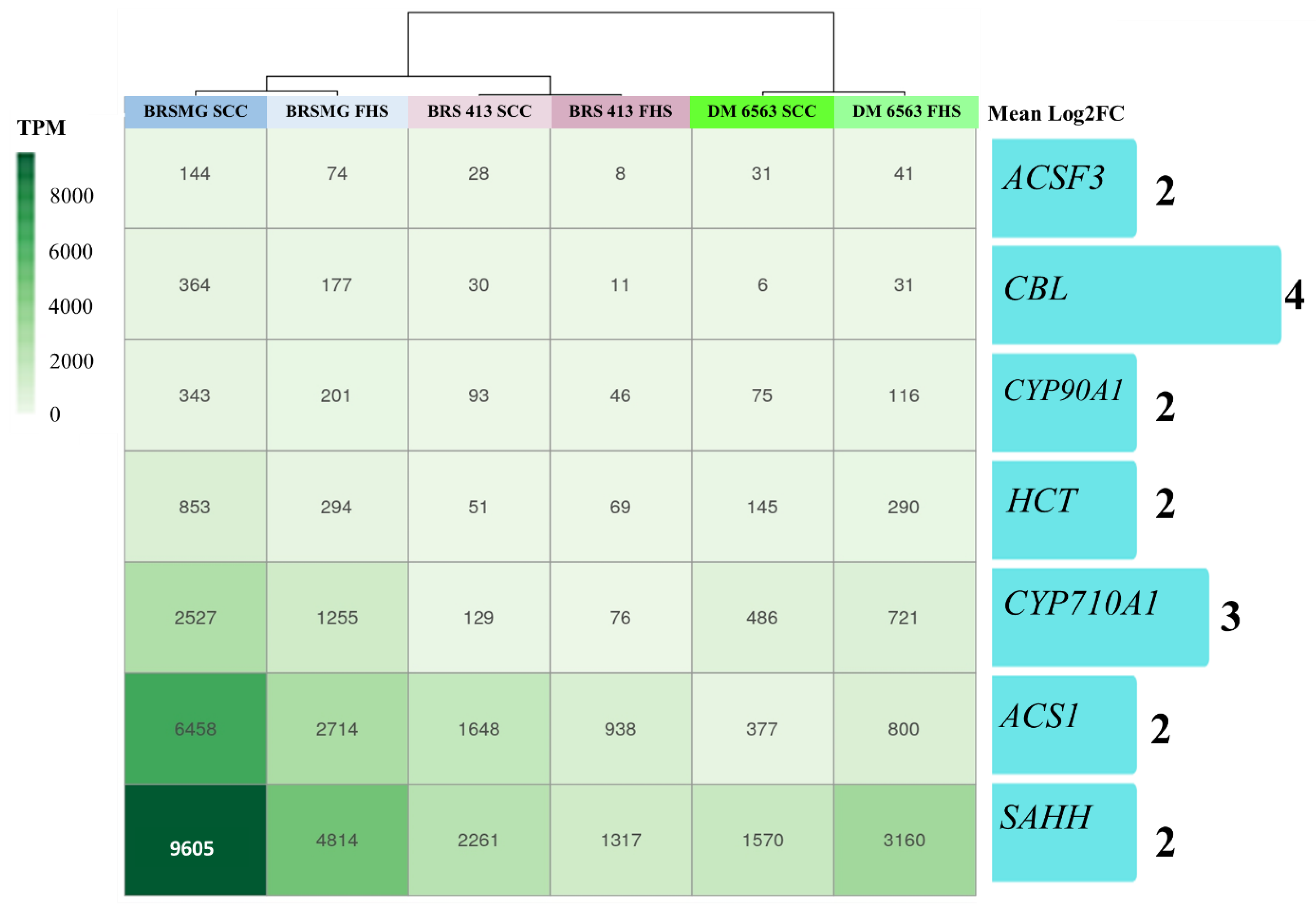
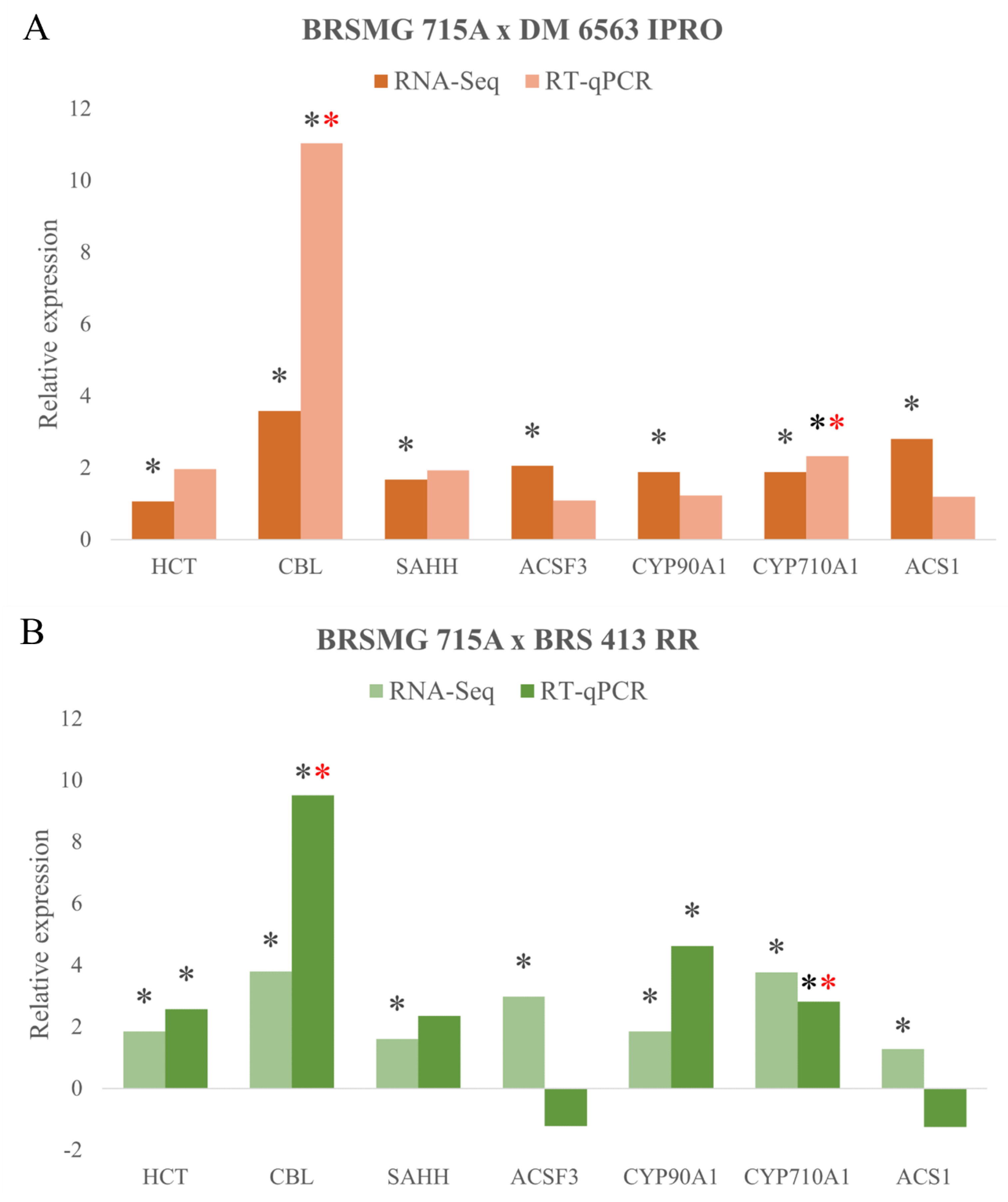
2.3. Relative and Absolute Expression
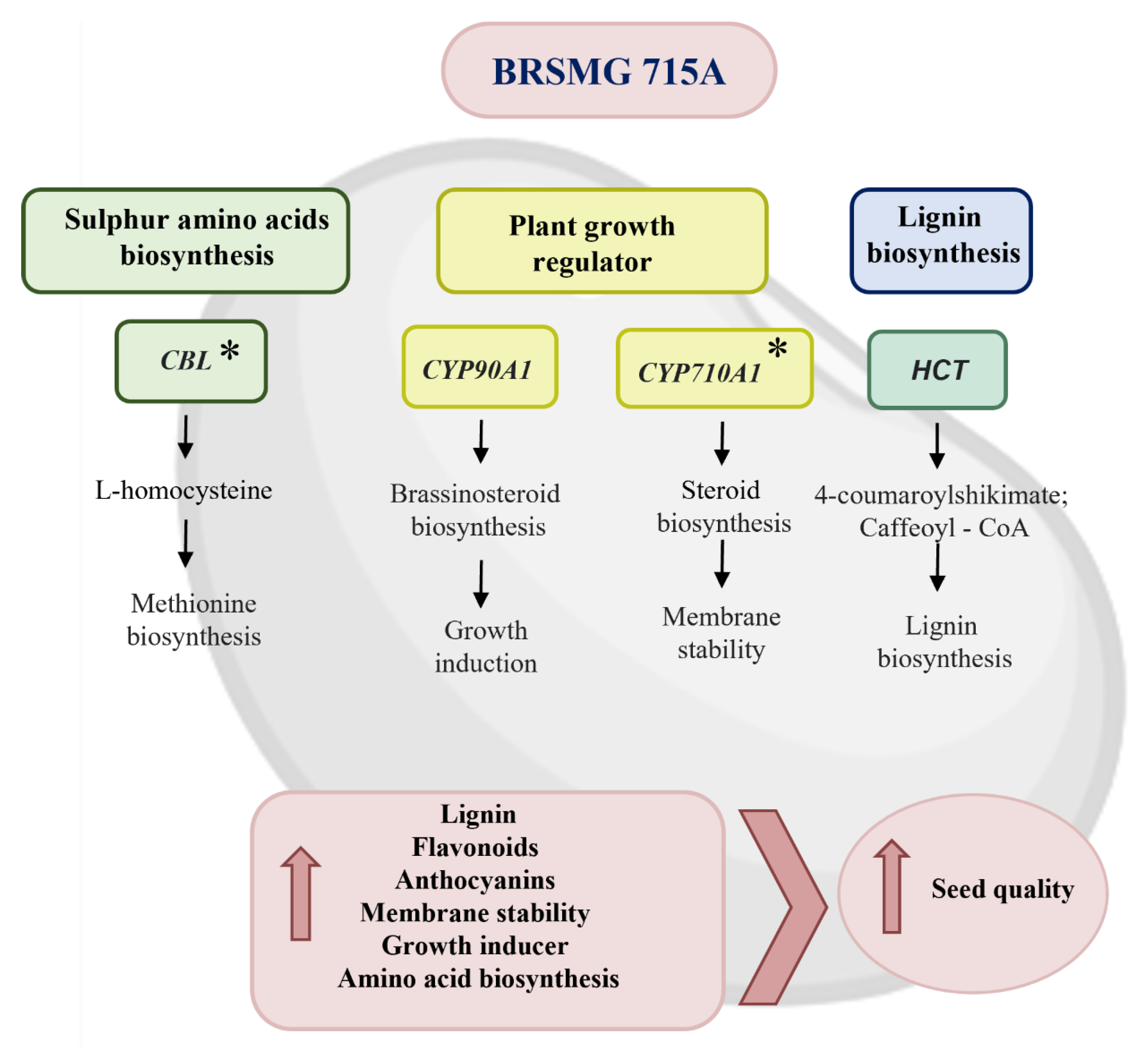
2.3.1. Genes Involved in Amino Acid Biosynthesis
2.3.2. Genes Involved in Plant Growth Regulators
2.3.3. Genes Involved in Fatty Acid Biosynthesis
2.3.4. Genes Involved in Lignin Biosynthesis
2.4. Evaluation of the Number of Copies and Orthologs in Arabidopsis
