
Isolation and Characterization of the Genes Involved in the Berberine Synthesis Pathway in Asian Blue Cohosh, Caulophyllum robustum
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Berberine Extraction and Quantification
2.3. RNA Extraction and Transcriptome Sequencing
2.4. De Novo Assembly
2.5. Functional Annotations
2.6. Isolation of Berberine Synthesis Genes
2.7. Phylogenetic Analysis
3. Results
3.1. Berberine Contents
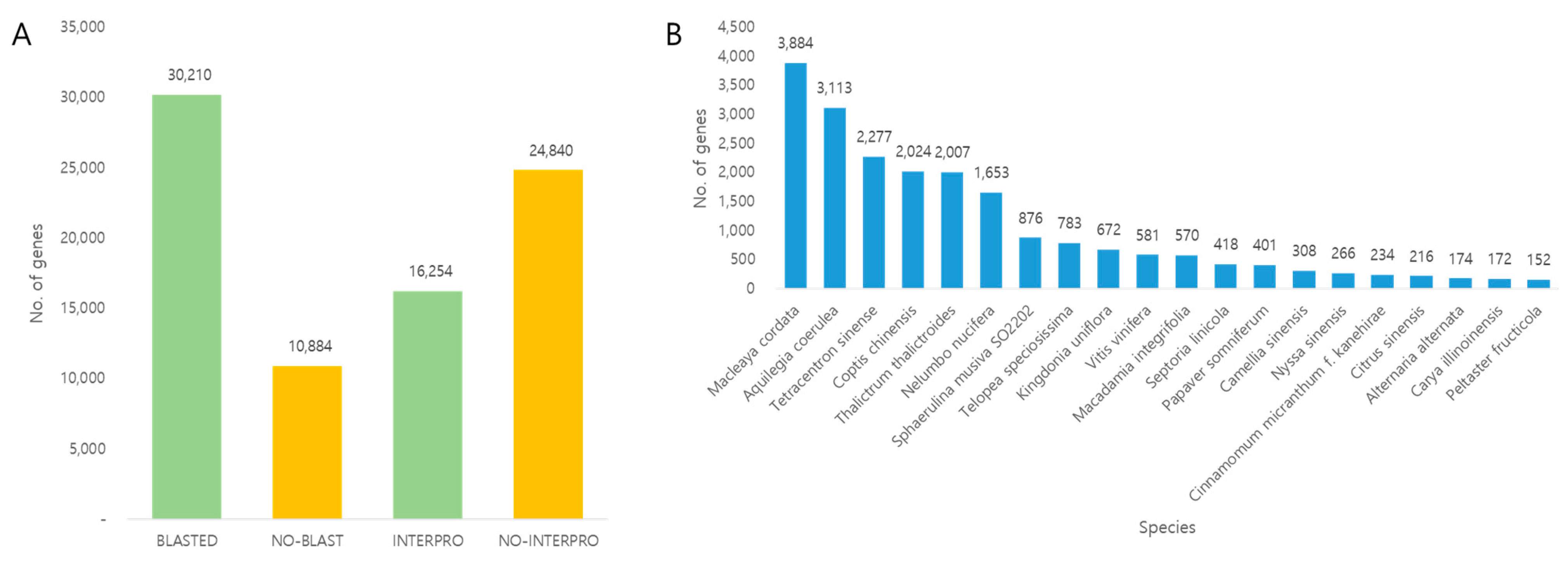
3.2. Transcriptome Sequencing
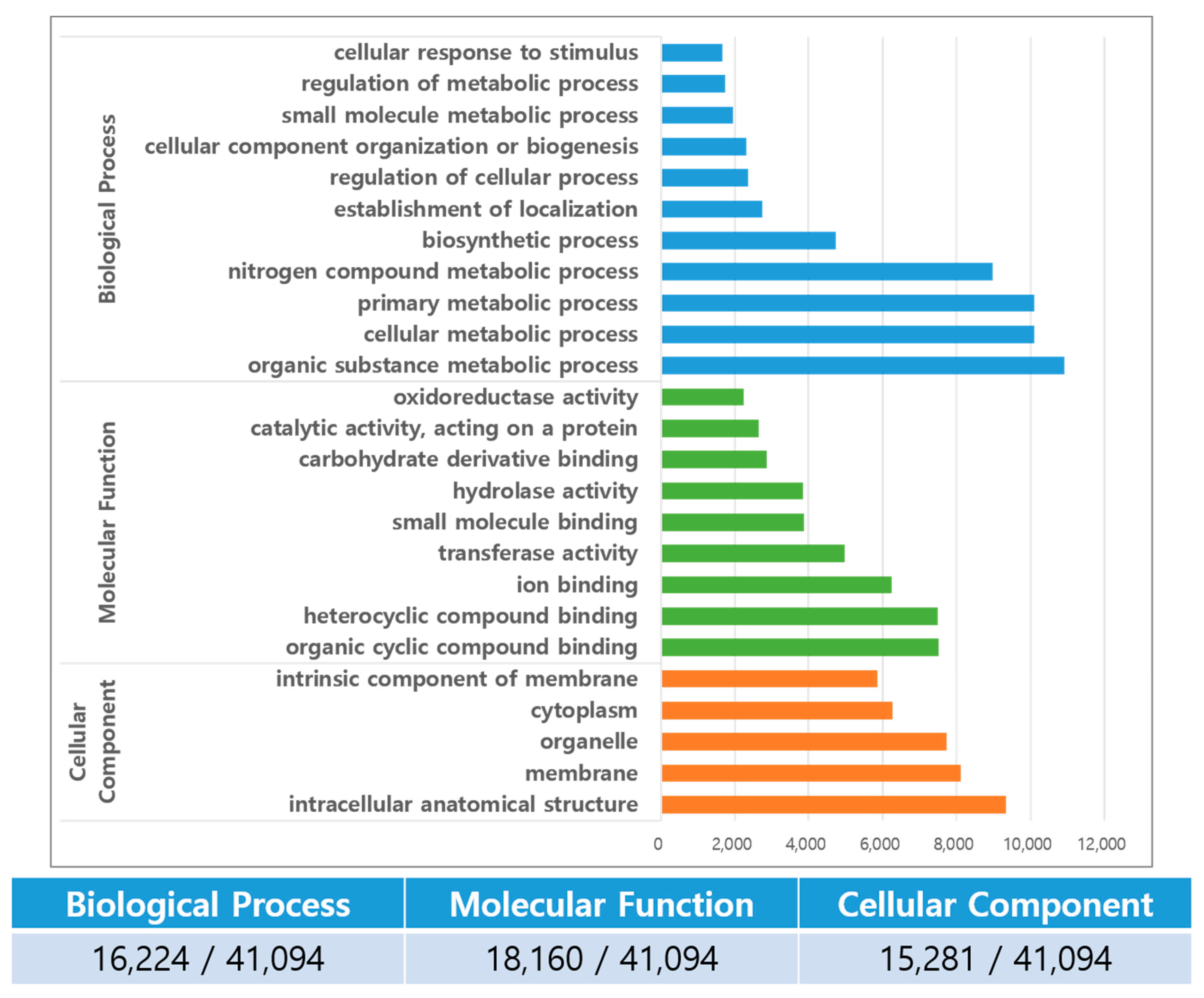
3.3. Functional Annotation and Gene Ontology (GO)
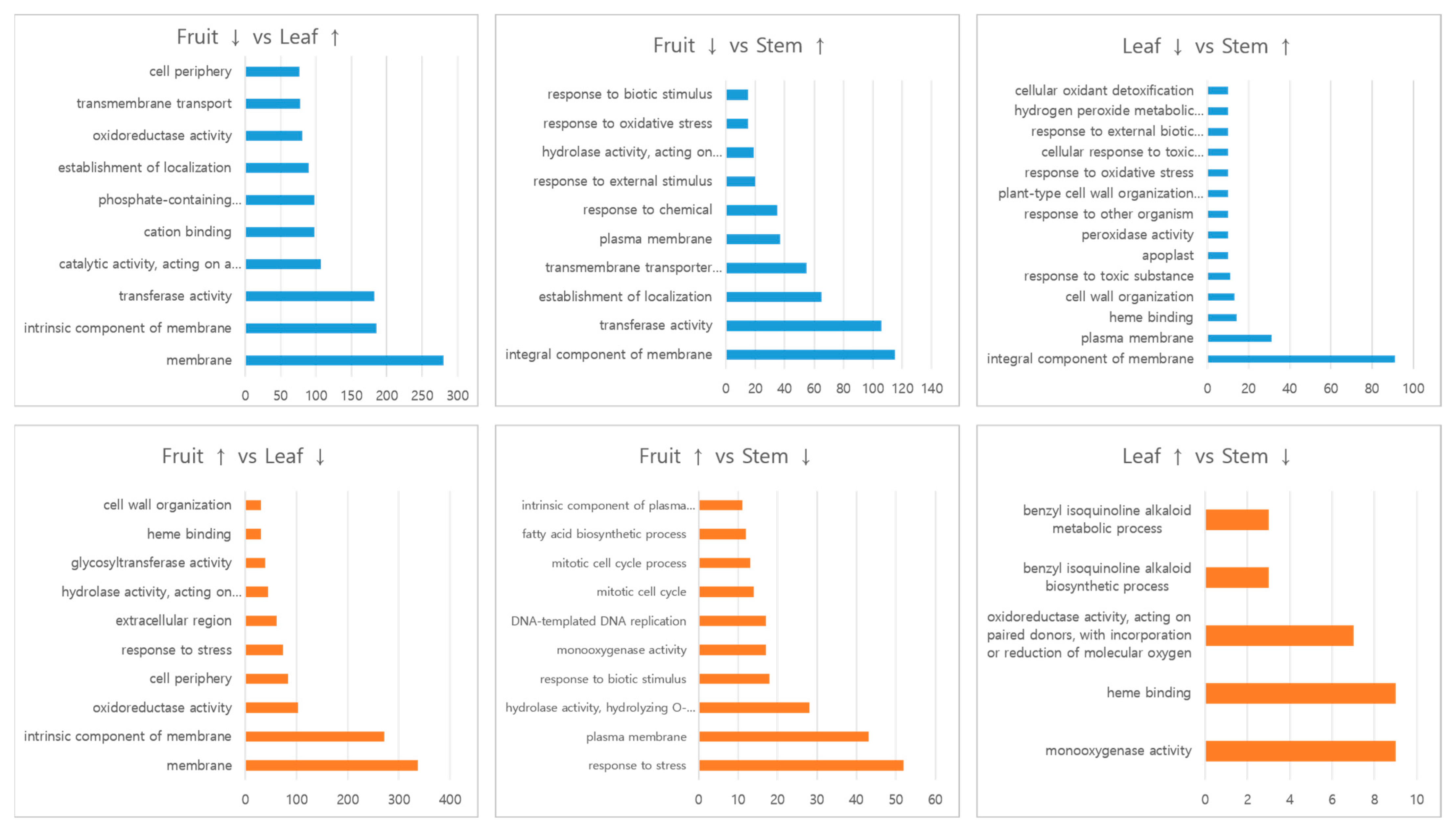
3.4. Differences of the Genes Expressed among Leaves, Stem, and Fruit in C. robustum
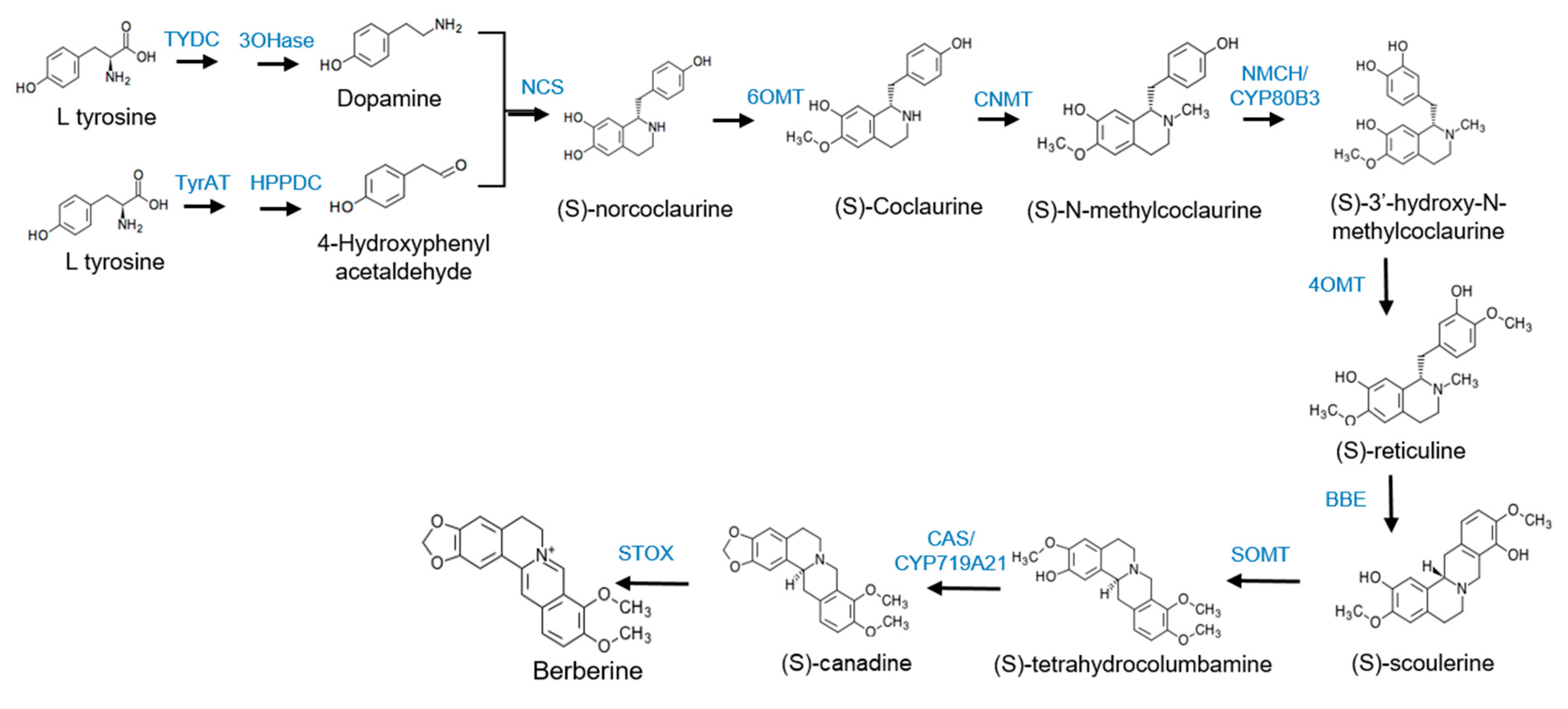
3.5. Isolation of Berberine Synthesis Genes
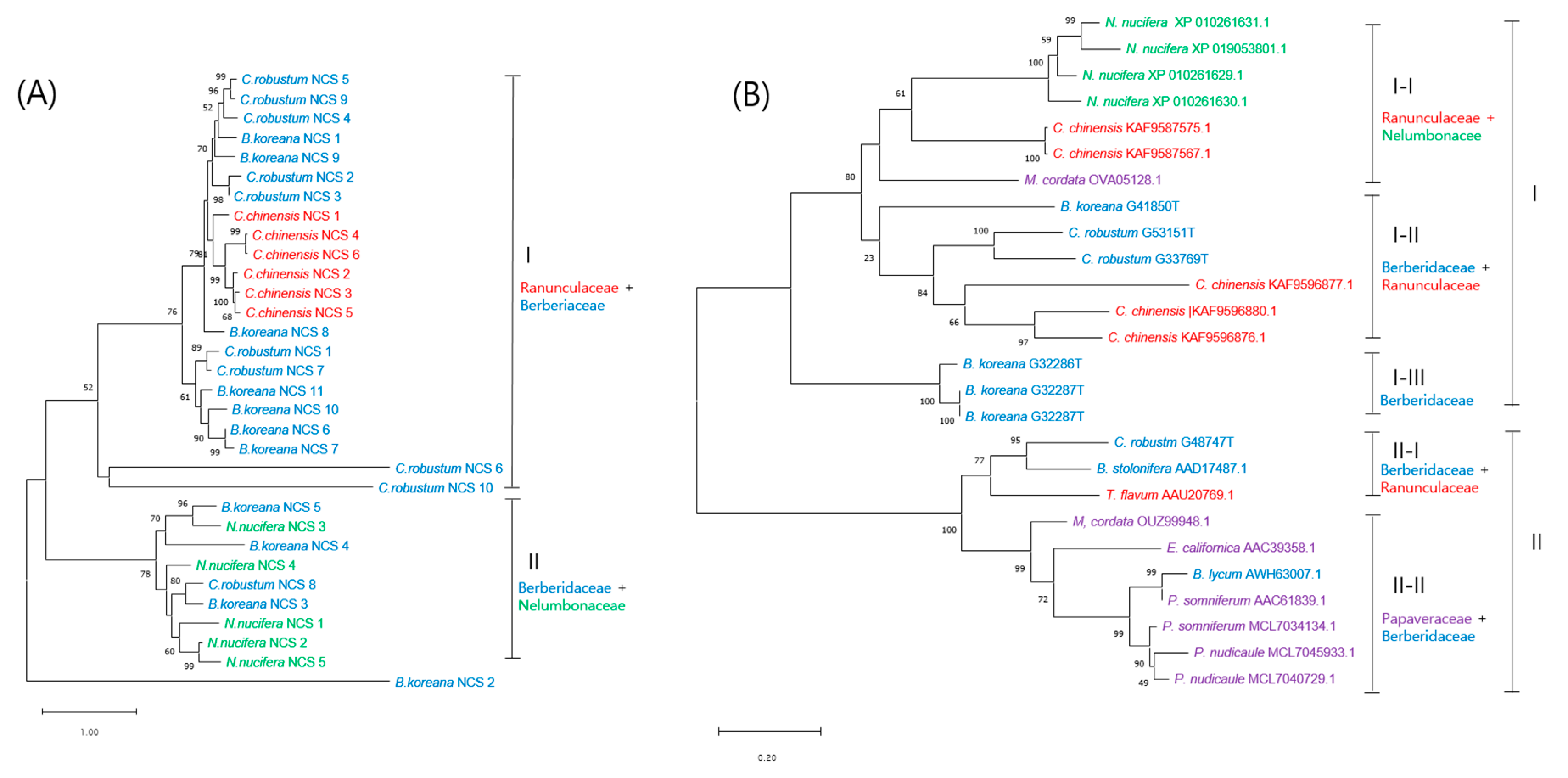
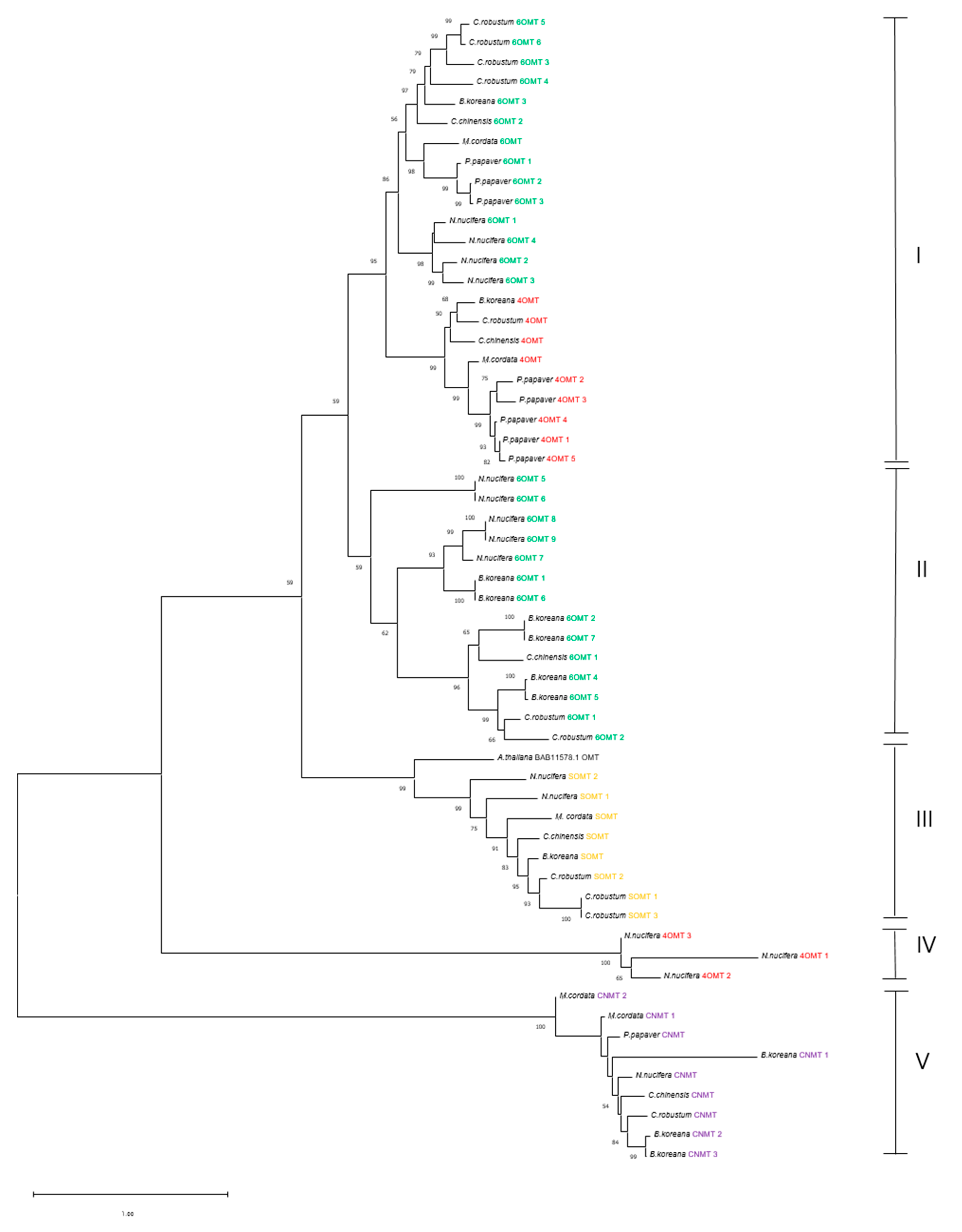
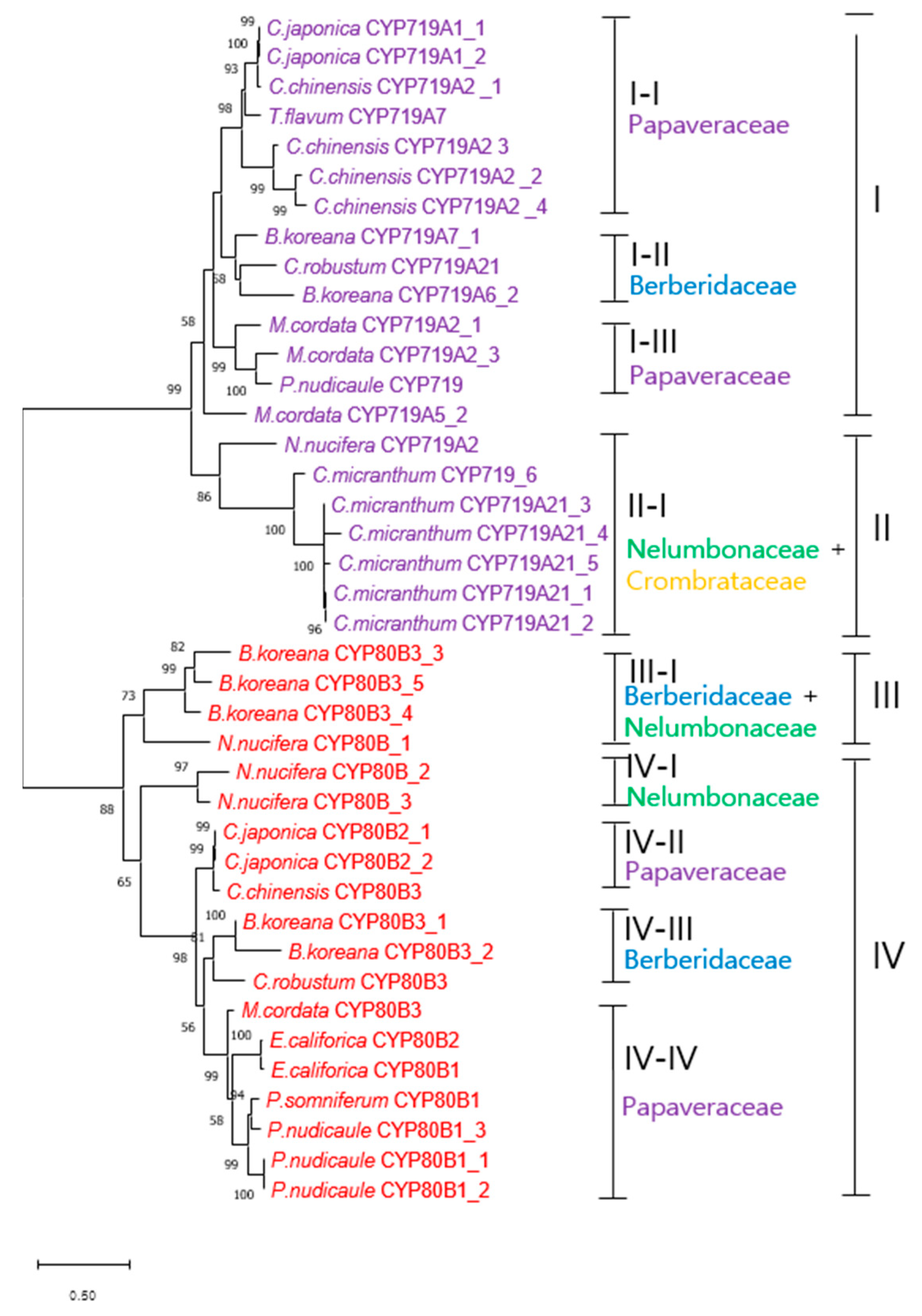
3.6. Phylogenetic Analysis of Methyltransferases and CYP450 Monooxygenases
4. Discussion
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loconte, H.; Estes, J.R. Generic relationships within Leonticeae (Berberidaceae). Can. J. Bot. 1989, 67, 2310–2316. [Google Scholar] [CrossRef]
- Hutchens, A.R. A Handbook of Native American Herbs: The Pocket Guide to 125 Medicinal Plants and Their Uses; Shambhala Publications: Boston, MA, USA, 1992. [Google Scholar]
- Satchithanandam, S.; Grundel, E.; Roach, J.; White, K.D.; Mazzola, E.; Ganzera, M.; Rader, J.I. Alkaloids and saponins in dietary supplements of blue cohosh (Caulophyllum thalictroides). J. AOAC Int. 2008, 91, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rader, J.I.; Pawar, R.S. Primary constituents of blue cohosh: Quantification in dietary supplements and potential for toxicity. Anal. Bioanal. Chem. 2013, 405, 4409–4417. [Google Scholar] [CrossRef]
- Kim, Y.-D. Berberidaceae. In The Genera of Vascular Plants of Korea; Flora of Korea Editorial Committee, Ed.; Academy Publishing Company: Seoul, Republic of Korea, 2007; pp. 206–209. [Google Scholar]
