
Physiological and Morphological Responses of Blackberry Seedlings to Different Nitrogen Forms
 , ,
, ,
Abstract
:1. Introduction
2. Results
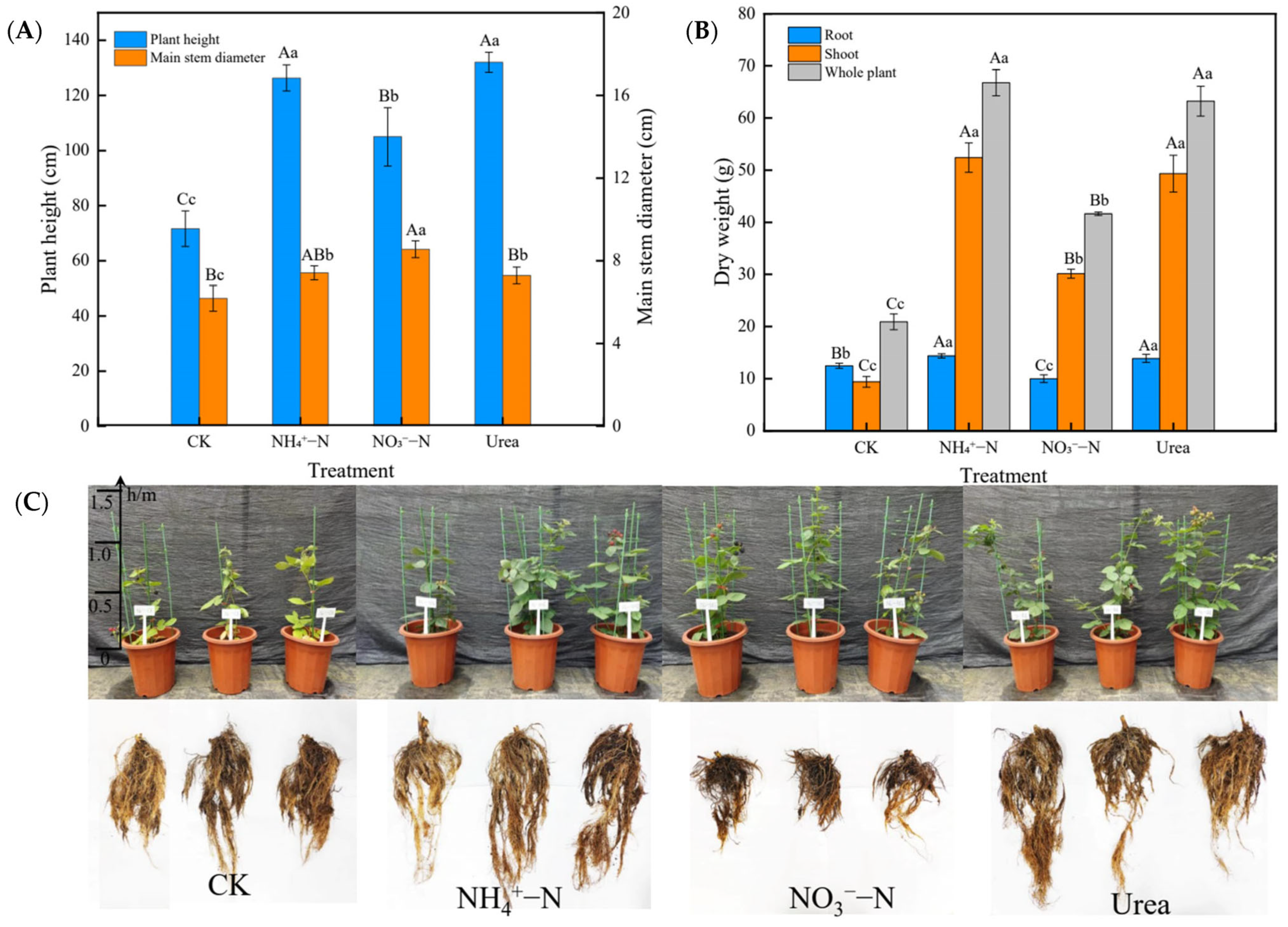
2.1. Growth Characteristics
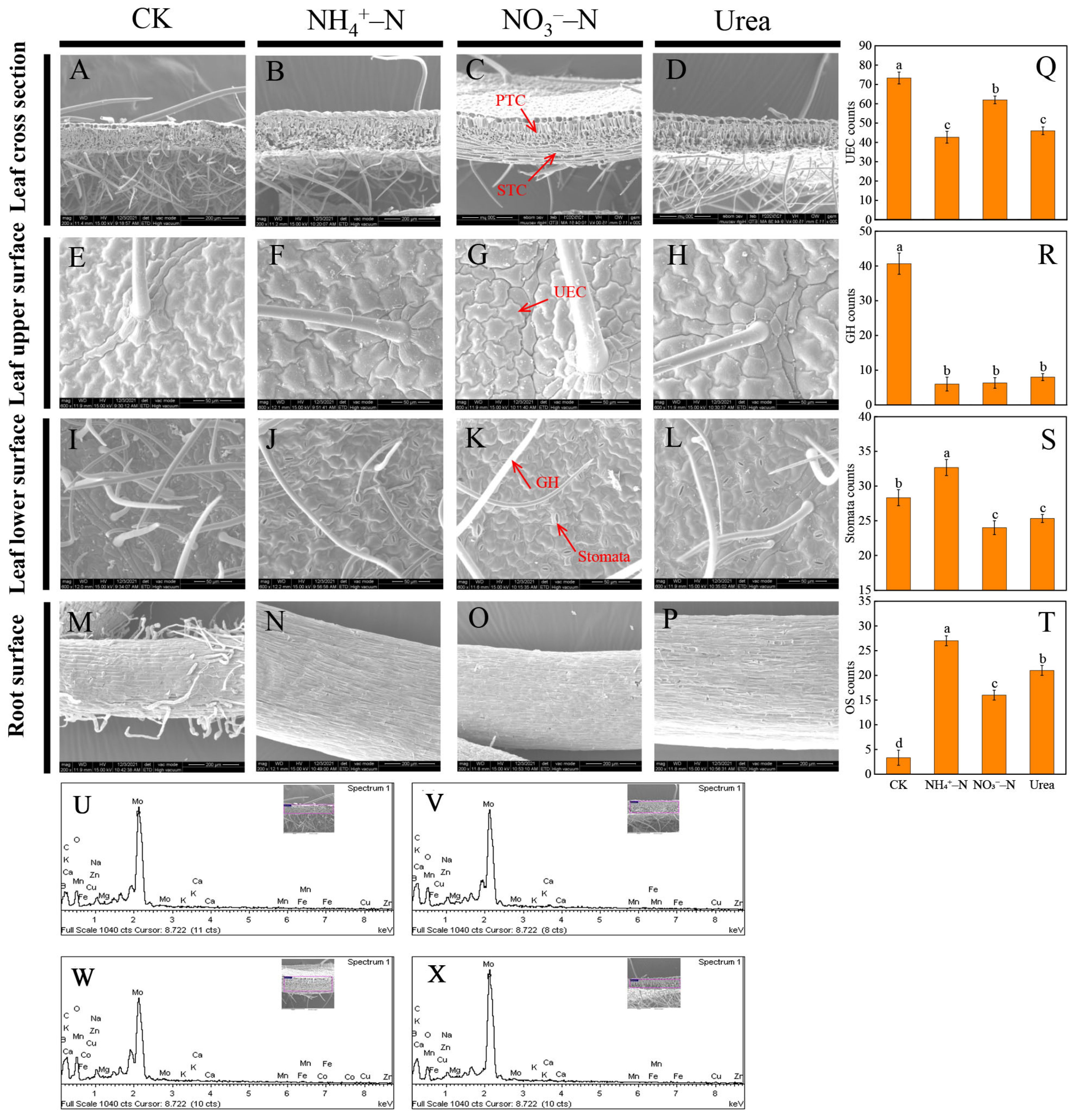
2.2. Morphological Changes
2.3. Cultivation Substrate Analysis
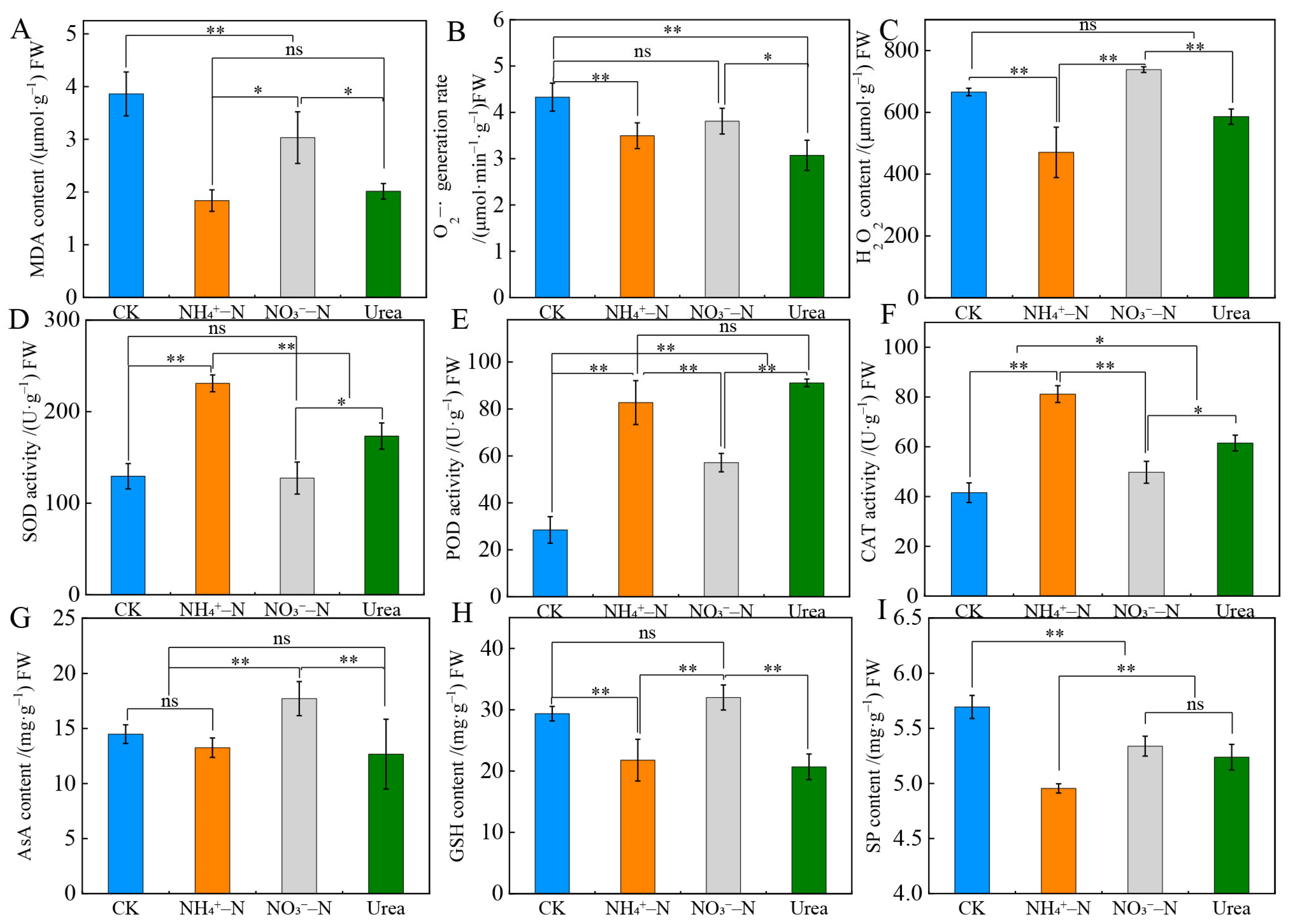
2.4. Root Physiology and Antioxidant Indicators
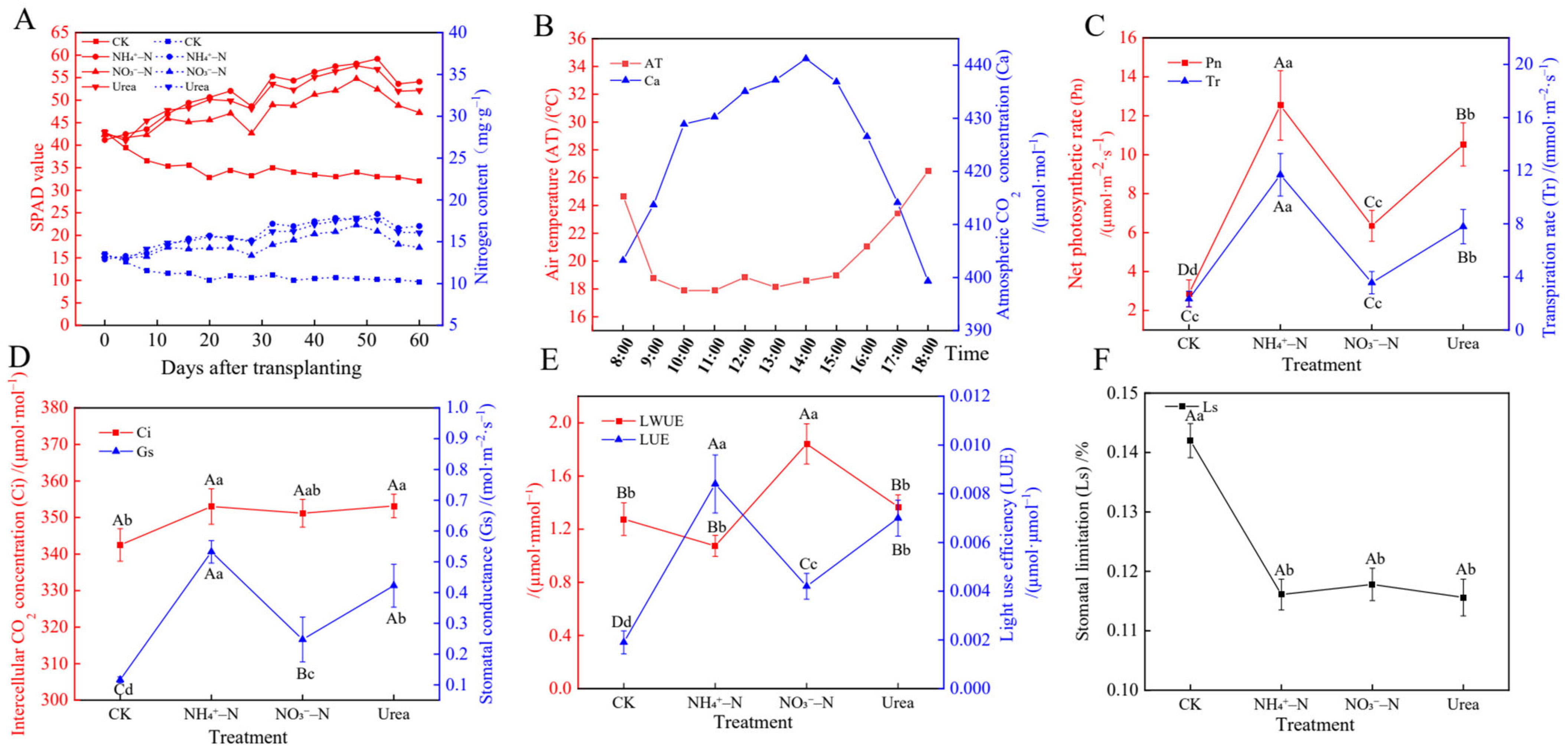
2.5. Photosynthesis and Gas Exchange
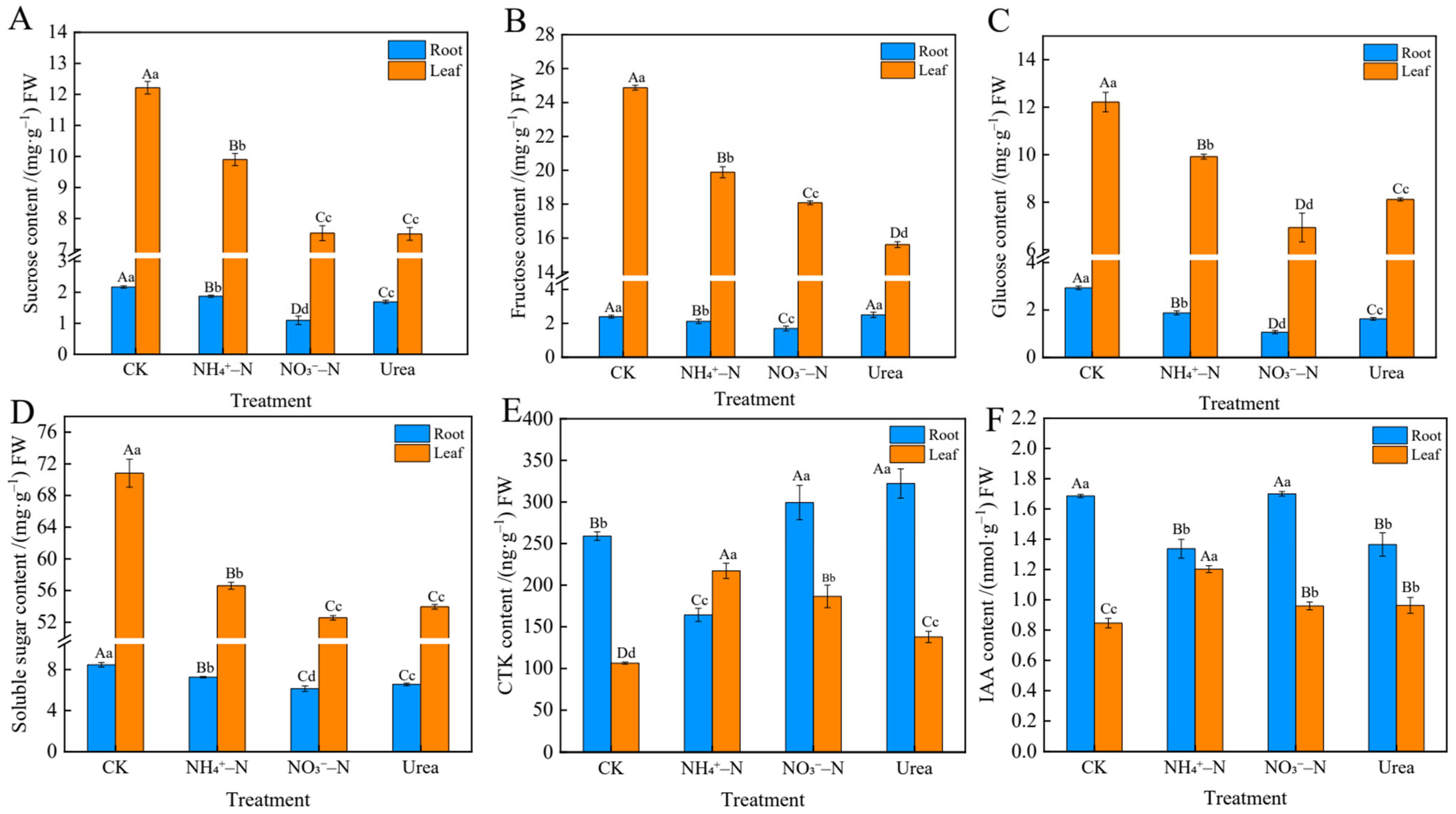
2.6. Sugar and Endogenous Hormone Levels in Roots and Leaves