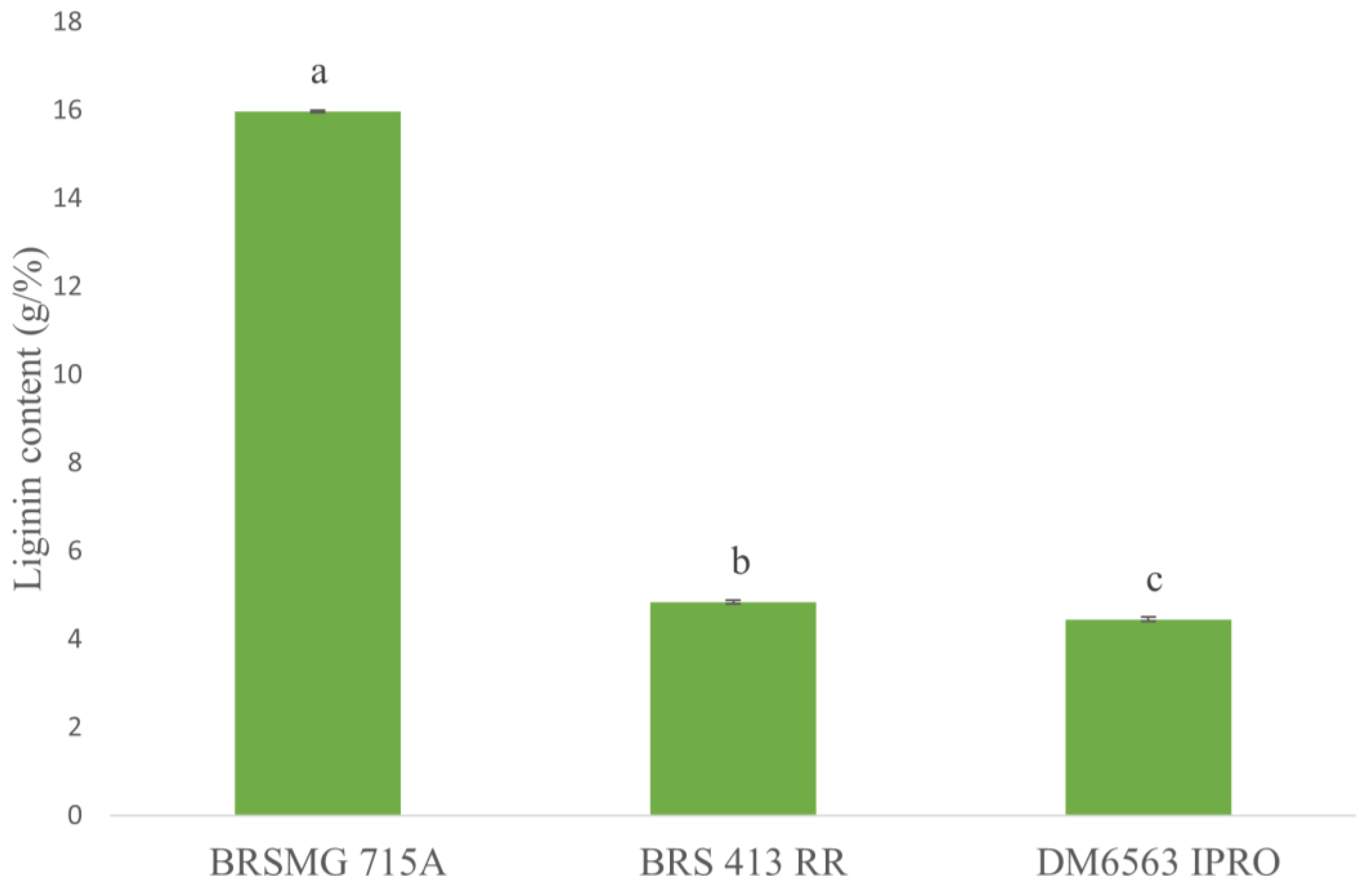
2.5. Assessing Lignin Content in the Seed Coat
3. Materials and Methods
3.1. RNA-Seq Analysis
3.2. Bioinformatics Analysis
3.3. RT-qPCR Analysis
3.4. Lignin Content of Soybean Seed Coat
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medic, J.; Atkinson, C.; Hurburgh, C.R. Current knowledge in soybean composition. J. Am. Oil Chem. Soc. 2014, 91, 363–384. [Google Scholar] [CrossRef]
- Sharma, S.; Kaur, M.; Goyal, R.; Gill, B.S. Physical characteristics and nutritional composition of some new soybean (Glycine max (L.) Merrill) genotypes. J. Food Sci. Technol. 2011, 51, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and Biotic Stresses in Soybean Production; Academic Press: Cambridge, MA, USA, 2016; pp. 1–26. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#rankings/countries_by_commodity (accessed on 11 January 2022).