- Xia, Y.G.; Li, G.Y.; Liang, J.; Yang, B.Y.; Lu, S.W.; Kuang, H.X. Genus caulophyllum: An overview of chemistry and bioactivity. Evid. Based Complement. Alternat. Med. 2014, 2014, 684508. [Google Scholar] [CrossRef] [Green Version]
- Qin, B.H.; Liu, X.Q.; Yuan, Q.Y.; Wang, J.; Han, H.Y. Anti-Inflammatory Triterpenoids from the Caulophyllum robustum Maximin LPS-Stimulated RAW264.7 Cells. Molecules 2018, 23, 1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Jung, J.C.; Ali, Z.; Khan, I.A.; Oh, S. Anti-Inflammatory Effect of Triterpene Saponins Isolated from Blue Cohosh (Caulophyllum thalictroides). Evid. Based Complement. Altern. Med. 2012, 2012, 798192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.; Yu, S.-S.; Du, D.; Wang, Y.-D. Bioactive constituents from toxic seed plants in China. RSC Adv. 2013, 3, 10078–10102. [Google Scholar] [CrossRef]
- Lu, S.; Liu, Y.; Cui, J.; Yang, B.; Li, G.; Guo, Y.; Kuang, H.; Wang, Q. Mechanism of Caulophyllum robustum Maxim against rheumatoid arthritis using LncRNA-mRNA chip analysis. Gene 2020, 722, 144105. [Google Scholar] [CrossRef]
- Ali, Z.; Khan, I.A. Alkaloids and saponins from blue cohosh. Phytochemistry 2008, 69, 1037–1042. [Google Scholar] [CrossRef]
- Singla, D.; Sharma, A.; Kaur, J.; Panwar, B.; Raghava, G.P. BIAdb: A curated database of benzylisoquinoline alkaloids. BMC Pharmacol. 2010, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Farrow, S.C.; Hagel, J.M.; Facchini, P.J. Transcript and metabolite profiling in cell cultures of 18 plant species that produce benzylisoquinoline alkaloids. Phytochemistry 2012, 77, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Liscombe, D.K.; Macleod, B.P.; Loukanina, N.; Nandi, O.I.; Facchini, P.J. Evidence for the monophyletic evolution of benzylisoquinoline alkaloid biosynthesis in angiosperms. Phytochemistry 2005, 66, 1374–1393. