
Toxic Effects of Perilla frutescens (L.) Britt. Essential Oil and Its Main Component on Culex pipiens pallens (Diptera: Culicidae)
Abstract
:1. Introduction
2. Results
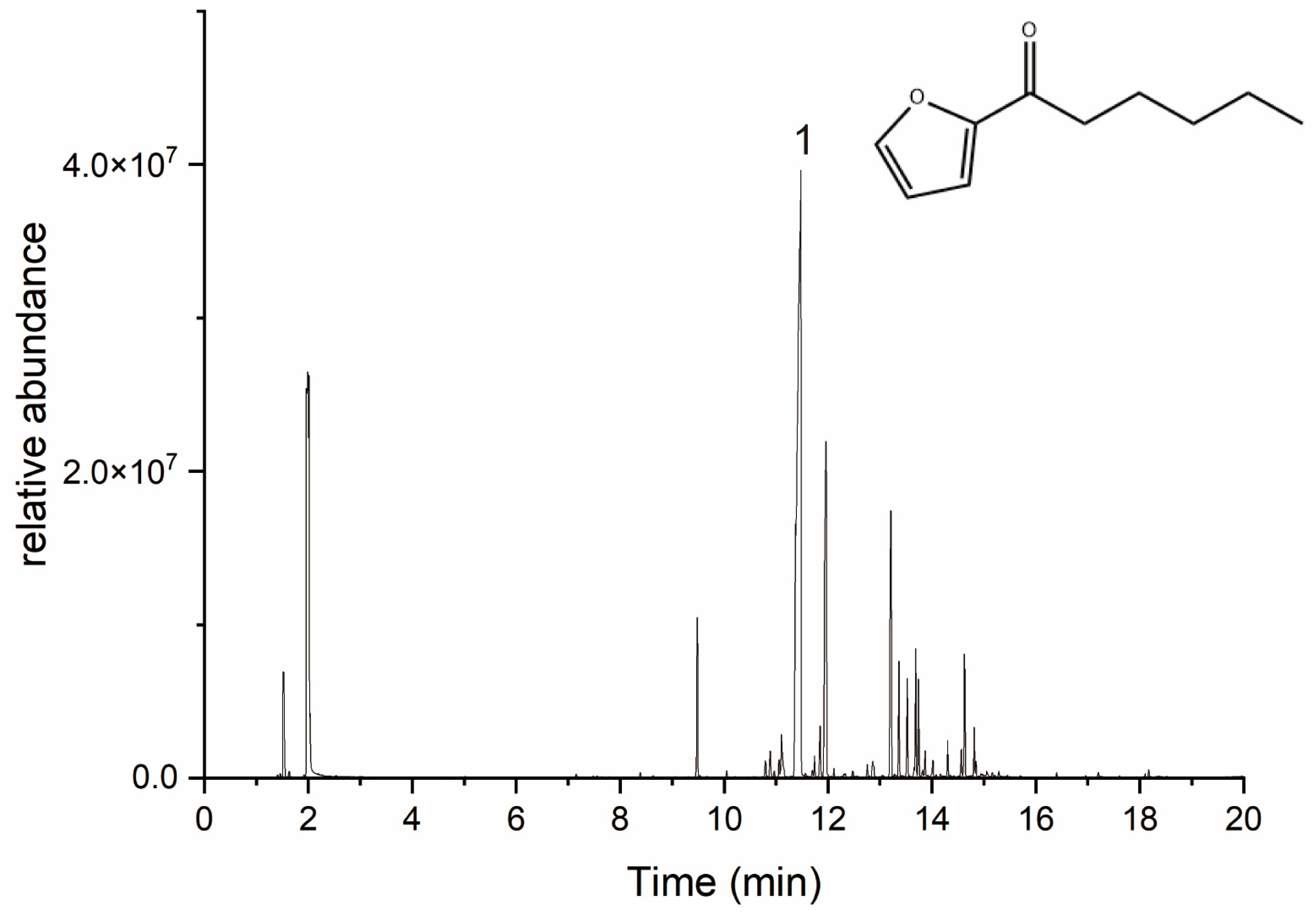
2.1. Gas Chromatography–Mass Spectrometry Analysis of Perilla Essential Oil Composition
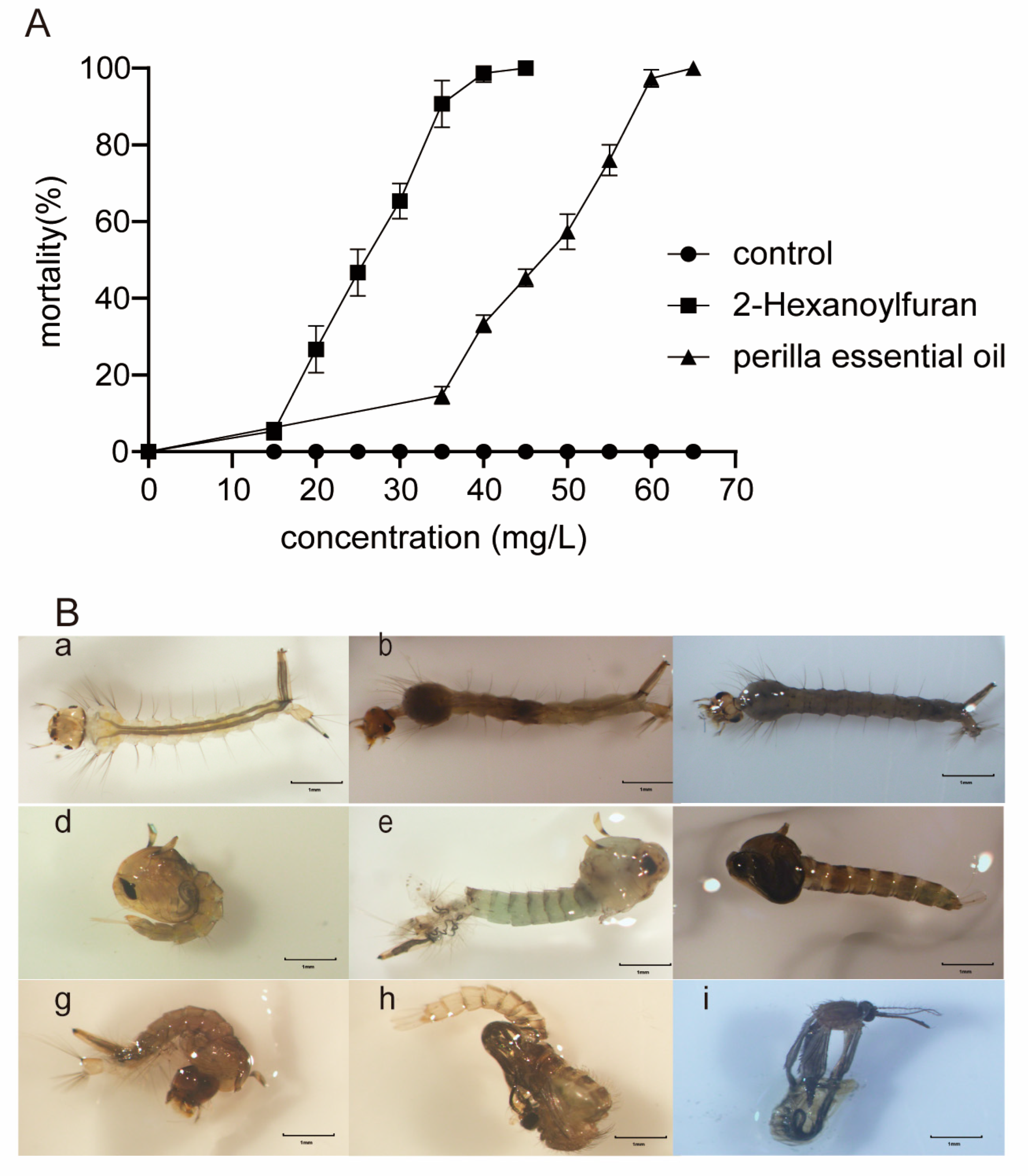
2.2. Larvicidal Activity
2.3. Ovicidal Activity
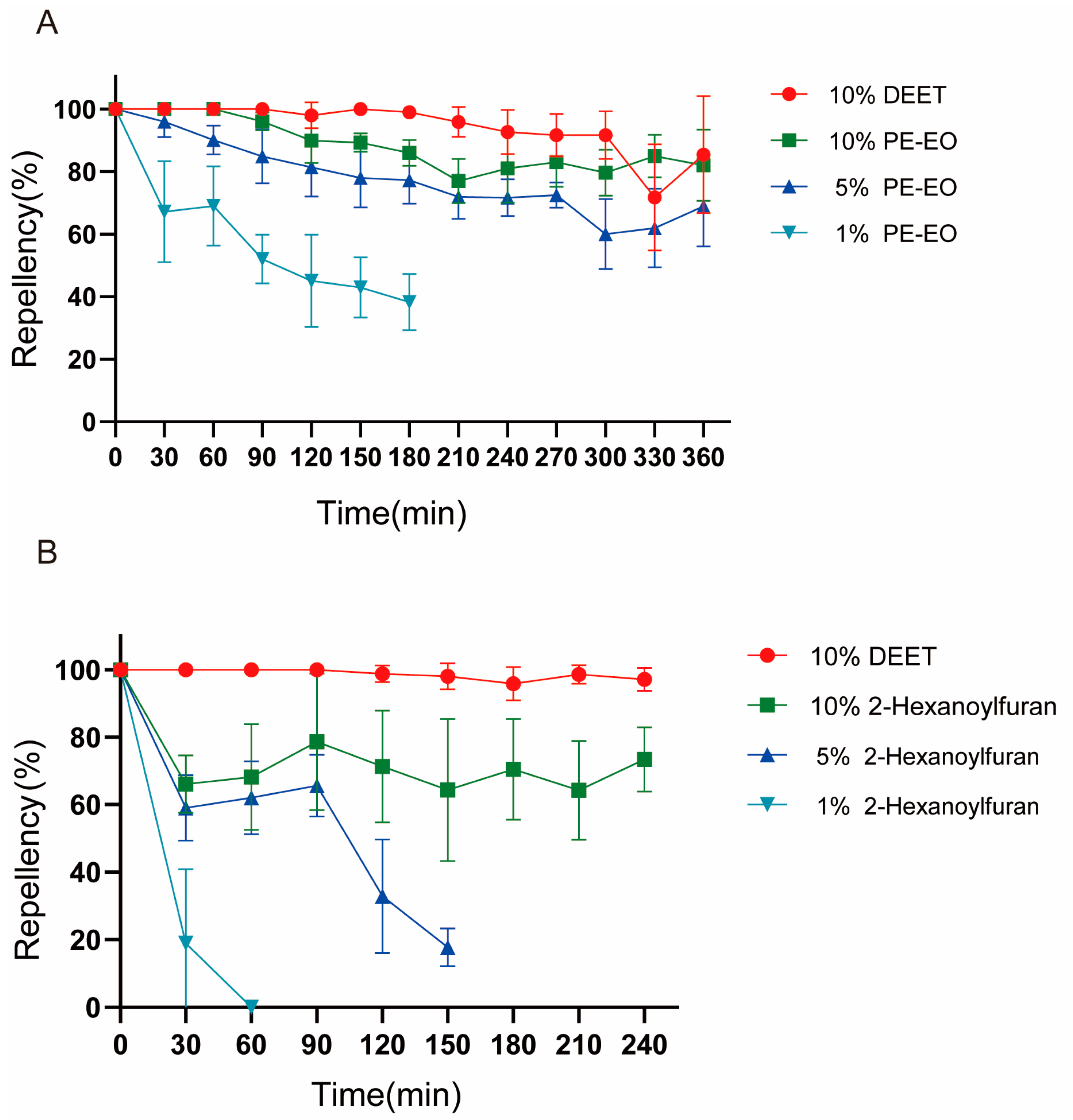
2.4. Repellency
2.5. Oviposition Assay
2.6. Gut Histopathology
2.7. Effects on Nontarget Species
3. Discussion
4. Materials and Methods
4.1. Mosquitoes
4.2. Plant Material and Extraction
4.3. Gas Chromatography–Mass Spectrometry Analysis of Essential Oil
4.4. Larvicidal Activity Bioassay
4.5. Ovicidal Activity Bioassay
4.6. Repellency Bioassay
4.7. Oviposition Assay
4.8. Gut Histopathology
4.9. Effects on Nontarget Organisms
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PE-EO | Perilla frutescens essential oil |
| WHO | World Health Organization |
| DEET | N, N-diethyl-m-toluamide |
| LC50 | median lethal concentration |
| LC90 | 90% lethal concentration |
| GC-MS | gas chromatography–mass spectrometry |
| ER | effective repellency |
| OAI | oviposition activity index |
| CCK8 | Cell Counting Kit-8 |
References
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achee, N.L.; Grieco, J.P.; Vatandoost, H.; Seixas, G.; Pinto, J.; Ching-Ng, L.; Martins, A.J.; Juntarajumnong, W.; Corbel, V.; Gouagna, C.; et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl. Trop. Dis. 2019, 13, e0006822. [Google Scholar] [CrossRef] [Green Version]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.-P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef] [PubMed]
- Namias, A.; Jobe, N.B.; Paaijmans, K.P.; Huijben, S. The need for practical insecticide-resistance guidelines to effectively inform mosquito-borne disease control programs. ELife 2021, 10, e65655. [Google Scholar] [CrossRef]