- Brazil: Oilseeds and Products Update|USDA Foreign Agricultural Service. Available online: https://www.fas.usda.gov/data/brazil-oilseeds-and-products-update-27 (accessed on 4 November 2021).
- Gwinner, R.; Alemu Setotaw, T.; Pasqual, M.; Dos Santos, J.B.; Zuffo, A.M.; Zambiazzi, E.V.; Bruzi, A.T. Genetic diversity in Brazilian soybean germplasm. Crop. Breed. Appl. Biotechnol. 2017, 17, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Liu, Z.; Hong, H.; Ma, Y.; Tian, L.; Li, X.; Li, Y.-H.; Guan, R.; Guo, Y.; Qiu, L.-J. Identification and validation of loci governing seed coat color by combining association mapping and bulk segregation analysis in soybean. PLoS ONE 2016, 11, e0159064. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Chang, S.K.C. Antioxidant capacity of seed coat, dehulled bean, and whole black soybeans in relation to their distributions of total phenolics, phenolic acids, anthocyanins, and isoflavones. J. Agric. Food Chem. 2008, 56, 8365–8373. [Google Scholar] [CrossRef]
- Bahry, C.A.; Dos Santos Acunha, T.; Fernando, J.A.; Chaves, F.C.; Nardino, M.; Zimmer, P.D. Chemical composition and structural characterization of contrasting colors of soybean seed coats. Semin. Agrar. 2015, 36, 1913–1926. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Cho, K.M. Changes occurring in compositional components of black soybeans maintained at room temperature for different storage periods. Food Chem. 2012, 131, 161–169. [Google Scholar] [CrossRef]
- Radix Astadi, I.; Paice, A.G. Black Soybean (Glycine max L: Merril) Seeds’ Antioxidant Capacity. In Nuts and Seeds in Health and Disease Prevention; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 229–236. [Google Scholar] [CrossRef]
- Wu, T.; Guo, X.; Zhang, M.; Yang, L.; Liu, R.; Yin, J. Anthocyanins in black rice, soybean and purple corn increase fecal butyric acid and prevent liver inflammation in high fat diet-induced obese mice. Food Funct. 2017, 8, 3178–3186. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. A critical review on polyphenols and health benefits of black soybeans. Nutrients 2017, 9, 455. [Google Scholar] [CrossRef] [Green Version]
- Menezes, M.D.; von Pinho, É.V.D.R.; José, S.C.B.R.; Baldoni, A.; Mendes, F.F. Aspectos químicos e estruturais da qualidade fisiológica de sementes de soja. Pesqui. Agropecu. Bras. 2009, 44, 1716–1723. [Google Scholar] [CrossRef]
- Carvalho, E.R.; Oliveira, J.A.; Reis, L.V.; Ferreira, T.F. Foliar manganese in the health and lignin quality of conventional and glyphosateresistant soybean seeds. Rev. Ciênc. Agron. 2015, 46, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Mertz-Henning, L.M.; Nagashima, A.I.; Krzyzanowski, F.C.; Binneck, E.; Henning, F.A. Relative quantification of gene expression levels associated with lignin biosynthesis in soybean seed coat. Seed Sci. Technol. 2015, 43, 445–455. [Google Scholar] [CrossRef]
- Abati, J.; Zucareli, C.; Brzezinski, C.R.; Krzyzanowski, F.C.; França-Neto, J.D.B.; Henning, F.A. Metabolites of the phenylpropanoid pathway and physiological quality of soybean seeds in storage. J. Seed Sci. 2021, 43. [Google Scholar] [CrossRef]
- Abati, J.; Zucareli, C.; Brzezinski, C.R.; Lopes, I.D.O.N.; Krzyzanowski, F.C.; Moraes, L.A.C.; Henning, F.A. Water absorption and storage tolerance of soybean seeds with contrasting seed coat characteristics. Acta Sci. Agron. 2022, 44. [Google Scholar] [CrossRef]
- Kuchlan, M.K.; Kuchlan, P.; Onkar, M.; Ramesh, A.; Husain, S.M. Influence of seed coat compactness around cotyledons, protein and mineral composition on mechanical strength of soybean [Glycine max (L.) Merrill] seed coat. Legume Res. 2018, 41, 246–252. [Google Scholar]
- Madureira, A.; Molinari, M.D.C.; Marin, S.R.R.; Pagliarini, R.F.; Henning, F.A.; Nepomuceno, A.L.; Finatto, T.; Mertz-Henning, L.M. Correlations between lignin content and related genes, weathering deterioration, and soybean seed quality at pre-harvest. J. Crop. Sci. Biotechnol. 2023, 1–9. [Google Scholar] [CrossRef]
- Moura, J.C.M.S.; Bonine, C.A.V.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef]
- Kuchlan, M.K.; Dadlani, M.; Samuel, D.V.K. Seed coat properties and longevity of soybean seeds. J. New Seeds 2020, 11, 239–249. [Google Scholar] [CrossRef]