2.7. Free Amino Acid Contents in Roots and Leaves
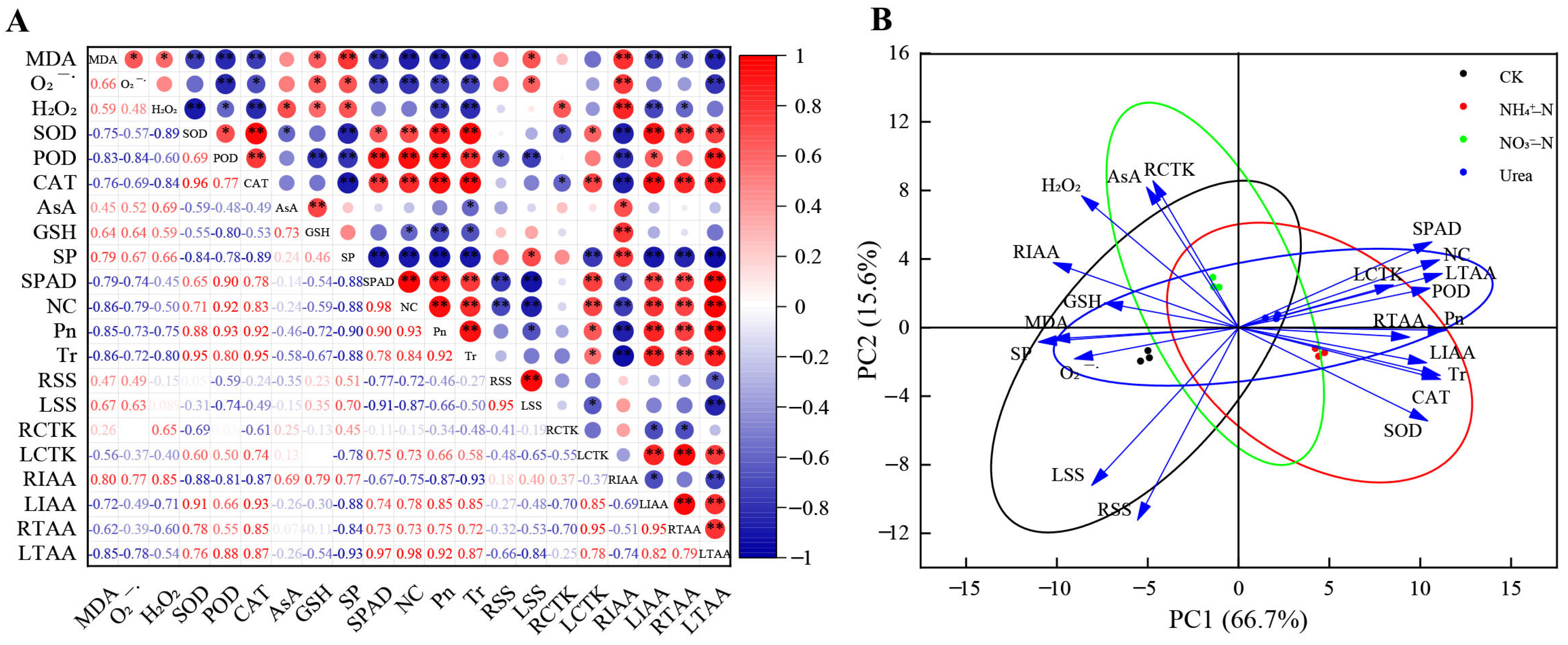
2.8. Correlation and PCA of Physiological Indexes with Different N Forms
2.9. Correlation between Growth Indicators and Principal Components
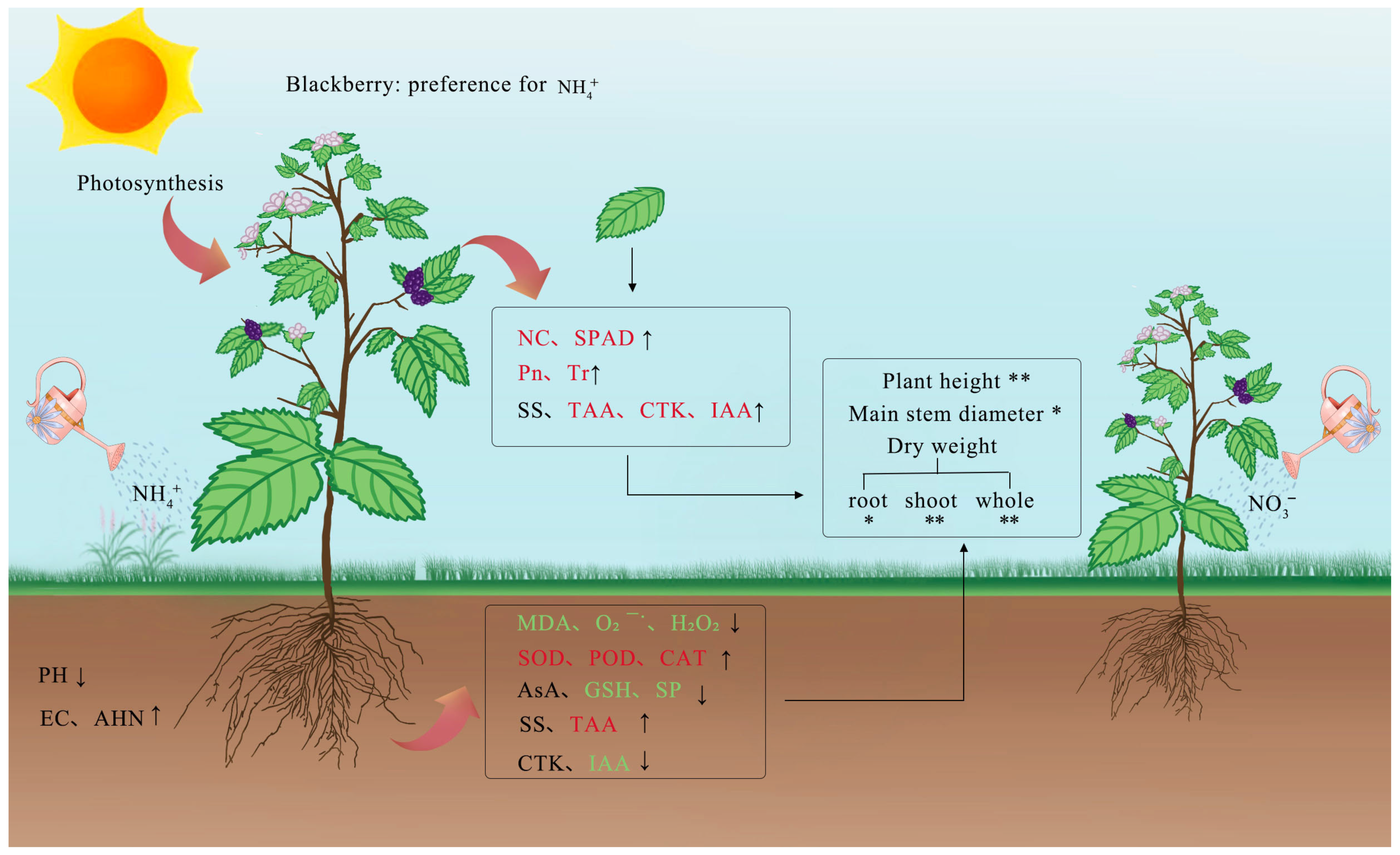
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Treatments
4.2. Assessment of Plant Growth, Biomass and Morphology
4.3. Determination of the Physical and Chemical Properties of the Cultivation Substrate
4.3.1. Measurement of pH and EC
4.3.2. Determination of Organic Matter