- Corbel, V.; Achee, N.L.; Chandre, F.; Coulibaly, M.B.; Dusfour, I.; Fonseca, D.M.; Grieco, J.; Juntarajumnong, W.; Lenhart, A.; Martins, A.J.; et al. Tracking Insecticide Resistance in Mosquito Vectors of Arboviruses: The Worldwide Insecticide resistance Network (WIN). PLoS Negl. Trop. Dis. 2016, 10, e0005054. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, J.; Khatri, M.; Arya, S.K. A treatise on Organophosphate pesticide pollution: Current strategies and advancements in their environmental degradation and elimination. Ecotoxicol. Environ. Saf. 2021, 207, 111483. [Google Scholar] [CrossRef]
- Silvério, M.R.S.; Espindola, L.S.; Lopes, N.P.; Vieira, P.C. Plant Natural Products for the Control of: The Main Vector of Important Arboviruses. Molecules 2020, 25, 3484. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A Review of Resistance Mechanisms of Synthetic Insecticides and Botanicals, Phytochemicals, and Essential Oils as Alternative Larvicidal Agents Against Mosquitoes. Front. Physiol. 2019, 10, 1591. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Qiu, J.-F.; Ma, L.-J.; Hu, Y.-J.; Li, P.; Wan, J.-B. Phytochemical and phytopharmacological review of Perilla frutescens L. (Labiatae), a traditional edible-medicinal herb in China. Food Chem. Toxicol. 2017, 108, 375–391. [Google Scholar] [CrossRef]
- Ahmed, H.M. Ethnomedicinal, Phytochemical and Pharmacological Investigations of (L.) Britt. Molecules 2018, 24, 102. [Google Scholar] [CrossRef] [Green Version]
- Tabanca, N.; Demirci, B.; Ali, A.; Ali, Z.; Blythe, E.K.; Khan, I.A. Essential oils of green and red Perilla frutescens as potential sources of compounds for mosquito management. Ind. Crops Prod. 2015, 65, 36–44. [Google Scholar] [CrossRef]
- Willis, M.D.; Saunders, M.E.; Ramoutar, D.; Tyszko, J.M.; Krenick, F.N.; Rohrbacher, A. Insecticidal compositions and methods of using the. same. Patent No. AU2017245410(A1), 26 October 2017. [Google Scholar]