- Liu, J.; Qin, W.-T.; Wu, H.-J.; Yang, C.-Q.; Deng, J.-C.; Iqbal, N.; Liu, W.-G.; Du, J.-B.; Shu, K.; Yang, F.; et al. Metabolism variation and better storability of dark- versus light-coloured soybean (Glycine max L. Merr.) seeds. Food Chem. 2017, 223, 104–113. [Google Scholar] [CrossRef]
- Adsul, A.T.; Chimote, V.P.; Deshmukh, M.P. Inheritance of Seed Longevity and Its Association with Other Seed-Related Traits in Soybean (Glycine max). Agric. Res. 2018, 7, 105–111. [Google Scholar] [CrossRef]
- Reese, P.F.; Boerma, H.R. Additional genes for green seed coat in soybean. J. Hered. 1989, 80, 86–88. [Google Scholar] [CrossRef]
- Tuteja, J.H.; Zabala, G.; Varala, K.; Hudson, M.; Vodkin, L.O. Endogenous, tissue-specific short interfering RNAs silence the chalcone synthase gene family in glycine max seed coatsWOA. Plant Cell 2009, 21, 3063–3077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre, M.; Kiegle, E.; Leo, G.; Ezquer, I. Carbohydrate reserves and seed development: An overview. Plant Reprod. 2018, 31, 263–290. [Google Scholar] [CrossRef] [PubMed]
- Tayade, R.; Kulkarni, K.P.; Jo, H.; Song, J.T.; Lee, J.D. Insight into the prospects for the improvement of seed starch in legume—A review. Front. Plant Sci. 2019, 10, 1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, Y.; Yan, Y.; Peng, H.; Long, Y.; Zhang, Y.; Jiang, Z.; Liu, P.; Zou, C.; Peng, H.; et al. Transcriptome sequencing analysis of maize embryonic callus during early redifferentiation. BMC Genom. 2019, 20, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.F.; Chang, T.Y.; Chiang, S.F.; Wang, W.D.; Charng, Y.Y.; Chiou, T.J. Arabidopsis inositol pentakisphosphate 2-kinase, AtIPK1, is required for growth and modulates phosphate homeostasis at the transcriptional level. Plant J. 2014, 80, 503–515. [Google Scholar] [CrossRef]
- Basak, N.; Krishnan, V.; Pandey, V.; Punjabi, M.; Hada, A.; Marathe, A.; Jolly, M.; Palaka, B.K.; Ampasala, D.R.; Sachdev, A. Expression profiling and in silico homology modeling of Inositol penta kis phosphate 2-kinase, a potential candidate gene for low phytate trait in soybean. 3 Biotech 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Azizkhani, N.; Mirzaei, S.; Torkzadeh-Mahani, M. Genome-wide identification and characterization of legume T2 Ribonuclease gene family and analysis of GmaRNS9, a soybean T2 Ribonuclease gene, function in nodulation. 3 Biotech 2021, 11, 1–21. [Google Scholar] [CrossRef]
- Belkhadir, Y.; Yang, L.; Hetzel, J.; Dangl, J.L.; Chory, J. The growth–defense pivot: Crisis management in plants mediated by LRR-RK surface receptors. Trends Biochem. Sci. 2014, 39, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, T.; Hatano, K.I.; Miyauchi, Y.; Suwa, Y.I.; Sawano, Y.; Tanokura, M. A secreted protein with plant-specific cysteine-rich motif functions as a mannose-binding lectin that exhibits antifungal activity. Plant Physiol. 2014, 166, 766–778. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, H.B.; Jez, J.M. Review: The promise and limits for enhancing sulfur-containing amino acid content of soybean seed. Plant Sci. 2018, 272, 14–21. [Google Scholar] [CrossRef]
- Raksha, B.R.; Siva, R.; Vino, S.; Babu, S. Spatio-varietal differences in stigmasterol biosynthesis in tomato and overexpression of a sterol desaturase gene for enhanced stigmasterol production. Vitr. Cell. Dev. Biol.-Plant 2016, 52, 571–579. [Google Scholar] [CrossRef]
- Singer, W.M.; Zhang, B.; Mian, M.A.R.; Huang, H. Soybean Amino Acids in Health, Genetics, and Evaluation. Soybean Hum. Consum. Anim. Feed 2019. [Google Scholar] [CrossRef] [Green Version]
- Malle, S.; Eskandari, M.; Morrison, M.; Belzile, F. Genome-wide association identifies several QTLs controlling cysteine and methionine content in soybean seed including some promising candidate genes. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Bai, Z.; Qi, T.; Liu, Y.; Wu, Z.; Ma, L.; Liu, W.; Cao, Y.; Bao, Y.; Fu, C. Alteration of S-adenosylhomocysteine levels affects lignin biosynthesis in switchgrass. Plant Biotechnol. J. 2018, 16, 2016–2026. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Ma, W.; Hu, D.; Zhang, X.; Yuan, W.; He, X.; Kan, G.; Yu, D. QTL Mapping for Protein and Sulfur-Containing Amino Acid Contents Using a High-Density Bin-Map in Soybean (Glycine max L. Merr.). J. Agric. Food Chem. 2019, 67, 12313–12321. [Google Scholar] [CrossRef]