4.3.3. Determination of Alkali-Hydrolysable N
4.4. Measurement of Root Physiology and Antioxidant Indicators
4.5. Analysis of Photosynthetic Parameters
4.6. Determination of the Sugar and Endogenous Hormone Contents in Roots and Leaves
4.7. Assessment of Free Amino Acid Contents in Roots and Leaves
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, H.; Zhang, C.; Wu, W.; Li, W.; Wei, Y.; Dong, S. Physiological responses of blackberry cultivar ‘Ningzhi 1’ to drought stress. Russ. J. Plant Physiol. 2015, 62, 472–479. [Google Scholar] [CrossRef]
- Kaume, L.; Howard, L.R.; Devareddy, L. The blackberry fruit: A review on its composition and chemistry, metabolism and bioavailability, and health benefits. J. Agric. Food Chem. 2012, 60, 5716–5727. [Google Scholar] [CrossRef]
- Patel, A.; Rojas-Vera, J.; Dacke, C. Therapeutic constituents and actions of Rubus species. Curr. Med. Chem. 2004, 11, 1501–1512. [Google Scholar] [CrossRef]
- Strik, B.C.; Clark, J.R.; Finn, C.E.; Bañados, M.P. Worldwide blackberry production. HortTechnology 2007, 17, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Gan, P.; Chen, A. Environmental controls on soil pH in