- Zhong, Y.; Zheng, Q.; Hu, P.; Huang, X.; Yang, M.; Ren, G.; Li, J.; Du, Q.; Liu, S.; Zhang, K.; et al. Sedative and hypnotic effects of Perilla frutescens essential oil through GABAergic system pathway. J. Ethnopharmacol. 2021, 279, 113627. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Zeng, X.; Zhang, S.; Wang, Y.; Zhang, P.; Lü, A.; Peng, X. Regional variation in components and antioxidant and antifungal activities of Perilla frutescens essential oils in China. Ind. Crops Prod. 2014, 59, 69–79. [Google Scholar] [CrossRef]
- Karpiński, T.M. Essential Oils of Lamiaceae Family Plants as Antifungals. Biomolecules 2020, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranasinghe, H.A.K.; Amarasinghe, L.D. Naturally Occurring Microbiota Associated with Mosquito Breeding Habitats and Their Effects on Mosquito Larvae. Biomed. Res. Int. 2020, 2020, 4065315. [Google Scholar] [CrossRef]
- Kelly, P.H.; Yingling, A.V.; Ahmed, A.; Hurwitz, I.; Ramalho-Ortigao, M. Defining the mechanisms of action and mosquito larva midgut response to a yeast-encapsulated orange oil larvicide. Parasites Vectors 2022, 15, 183. [Google Scholar] [CrossRef]
- Luz, T.R.S.A.; de Mesquita, L.S.S.; Amaral, F.M.M.d.; Coutinho, D.F. Essential oils and their chemical constituents against Aedes aegypti L. (Diptera: Culicidae) larvae. Acta Trop. 2020, 212, 105705. [Google Scholar] [CrossRef]
- Rants'o, T.A.; Koekemoer, L.L.; Panayides, J.-L.; van Zyl, R.L. Potential of Essential Oil-Based Anticholinesterase Insecticides against Vectors: A Review. Molecules 2022, 27, 7026. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Kim, J.-R.; Oh, S.M.; Jung, J.W.; Ahn, Y.-J.; Kwon, H.W. Novel histopathological and molecular effects of natural compound pellitorine on larval midgut epithelium and anal gills of Aedes aegypti. PLoS ONE 2013, 8, e80226. [Google Scholar] [CrossRef] [Green Version]
- Fiaz, M.; Martínez, L.C.; Costa, M.d.S.; Cossolin, J.F.S.; Plata-Rueda, A.; Gonçalves, W.G.; Sant'Ana, A.E.G.; Zanuncio, J.C.; Serrão, J.E. Squamocin induce histological and ultrastructural changes in the midgut cells of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Ecotoxicol. Environ. Saf. 2018, 156, 1–8. [Google Scholar] [CrossRef]