- Maimann, S.; Wagner, C.; Kreft, O.; Zeh, M.; Willmitzer, L.; Höfgen, R.; Hesse, H. Transgenic potato plants reveal the indispensable role of cystathionine β-lyase in plant growth and development. Plant J. 2000, 23, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Yang, W.; Zhang, X.; Peng, T.; Zou, Y.; Zhang, T.; Wang, H.; Liu, X.; Tao, L. Cystathionine beta-lyase is crucial for embryo patterning and the maintenance of root stem cell niche in Arabidopsis. Plant J. 2019, 99, 536–555. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, X.; Zuo, L.; Wang, H.; Yu, D. Identification and functional characterization of the sulfate transporter gene GmSULTR1;2b in soybean. BMC Genom. 2016, 17, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Chiba, Y.; Hirai, M.Y. Metabolism and Regulatory Functions of O-Acetylserine, S-Adenosylmethionine, Homocysteine, and Serine in Plant Development and Environmental Responses. Front. Plant Sci. 2021, 12, 593. [Google Scholar] [CrossRef]
- Pattyn, J.; Vaughan-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: A complex multilevel control circuitry. New Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Koda, Y.; Zheng, S.H.; Yuasa, T.; Iwaya-Inoue, M. Regulation of soybean seed germination through ethylene production in response to reactive oxygen species. Ann. Bot. 2013, 111, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Ahammed, G.J.; Gantait, S.; Mitra, M.; Yang, Y.; Li, X. Role of ethylene crosstalk in seed germination and early seedling development: A review. Plant Physiol. Biochem. 2020, 151, 124–131. [Google Scholar] [CrossRef]
- Ohnishi, T.; Godza, B.; Watanabe, B.; Fujioka, S.; Hategan, L.; Ide, K.; Shibata, K.; Yokota, T.; Szekeres, M.; Mizutani, M. CYP90A1/CPD, a brassinosteroid biosynthetic cytochrome P450 of Arabidopsis, catalyzes C-3 oxidation. J. Biol. Chem. 2012, 287, 31551–31560. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Dong, N.; Niu, M.; Zhang, X.; Li, L.; Liu, J.; Liu, B.; Tong, H. Brassinosteroid-regulated plant growth and development and gene expression in soybean. Crop. J. 2019, 7, 411–418. [Google Scholar] [CrossRef]
- Wang, M.; Xu, X.; Zhang, X.; Sun, S.; Wu, C.; Hou, W.; Wang, Q.; Han, T. Functional analysis of GmCPDs and investigation of their roles in flowering. PLoS ONE 2015, 10, e0118476. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Wang, Z.; Guo, X.; Liu, F.; Jiang, J.; Liu, G. BpMADS12 gene role in lignin biosynthesis of Betula platyphylla Suk by transcriptome analysis. J. For. Res. 2016, 27, 1111–1120. [Google Scholar] [CrossRef]
- Fan, C.; Yu, H.; Qin, S.; Li, Y.; Alam, A.; Xu, C.; Fan, D.; Zhang, Q.; Wang, Y.; Zhu, W.; et al. Brassinosteroid overproduction improves lignocellulose quantity and quality to maximize bioethanol yield under green-like biomass process in transgenic poplar. Biotechnol. Biofuels 2020, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant sterols: Diversity, biosynthesis, and physiological functions. Biochemistry 2016, 81, 819–834. [Google Scholar] [CrossRef]
- Aboobucker, S.I.; Suza, W.P. Why do plants convert sitosterol to stigmasterol? Front. Plant Sci. 2019, 10, 354. [Google Scholar] [CrossRef] [Green Version]
- Griebel, T.; Zeier, J. A role for β-sitosterol to stigmasterol conversion in plant-pathogen interactions. Plant J. 2010, 63, 254–268. [Google Scholar] [CrossRef]
- Wang, K.; Senthil-Kumar, M.; Ryu, C.M.; Kang, L.; Mysore, K.S. Phytosterols play a key role in plant innate immunity against bacterial pathogens by regulating nutrient efflux into the apoplast. Plant Physiol. 2012, 158, 1789–1802. [Google Scholar] [CrossRef] [Green Version]
- Cabianca, A.; Müller, L.; Pawlowski, K.; Dahlin, P. Changes in the plant β-sitosterol/stigmasterol ratio caused by the plant parasitic nematode meloidogyne incognita. Plants 2021, 10, 292. [Google Scholar] [CrossRef]
- Suo, X.; Xu, F.; Tan, K.; Huang, L.; Bao, C.; Luo, M. Functions of phytosterols in seed development of upland cotton (Gossypium hirsutum L.). Ind. Crops Prod. 2021, 170, 113802. [Google Scholar] [CrossRef]