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Perdomo, I.M.; Facchini, P.J. Benzylisoquinoline Alkaloids Biosynthesis in Sacred Lotus. Molecules 2018, 23, 2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imenshahidi, M.; Hosseinzadeh, H. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res. 2019, 33, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, Z.; He, L. An approach to develop binary chromatographic fingerprints of the total alkaloids from Caulophyllum robustum by high performance liquid chromatography/diode array detector and gas chromatography/mass spectrometry. J. Pharm. Biomed. Anal. 2007, 43, 1667–1672. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Jeong, S.W.; Lee, W.S.; Park, S.; Kim, Y.H.; Kim, G.S.; Lee, S.J.; Jin, J.S.; Kim, C.Y.; Lee, J.E.; et al. Determination of Polyphenol Components of Korean Prostrate Spurge (Euphorbia supina) by Using Liquid Chromatography-Tandem Mass Spectrometry: Overall Contribution to Antioxidant Activity. J. Anal. Methods Chem. 2014, 2014, 418690. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Roy, N.S.; Choi, I.Y.; Um, T.; Jeon, M.J.; Kim, B.Y.; Kim, Y.D.; Yu, J.K.; Kim, S.; Kim, N.S. Gene Expression and Isoform Identification of PacBio Full-Length cDNA Sequences for Berberine Biosynthesis in Berberis koreana. Plants 2021, 10, 1314. [Google Scholar] [CrossRef]
- Choi, B.S.; Choi, S.K.; Kim, N.S.; Choi, I.Y. NBLAST: A graphical user interface-based two-way BLAST software with a dot plot viewer. Genomics Inform. 2022, 20, e36. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Battistuzzi, F.U. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bird, D.A.; Facchini, P.J. Berberine bridge enzyme, a key branch-point enzyme in benzylisoquinoline alkaloid biosynthesis, contains a vacuolar sorting determinant. Planta 2001, 213, 888–897. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Kaushik, N. Phytochemical and pharmacological studies in genus Berberis. Phytochem. Rev. 2013, 11, 523–542. [Google Scholar] [CrossRef]