- Bibi, R.; Tariq, R.M.; Rasheed, M. Toxic assessment, growth disrupting and neurotoxic effects of red seaweeds' botanicals against the dengue vector mosquito Aedes aegypti L. Ecotoxicol. Environ. Saf. 2020, 195, 110451. [Google Scholar] [CrossRef] [PubMed]
- Kabir, K.E.; Choudhary, M.I.; Ahmed, S.; Tariq, R.M. Growth-disrupting, larvicidal and neurobehavioral toxicity effects of seed extract of Seseli diffusum against Aedes aegypti (L.) (Diptera: Culicidae). Ecotoxicol. Environ. Saf. 2013, 90, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Plant-borne ovicides in the fight against mosquito vectors of medical and veterinary importance: A systematic review. Parasitol. Res. 2015, 114, 3201–3212. [Google Scholar] [CrossRef] [PubMed]
- Faustino, C.G.; de Medeiros, F.A.; Galardo, A.K.R.; Rodrigues, A.B.L.; da Costa, A.L.P.; Martins, R.L.; Brandão, L.B.; Santos, L.L.; de Medeiros, M.A.A.; de Castro Cantuária, P.; et al. Biocidal Activity of a Nanoemulsion Containing Essential Oil from Resin against (Diptera: Culicidae). Molecules 2021, 26, 6439. [Google Scholar] [CrossRef]
- Suman, D.S.; Wang, Y.; Bilgrami, A.L.; Gaugler, R. Ovicidal activity of three insect growth regulators against Aedes and Culex mosquitoes. Acta Trop. 2013, 128, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.D.d.L.; de Moura, K.S.; Napoleão, T.H.; Santos, G.K.N.; Coelho, L.C.B.B.; Navarro, D.M.d.A.F.; Paiva, P.M.G. Oviposition-stimulant and ovicidal activities of Moringa oleifera lectin on Aedes aegypti. PLoS ONE 2012, 7, e44840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muturi, E.J.; Selling, G.W.; Doll, K.M.; Hay, W.T.; Ramirez, J.L. Leptospermum scoparium essential oil is a promising source of mosquito larvicide and its toxicity is enhanced by a biobased emulsifier. PLoS ONE 2020, 15, e0229076. [Google Scholar] [CrossRef] [Green Version]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Rakotosaona, R.; Nzekoue, F.K.; Canale, A.; Nicoletti, M.; Maggi, F. Insecticidal and mosquito repellent efficacy of the essential oils from stem bark and wood of Hazomalania voyronii. J. Ethnopharmacol. 2020, 248, 112333. [Google Scholar] [CrossRef]