- Song, J.; Sun, X.; Zhang, K.; Liu, S.; Wang, J.; Yang, C.; Jiang, S.; Siyal, M.; Li, X.; Qi, Z.; et al. Identification of QTL and genes for pod number in soybean by linkage analysis and genome-wide association studies. Mol. Breed. 2020, 40, 1–14. [Google Scholar] [CrossRef]
- Ariyoshi, Y.; Itoyama, H.; Nakagawa, A.C.S.; Ario, N.; Kondo, Y.; Tomita, Y.; Tanaka, S.; Nakashima, M.; Tomioka, K.; Iwaya-Inoue, M.; et al. Regulation of brassinosteroid on pod growth through cell hypertrophy in soybean (Glycine max (L.) Merr.). Plant Growth Regul. 2016, 80, 391–395. [Google Scholar] [CrossRef]
- Chen, H.; Kim, H.U.; Weng, H.; Browse, J. Malonyl-CAa synthetase, encoded by ACYL activating ENZYME13, is essential for growth and development of Arabidopsis. Plant Cell 2011, 23, 2247–2262. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi, M.; Dhaubhadel, S. Proteomic insights into synthesis of isoflavonoids in soybean seeds. Proteomics 2015, 15, 1646–1657. [Google Scholar] [CrossRef]
- Shadle, G.; Chen, F.; Srinivasa Reddy, M.S.; Jackson, L.; Nakashima, J.; Dixon, R.A. Down-regulation of hydroxycinnamoyl CoA: Shikimate hydroxycinnamoyl transferase in transgenic alfalfa affects lignification, development and forage quality. Phytochemistry 2007, 68, 1521–1529. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Bhattarai, K.; Rajasekar, S.; Dixon, R.A.; Monteros, M.J. Agronomic Performance and Lignin Content of HCT Down-Regulated Alfalfa (Medicago sativa L.). Bioenergy Res. 2018, 11, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Ha, C.M.; Fine, D.; Bhatia, A.; Rao, X.; Martin, M.Z.; Engle, N.L.; Wherritt, D.J.; Tschaplinski, T.J.; Sumner, L.W.; Dixon, R.A. Ectopic Defense Gene Expression Is Associated with Growth Defects in Medicago truncatula Lignin Pathway Mutants. Plant Physiol. 2019, 181, 63–84. [Google Scholar] [CrossRef] [Green Version]
- Serrani-Yarce, J.C.; Escamilla-Trevino, L.; Barros, J.; Gallego-Giraldo, L.; Pu, Y.; Ragauskas, A.; Dixon, R.A. Targeting hydroxycinnamoyl CoA: Shikimate hydroxycinnamoyl transferase for lignin modification in Brachypodium distachyon. Biotechnol. Biofuels 2021, 14, 1–17. [Google Scholar] [CrossRef]
- Huth, C.; Mertz-Henning, L.M.; Lopes, S.J.; Tabaldi, L.A.; Rossato, L.V.; Krzyzanowski, F.C.; Henning, F.A. Susceptibility to weathering damage and oxidative stress on soybean seeds with different lignin contents in the seed coat 1. J. Seed Sci. 2016, 38, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Bellaloui, N.; Mengistu, A.; Fisher, D.K.; Abel, C.A. Soybean seed composition constituents as affected by drought and phomopsis in phomopsis susceptible and resistant genotypes. J. Crop Improv. 2012, 26, 428–453. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Arnqvist, L.; Persson, M.; Jonsson, L.; Dutta, P.C.; Sitbon, F. Overexpression of CYP710A1 and CYP710A4 in transgenic Arabidopsis plants increases the level of stigmasterol at the expense of sitosterol. Planta 2008, 227, 309–317. [Google Scholar] [CrossRef]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of Hydroxycinnamoyl-Coenzyme A Shikimate/Quinate Hydroxycinnamoyltransferase Affects Phenylpropanoid Biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [Green Version]
- Eudes, A.; Pereira, J.H.; Yogiswara, S.; Wang, G.; Teixeira Benites, V.; Baidoo, E.E.; Lee, T.S.; Adams, P.; Keasling, J.; Loqué, D. Exploiting the substrate promiscuity of hydroxycinnamoyl-CoA: Shikimate hydroxycinnamoyl transferase to reduce lignin. Plant Cell Physiol. 2016, 57, 568–579. [Google Scholar] [CrossRef]
- Ciabotti, S.; Silva, A.C.B.B.; Juhasz, A.C.P.; Mendonça, C.D.; Tavano, O.L.; Mandarino, J.M.G.; Gonçalves, C.A.A. Chemical composition, protein profile, and isoflavones content in soybean genotypes with different seed coat colors. Int. Food Res. J. 2016, 23, 621–629. [Google Scholar]
- Chandra, S.; Talukdar, A.; Taak, Y.; Yadav, R.R.; Saini, M.; Sipani, N.S. Seed longevity studies in wild type, cultivated and inter-specific recombinant inbred lines (RILs) of soybean [Glycine max (L.) Merr.]. Genet. Resour. Crop. Evol. 2022, 69, 399–409. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Andrews, S. FastQC-A Quality Control Application for FastQ Files; Babraham Bioinformatics: Babraham, UK, 2010. [Google Scholar]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAM tools. Bioinformatics 2019, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Racine, J.S. RStudio: A Platform-Independent IDE for R and Sweave. J. Appl. Econ. 2012, 27, 167–172. [Google Scholar] [CrossRef]