- He, S.M.; Liang, Y.L.; Cong, K.; Chen, G.; Zhao, X.; Zhao, Q.M.; Zhang, J.J.; Wang, X.; Dong, Y.; Yang, J.L.; et al. Identification and Characterization of Genes Involved in Benzylisoquinoline Alkaloid Biosynthesis in Coptis Species. Front. Plant. Sci. 2018, 9, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Winzer, T.; He, Z.; Graham, I.A. Over 100 Million Years of Enzyme Evolution Underpinning the Production of Morphine in the Papaveraceae Family of Flowering Plants. Plant. Commun. 2020, 1, 100029. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Liu, W.; Xu, M.; Jiang, R.; Xia, L.; Wang, P.; Li, H.; Tang, Z.; Zheng, Q.; Zeng, J. Modulation of benzylisoquinoline alkaloid biosynthesis by overexpression berberine bridge enzyme in Macleaya cordata. Sci. Rep. 2018, 8, 17988. [Google Scholar] [CrossRef] [Green Version]
- Worberg, A.; Quandt, D.; Barniske, A.-M.; Löhne, C.; Hilu, K.W.; Borsch, T. Phylogeny of basal eudicots: Insights from non-coding and rapidly evolving DNA. Org. Divers. Evol. 2007, 7, 55–77. [Google Scholar] [CrossRef] [Green Version]
- Zhong, F.; Huang, L.; Qi, L.; Ma, Y.; Yan, Z. Full-length transcriptome analysis of Coptis deltoidea and identification of putative genes involved in benzylisoquinoline alkaloids biosynthesis based on combined sequencing platforms. Plant. Mol. Biol. 2020, 102, 477–499. [Google Scholar] [CrossRef]
- Itoh, A.; Saitoh, T.; Tani, K.; Uchigaki, M.; Sugimoto, Y.; Yamada, J.; Nakajima, H.; Ohshiro, H.; Sun, S.; Tanahashi, T. Bisbenzylisoquinoline Alkaloids from Nelumbo nucifera. Chem. Pharm. Bull. 2011, 59, 947–951. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Zhao, L.; Fang, T.; Xiong, Y.; Ogutu, C.; Yang, D.; Vimolmangkang, S.; Liu, Y.; Han, Y. Investigation of benzylisoquinoline alkaloid biosynthetic pathway and its transcriptional regulation in lotus. Hortic. Res. 2018, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Wenkert, E. The Alkaloids. Chemistry and Physiology. Volume VIII. The Indole Alkaloids. J. Am. Chem. Soc. 1966, 88, 3681. [Google Scholar] [CrossRef]