- Legeay, S.; Clere, N.; Apaire-Marchais, V.; Faure, S.; Lapied, B. Unusual modes of action of the repellent DEET in insects highlight some human side effects. Eur. J. Pharmacol. 2018, 825, 92–98. [Google Scholar] [CrossRef]
- Norris, E.J.; Coats, J.R. Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control. Int. J. Environ. Res. Public Health 2017, 14, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-X.; Ma, Y.-P.; Zhang, H.-X.; Sun, H.-Z.; Su, H.-H.; Pei, S.-J.; Du, Z.-Z. Repellent, larvicidal and adulticidal activities of essential oil from Dai medicinal plant against. Plant Divers. 2021, 43, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition Deterrent and Larvicidal and Pupaecidal Activity of Seven Essential Oils and their Major Components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism-antagonism Effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaalan, E.A.-S.; Canyon, D.V. Mosquito oviposition deterrents. Environ. Sci. Pollut. Res. Int. 2018, 25, 10207–10217. [Google Scholar] [CrossRef] [PubMed]
- Canyon, D.V.; Muller, R. Oviposition and olfaction responses of Aedes aegypti mosquitoes to insecticides. Trop. Biomed. 2013, 30, 691–698. [Google Scholar] [PubMed]
- Rajeswary, M.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Benelli, G. Zingiber cernuum (Zingiberaceae) essential oil as effective larvicide and oviposition deterrent on six mosquito vectors, with little non-target toxicity on four aquatic mosquito predators. Environ. Sci. Pollut. Res. Int. 2018, 25, 10307–10316. [Google Scholar] [CrossRef]