- Molinari, M.; Mendonça, J.A.; Marin, D.R.; Mertz-henning, L.; Nepomuceno, A.L. Transcriptome analysis using RNA-Seq from experiments with and without biological replicates: A review. Rev. Ciênc. Agrár. 2021, 64, 1–13. [Google Scholar]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, R.G.; Cote, C. Mathematics of quantitative kinetic PCR and the application of standard curves. Nucleic Acids Res. 2003, 31, e93. [Google Scholar] [CrossRef] [PubMed]
- Gillman, J.D.; Tetlow, A.; Lee, J.D.; Shannon, J.G.; Bilyeu, K. Loss-of-function mutations affecting a specific Glycine max R2R3 MYB transcription factor result in brown hilum and brown seed coats. BMC Plant Biol. 2011, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Canteri, M.G.; Althaus, R.A.; das Virgens Filho, J.S.; Giglioti, E.A.; Godoy, C.V. SASM-AGRI-Sistema para Análise e Separação de Médias em Experimentos Agrícolas Pelos Métodos Scott-Knott, Tukey e Duncan; ScienceOpen, Inc.: Boston, MA, USA; Berlin, Germany; Budapest, Hungary, 2001. [Google Scholar]
- Moreira-Vilar, F.C.; Siqueira-Soares, R.D.C.; Finger-Teixeira, A.; De Oliveira, D.M.; Ferro, A.P.; Da Rocha, G.J.; Ferrarese, M.D.L.L.; Dos Santos, W.D.; Ferrarese-Filho, O. The Acetyl Bromide Method Is Faster, Simpler and Presents Best Recovery of Lignin in Different Herbaceous Tissues than Klason and Thioglycolic Acid Methods. PLoS ONE 2014, 9, e110000. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | PhytoMine Annotation | KEGG |
|---|---|---|
| Glyma.06G213600 | isoamylase/debranching enzyme | sugar metabolism |
| Glyma.06G043900 | cyclin A (CCNA) | seed development |
| Glyma.01G016700 | inositol-pentakisphosphate 2-kinase/ip5 2-kinase | phytic acid biosynthetic pathway |
| Glyma.02G064300 | ribonuclease t2 [ec:3.1.27.1] (e3.1.27.1) | nodule development |
| Glyma.02G293700 | N/A | uncharacterized protein |
| Glyma.06G267400 | leucine-rich repeat-containing protein | disease resistance |
| Glyma.09G075800 | N/A | uncharacterized protein |
| Glyma.13G194800 | leucine-rich repeat-containing protein | disease resistance |
| Glyma.13G194900 | leucine-rich repeat-containing protein | disease resistance |
| Glyma.16G028300 | N/A | uncharacterized protein |
| Glyma.18G166600 | phosphatidylinositol glycan, class s (PIGS) | protein glycosilation |
| Glyma.20G138000 | cysteine-rich repeat secretory protein | disease resistance |
| GeneID | SCN | Copy | %S | Ortholog | %S | Best-Hit-Arabi | S% | Proteic Domain |
|---|---|---|---|---|---|---|---|---|
| Glyma.11G228900 | 2 | Glyma.18G028300.1 | 92 | N/A | N/A | AT5G05690.1 | 80 | p450 |
| Glyma.13G217400 | 2 | Glyma.15G095000.1 | 90 | AT2G34500.1 | 70 | N/A | N/A | p450 |
| Glyma.08G204300 | 2 | Glyma.07G019100.1 | 94 | N/A | N/A | AT3G16170.1 | 61 | AMP-binding enzyme |
| Glyma.03G129700 | 2 | Glyma.19G132000.1 | 92 | N/A | N/A | AT3G57050.1 | 80 | Cys/Met metabolism PLP-dependent enzyme |
| Glyma.02G254600 | 2 | Glyma.14G061800.1 | 92 | AT2G39980.1 | 70 | N/A | N/A | Transferase family |
| Glyma.16G032200 | 2 | Glyma.07G065700 | 96 | N/A | N/A | AT3G61510.1 | 63 | Aminotransferase class I and II |
| Glyma.08G108800 | 3 | Glyma.05G152000.1/Glyma.11G254700.1 | 99/93 | N/A | N/A | AT4G13940.1 | 93 | S-adenosyl-L-homocysteine hydrolase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kafer, J.M.; Molinari, M.D.C.; Henning, F.A.; Koltun, A.; Marques, V.V.; Marin, S.R.R.; Nepomuceno, A.L.; Mertz-Henning, L.M. Transcriptional Profile of Soybean Seeds with Contrasting Seed Coat Color. Plants 2023, 12, 1555. https://doi.org/10.3390/plants12071555
Kafer JM, Molinari MDC, Henning FA, Koltun A, Marques VV, Marin SRR, Nepomuceno AL, Mertz-Henning LM. Transcriptional Profile of Soybean Seeds with Contrasting Seed Coat Color. Plants. 2023; 12(7):1555. https://doi.org/10.3390/plants12071555
Chicago/Turabian StyleKafer, João M., Mayla D. C. Molinari, Fernando A. Henning, Alessandra Koltun, Viviani V. Marques, Silvana R. R. Marin, Alexandre L. Nepomuceno, and Liliane M. Mertz-Henning. 2023. "Transcriptional Profile of Soybean Seeds with Contrasting Seed Coat Color" Plants 12, no. 7: 1555. https://doi.org/10.3390/plants12071555