- Hagel, J.M.; Morris, J.S.; Lee, E.J.; Desgagne-Penix, I.; Bross, C.D.; Chang, L.; Chen, X.; Farrow, S.C.; Zhang, Y.; Soh, J.; et al. Transcriptome analysis of 20 taxonomically related benzylisoquinoline alkaloid-producing plants. BMC Plant. Biol. 2015, 15, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.D.; Dang, T.T. Cytochrome P450 Enzymes as Key Drivers of Alkaloid Chemical Diversification in Plants. Front. Plant. Sci. 2021, 12, 682181. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.S.; Park, N.I.; Kim, N.S.; Park, Y.; Kim, B.Y.; Kim, Y.D.; Yu, J.K.; Kim, Y.I.; Um, T.; Kim, S.; et al. Comparative Transcriptomics for Genes Related to Berberine and Berbamine Biosynthesis in Berberidaceae. Plants 2022, 11, 2676. [Google Scholar] [CrossRef]
- Lee, E.J.; Facchini, P. Norcoclaurine synthase is a member of the pathogenesis-related 10/Bet v1 protein family. Plant. Cell 2010, 22, 3489–3503. [Google Scholar] [CrossRef] [Green Version]
- Daniel, B.; Wallner, S.; Steiner, B.; Oberdorfer, G.; Kumar, P.; van der Graaff, E.; Roitsch, T.; Sensen, C.W.; Gruber, K.; Macheroux, P. Structure of a Berberine Bridge Enzyme-Like Enzyme with an Active Site Specific to the Plant Family Brassicaceae. PLoS ONE 2016, 11, e0156892. [Google Scholar] [CrossRef] [Green Version]
- Mattevi, A.; Fraaije, M.W.; Mozzarelli, A.; Olivi, L.; Coda, A.; van Berkel, W.J.H. Crystal structures and inhibitor binding in the octameric flavoenzyme vanillyl-alcohol oxidase: The shape of the active-site cavity controls substrate specificity. Structure 1997, 5, 907–920. [Google Scholar] [CrossRef] [Green Version]
- Liscombe, D.K.; Louie, G.V.; Noel, J.P. Architectures, mechanisms and molecular evolution of natural product methyltransferases. Nat. Prod. Rep. 2012, 29, 1238–1250. [Google Scholar] [CrossRef]
- Morris, J.S.; Facchini, P.J. Molecular Origins of Functional Diversity in Benzylisoquinoline Alkaloid Methyltransferases. Front. Plant. Sci. 2019, 10, 1058. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, R.K.; Bruneau, A.; Bantignies, B. Plant O-methyltransferases: Molecular analysis, common signature and classification. Plant. Mol. Biol. 1998, 36, 1–10. [Google Scholar] [CrossRef]