- Galvão, J.G.; Cerpe, P.; Santos, D.A.; Gonsalves, J.K.; Santos, A.J.; Nunes, R.K.; Lira, A.A.; Alves, P.B.; La Corte, R.; Blank, A.F.; et al. Lippia gracilis essential oil in β-cyclodextrin inclusion complexes: An environmentally safe formulation to control Aedes aegypti larvae. Pest Manag. Sci. 2019, 75, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Lobato Rodrigues, A.B.; Martins, R.L.; Rabelo, É.d.M.; Tomazi, R.; Santos, L.L.; Brandão, L.B.; Faustino, C.G.; Ferreira Farias, A.L.; Dos Santos, C.B.R.; de Castro Cantuária, P.; et al. Development of nano-emulsions based on Ayapana triplinervis essential oil for the control of Aedes aegypti larvae. PLoS ONE 2021, 16, e0254225. [Google Scholar] [CrossRef]
- Yang, X.; Xu, Y.; Yin, Q.; Zhang, H.; Yin, H.; Sun, Y.; Ma, L.; Zhou, D.; Shen, B. Physiological characterization of chitin synthase A responsible for the biosynthesis of cuticle chitin in Culex pipiens pallens (Diptera: Culicidae). Parasites Vectors 2021, 14, 234. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, S.; Zhang, H.; Yin, H.; Wang, H.; Zhou, D.; Sun, Y.; Ma, L.; Shen, B.; Zhu, C. MiR-279-3p regulates deltamethrin resistance through CYP325BB1 in Culex pipiens pallens. Parasites Vectors 2021, 14, 528. [Google Scholar] [CrossRef]
- Li, N.; Zhang, Z.J.; Li, X.J.; Li, H.Z.; Cui, L.X.; He, D.L. Microcapsules biologically prepared using Perilla frutescens (L.) Britt. essential oil and their use for extension of fruit shelf life. J. Sci. Food Agric. 2017, 98, 1033. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Manh, H.D.; Tuyet, O.T. Larvicidal and Repellent Activity of L. Essential Oil against. Insects 2020, 11, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, P.; Stalin, A.; Gabriel Paulraj, M.; Balakrishna, K.; Ignacimuthu, S.; Abdullah Al-Dhabi, N. Biocontrol and non-target effect of fractions and compound isolated from Streptomyces rimosus on the immature stages of filarial vector Culex quinquefasciatus Say (Diptera: Culicidae) and the compound interaction with Acetylcholinesterase (AChE1). Ecotoxicol. Environ. Saf. 2018, 161, 120–128. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines for Efficacy Testing of Mosquito Repellents for Human Skin; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Yu, K.-X.; Wong, C.-L.; Ahmad, R.; Jantan, I. Mosquitocidal and Oviposition Repellent Activities of the Extracts of Seaweed Bryopsis pennata on Aedes aegypti and Aedes albopictus. Molecules 2015, 20, 14082–14102. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | RT Time/Min | Peak Area (%) | Compound Name | CAS Number |