- Danielson, P.B. The cytochrome P450 superfamily: Biochemistry, evolution and drug metabolism in humans. Curr. Drug. Metab. 2002, 3, 561–597. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, M. Impacts of diversification of cytochrome P450 on plant metabolism. Biol. Pharm. Bull. 2012, 35, 824–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizutani, M.; Sato, F. Unusual P450 reactions in plant secondary metabolism. Arch. Biochem. Biophys. 2011, 507, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Schuler, M.A. Cytochrome P450 Genes from the Sacred Lotus Genome. Trop. Plant. Biol. 2013, 6, 138–151. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef]
- Hori, K.; Yamada, Y.; Purwanto, R.; Minakuchi, Y.; Toyoda, A.; Hirakawa, H.; Sato, F. Mining of the Uncharacterized Cytochrome P450 Genes Involved in Alkaloid Biosynthesis in California Poppy Using a Draft Genome Sequence. Plant. Cell. Physiol. 2018, 59, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Tanko, Y.; Suleiman, I.; Jimoh, A.; Musa, S.; Mohammed, K.; Salisu, A.; Yusuf, R. Anti-diabetic effect of ethanol leaf extract of Combretum micranthum blood glucose and oxidative stress biomarkers on alloxan induced diabetic in wistar rats. Bayero J. Pure Appl. Sci. 2017, 10, 108–114. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Order | Family | Common Name | Analyses * |
|---|---|---|---|---|
| Caulophyllum robustum | Ranunculales | Berberidaceae | Asian blue cohosh | 1, 2, 3, 4, 5 |
| Berberis koreana | Ranunculales | Berberidaceae | Barberry | 1, 2, 3, 4, 5 |
| Berberis lycium | Ranunculales | Berberidaceae | Barberry | 3, 4 |
| Berberis stoliniferum | Ranunculales | Berberidaceae | Barberry | 3 |
| Combretum micranthum | Myrtales | Combretaceae | Kinkeliba | 5 |
| Coptis chinensis | Ranunculales | Ranunculaceae | Chinese goldthread | 1, 2, 3, 4, 5 |
| Coptis japonica | Ranunculales | Ranunculaceae | Chinese goldthread | 5 |
| Thalictrum flavum | Ranunculales | Ranunculaceae | Common meadow-rue | 3, 4 |
| Papaver nudicaule | Ranunculales | Papaveraceae | Iceland poppy | 1, 3, 4, 5 |
| Papaver somniferum | Ranunculales | Papaveraceae | Opium poppy | 3, 4, 5 |
| Eschscholzia california | Ranunculales | Papaveraceae | California poppy | 3, 4, 5 |
| Macleaya cordata | Ranunculales | Papaveraceae | Plume puppy | 1, 3, 4, 5 |
| Nelumbo nucifera | Proteales | Nelumbonaceae | Sacred lotus | 1, 2, 3, 4, 5 |
| Cinnamon micranthum | Laurales | Lauraceae | Cinnamon | 5 |
| Fruit vs. Leaf | Leaf vs. Stem | Fruit vs. Stem | |
|---|---|---|---|
| Up | 761 | 326 | 536 |
| Down | 1135 | 105 | 768 |
| Enzymes | EC Numbers | C. robustum | B. koreana | N. nucifera | P. nudicaule | C. chinensis | M. cordata |
|---|---|---|---|---|---|---|---|
| TYDC | 4.1.1.25 | 1 | 2 | 5 | 10 | 5 | 12 |
| 3OHase | 1.14.16.2 | 1 | 2 | 1 | 4 | 3 | * |
| TyrAT | 2.6.1.5 | 2 | 1 | 8 | 3 | 3 | 4 |
| HPPDC | 4.1.1.80 | 1 | 2 | 5 | 10 | 5 | 12 |
| NCS | 4.2.1.78 | 10 | 11 | 5 | - | 6 | 1 |
| 6OMT | 2.1.1.128 | 8 | 7 | 9 | 3 | 3 | 1 |
| CNMT | 2.1.140 | 1 | 3 | 1 | 1 | 1 | 2 |
| NMCH/CYP80B3 | 1.14.13.71 | 1 | 5 | 4 | 3 | 1 | 1 |
| 4OMT | 2.1.1.16 | 1 | 1 | 4 | 5 | 1 | 1 |
| BBE | 1.21.3.3 | 4 | 5 | 4 | 3 | 5 | 5 |
| SOMT | 2.1.1.247 | 3 | 1 | 2 | 1 | 1 | 1 |
| CAS/CYP719A21 | 1.14.21.5 | 1 | 2 | 1 | 2 | 4 | 3 |
| STOX | 1.3.3.8 | 2 | 1 | 3 | - | 1 | 6 |
| Family | Number of Subfamilies | Genes (Number of Genes) | Number of Genes |
|---|---|---|---|
| CYP3 | 1 | CYP3a3 (1) | 1 |
| CYP6 | 1 | CYP6a2 (1) | 1 |
| CYP52 | 1 | CYP52A13 (1) | 1 |
| CYP71 | 11 | CYP71A1 (13), CYPA2 (2), CYP71A9 (1), CYPAU50 (4), CYP71A19 (1), VYP71B34 (1), CYPBE30 (1), CYP71D8 (1), CYPD9 (1), CYP71D11 (1), CYP71B10 (1) | 27 |
| CYP72 | 2 | CYP72A (8), CYP72A219 (14) | 22 |
| CYP76 | 1 | CYP76A1 (2) | 3 |
| CYP77 | 2 | CYP77A2 (1), CYP77A3 (1) | 2 |