|---|---|---|---|---|
| 1 | 7.157 | 0.11% | Benzaldehyde | 100-52-7 |
| 2 | 8.391 | 0.12% | (+)-(R)-limonene | 5989-27-5 |
| 3 | 9.486 | 3.64% | Linalool | 78-70-6 |
| 4 | 10.802 | 0.42% | 3-Methyl-1-(3-methyl-2-furyl)-1-butanone | 488-05-1 |
| 5 | 10.892 | 0.63% | 1-Propanone, 1-(3-cyclohexen-1-yl)-2,2-dimethyl- | 16076-65-6 |
| 6 | 11.066 | 0.40% | 1H-Indene, 1-ethylideneoctahydro-7a-methyl-, (1E,3aR,7aS)-rel- | 56324-68-6 |
| 7 | 11.477 | 49.77% | 2-hexanoylfuran | 14360-50-0 |
| 8 | 11.706 | 0.20% | Cyclohexanemethanol, 4-(1-methylethenyl)-, trans- | 22451-48-5 |
| 9 | 11.747 | 0.44% | dl-Perillaldehyde | 2111-75-3 |
| 10 | 11.964 | 14.13% | Naphthalene, decahydro-1-methyl-2-methylene- | 90548-09-7 |
| 11 | 12.118 | 0.20% | Methylgeranate | 2349-14-6 |
| 12 | 12.481 | 0.20% | Eugenol | 97-53-0 |
| 13 | 12.865 | 0.65% | β-Elemene | 515-13-9 |
| 14 | 13.214 | 7.64% | β-Caryophyllene | 87-44-5 |
| 15 | 13.368 | 2.18% | (E)-β-Farnesene | 18794-84-8 |
| 16 | 13.53 | 1.95% | α-Caryophyllene | 6753-98-6 |
| 17 | 13.661 | 0.21% | β-Ionone | 14901-07-6 |
| 18 | 13.692 | 2.40% | (Z,E)- α-farnesene | 26560-14-5 |
| 19 | 13.746 | 2.10% | β-Cubebene | 13744-15-5 |
| 20 | 13.821 | 0.19% | (E,E)-α-Farnesene | 502-61-4 |
| 21 | 13.872 | 0.60% | 1-methyl-4-(1-methylethylidene)-2-(1-methylvinyl)-1-vinylcyclohexane | 3242-08-8 |
| 22 | 14.024 | 0.47% | beta-Cadinene | 523-47-7 |
| 23 | 14.306 | 0.65% | (±)-trans-Nerolidol | 40716-66-3 |
| 24 | 14.569 | 0.63% | 1H-Cycloprop[e]azulen-7-ol, decahydro-1,1,7-trimethyl-4-methylene-, (1aS,4aS,7R,7aS,7bS)- | 77171-55-2 |
| 25 | 14.632 | 2.68% | (−)-Caryophyllene oxide | 1139-30-6 |
| 26 | 14.816 | 0.99% | Apiole | 523-80-8 |
| 27 | 14.852 | 0.33% | humulene epoxide ii | 19888-34-7 |