| CYP78 | 3 | CYP78A5 (2), CYP78A7 (6), CYP78A9 (2) | 10 |
| CYP79 | 2 | CYP79A68 (1), CYP89A2 (1) | 2 |
| CYP80 | 1 | CYP80B3 (1) | 1 |
| CYP81 | 2 | CYP81Q32 (3), CYP89Q39 (1) | 4 |
| CYP82 | 2 | CYP82A3 (1), CYP82C4 (2), CYP82D47 (1) | 2 |
| CYP84 | 1 | CYP84A1 (3) | 3 |
| CYP85 | 1 | CYP85A (3) | 3 |
| CYP86 | 3 | CYP86A1 (1), CYP86A8 (1), CYP86B1 (2) | 4 |
| CYP87 | 1 | CYP87A3 (2) | 2 |
| CYP89 | 1 | CYP89A2 (3) | 3 |
| CYP90 | 2 | CYP90A1, CYP90B1 | 2 |
| CYP94 | 5 | CYP94A1 (1), CYP94A2 (6), CYP94B3 (2), CYP94C1 (2), CYP94C1 (1) | 12 |
| CYP97 | 1 | CYP97B2 | 1 |
| CYP98 | 1 | CYP98A2 (2) | 2 |
| CYP313 | 1 | CYP313a2 | 1 |
| CYP704 | 2 | CYP704B1 (1), CYP704C1 (3) | 4 |
| CYP709 | 1 | CYP709B2 (1) | 1 |
| CYP711 | 1 | CYP711A1 (4) | 4 |
| CYP714 | 1 | CYP714 (3) | 3 |
| CYP716 | 1 | CYP716B1 (4) | 4 |
| CYP719 | 1 | CYP719A1 (1) | 1 |
| CYP724 | 1 | CYP724B1 (3) | 3 |
| CYP734 | 1 | CYP734A1 (2) | 2 |
| CYP736 | 1 | CYP736A2 (1) | 1 |
| Not classified | 52 | ||
| Total | 56 | 184 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, N.-I.; Roy, N.S.; Park, Y.; Choi, B.-S.; Jeon, M.J.; Oh, J.Y.; Kim, B.-Y.; Kim, Y.-D.; Kim, Y.-I.; Um, T.; et al. Isolation and Characterization of the Genes Involved in the Berberine Synthesis Pathway in Asian Blue Cohosh, Caulophyllum robustum. Plants 2023, 12, 1483. https://doi.org/10.3390/plants12071483
Park N-I, Roy NS, Park Y, Choi B-S, Jeon MJ, Oh JY, Kim B-Y, Kim Y-D, Kim Y-I, Um T, et al. Isolation and Characterization of the Genes Involved in the Berberine Synthesis Pathway in Asian Blue Cohosh, Caulophyllum robustum. Plants. 2023; 12(7):1483. https://doi.org/10.3390/plants12071483
Chicago/Turabian StylePark, Nam-Il, Neha Samir Roy, Yeri Park, Beom-Soon Choi, Mi Jin Jeon, Ji Yeon Oh, Bo-Yun Kim, Young-Dong Kim, Yong-In Kim, Taeyoung Um, and et al. 2023. "Isolation and Characterization of the Genes Involved in the Berberine Synthesis Pathway in Asian Blue Cohosh, Caulophyllum robustum" Plants 12, no. 7: 1483. https://doi.org/10.3390/plants12071483