| 28 | 15.168 | 0.19% | α-cadinol | 481-34-5 |
| 29 | 17.205 | 0.16% | Palmitic acid | 57-10-3 |
| 30 | 18.175 | 0.16% | Phytol | 150-86-7 |
| Total | 94.42% |
| Treatment | Concentration (mg/L) | Egg Hatchability (%) | F Value | p Value |
|---|---|---|---|---|
| control | - | 100 ± 0.0 | - | - |
| PE-EO | 70 | 90.2 ± 2.4 a | 383.9 | <0.001 |
| 80 | 65.2 ± 2.4 b | |||
| 90 | 41.7 ± 4.2 c | |||
| 100 | 19.4 ± 6.5 d | |||
| 110 | 15.3 ± 2.4 d | |||
| 120 | 1.30 ± 2.4 e | |||
| 2-Hexanoylfuran | 40 | 86.1 ± 2.4 a | 153.21 | <0.001 |
| 50 | 62.5 ± 4.2 b | |||
| 60 | 56.9 ± 2.4 b | |||
| 70 | 23.6 ± 8.7 c | |||
| 80 | 1.38 ± 2.4 d |
| Treatment | Concentration (mg/L) | ER% | OAI | Effect (A/N/R) |
|---|---|---|---|---|
| PE-EO | 20 | 49.4 ± 12 c | −0.33 | R |
| 40 | 81.3 ± 1.8 b | −0.69 | R | |
| 60 | 87.7 ± 1.5 ab | −0.78 | R | |
| 80 | 90.0 ± 1.0 ab | −0.82 | R | |
| 100 | 94.7 ± 2.7 a | −0.89 | R | |
| 2-Hexanoylfuran | 10 | 62.6 ± 2.1 d | −0.46 | R |
| 15 | 72.1 ± 5.6 c | −0.57 | R | |
| 20 | 85.1 ± 0.7 b | −0.74 | R | |
| 25 | 88.1 ± 1.5 ab | −0.79 | R | |
| 30 | 93.1 ± 2.3 a | −0.87 | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Zhang, W.; Zheng, J.; Xu, J.; Wang, H.; Du, J.; Zhou, D.; Sun, Y.; Shen, B. Toxic Effects of Perilla frutescens (L.) Britt. Essential Oil and Its Main Component on Culex pipiens pallens (Diptera: Culicidae). Plants 2023, 12, 1516. https://doi.org/10.3390/plants12071516
Zhang R, Zhang W, Zheng J, Xu J, Wang H, Du J, Zhou D, Sun Y, Shen B. Toxic Effects of Perilla frutescens (L.) Britt. Essential Oil and Its Main Component on Culex pipiens pallens (Diptera: Culicidae). Plants. 2023; 12(7):1516. https://doi.org/10.3390/plants12071516
Chicago/Turabian StyleZhang, Ruimin, Wenxing Zhang, Junnan Zheng, Jingwei Xu, Huan Wang, Jiajia Du, Dan Zhou, Yan Sun, and Bo Shen. 2023. "Toxic Effects of Perilla frutescens (L.) Britt. Essential Oil and Its Main Component on Culex pipiens pallens (Diptera: Culicidae)" Plants 12, no. 7: 1516. https://doi.org/10.3390/plants12071516