planted forest and its response to nitrogen deposition. Environ. Res. 2019, 172, 159–165. [Google Scholar] [CrossRef]
- Liu, S.; Chi, Q.; Cheng, Y.; Zhu, B.; Li, W.; Zhang, X.; Huang, Y.; Müller, C.; Cai, Z.; Zhang, J. Importance of matching soil N transformations, crop N form preference, and climate to enhance crop yield and reducing N loss. Sci. Total Environ. 2019, 657, 1265–1273. [Google Scholar] [CrossRef]
- Khalsa, S.D.S.; Smart, D.R.; Muhammad, S.; Armstrong, C.M.; Sanden, B.L.; Houlton, B.Z.; Brown, P.H. Intensive fertilizer use increases orchard N cycling and lowers net global warming potential. Sci. Total Environ. 2020, 722, 137889. [Google Scholar] [CrossRef]
- Alt, D.S.; Doyle, J.W.; Malladi, A. Nitrogen-source preference in blueberry (Vaccinium sp.): Enhanced shoot nitrogen assimilation in response to direct supply of nitrate. J. Plant Physiol. 2017, 216, 79–87. [Google Scholar] [CrossRef]
- Guo, L.; Meng, H.; Teng, K.; Fan, X.; Zhang, H.; Teng, W.; Yue, Y.; Wu, J. Effects of Nitrogen Forms on the Growth and Nitrogen Accumulation in Buchloe dactyloides Seedlings. Plants 2022, 11, 2086. [Google Scholar] [CrossRef]
- Tian, J.; Pang, Y.; Yuan, W.; Peng, J.; Zhao, Z. Growth and nitrogen metabolism in Sophora japonica (L.) as affected by salinity under different nitrogen forms. Plant Sci. 2022, 322, 111347. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Wirdnam, C.; Meier, S.; Komarova, N.Y.; Mimmo, T.; Cesco, S.; Rentsch, D.; Pinton, R. Isolation and functional characterization of a high affinity urea transporter from roots of Zea mays. BMC Plant Biol. 2014, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Hessini, K.; Kronzucker, H.J.; Abdelly, C.; Cruz, C. Drought stress obliterates the preference for ammonium as an N source in the C4 plant Spartina alterniflora. J. Plant Physiol. 2017, 213, 98–107. [Google Scholar] [CrossRef]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef]
- Huang, H.; Yao, Q.; Xia, E.; Gao, L. Metabolomics and transcriptomics analyses reveal nitrogen influences on the accumulation of flavonoids and amino acids in young shoots of tea plant (Camellia sinensis L.) associated with tea flavor. J. Agric. Food Chem. 2018, 66, 9828–9838. [Google Scholar] [CrossRef]
- Jampeetong, A.; Brix, H.; Kantawanichkul, S. Effects of inorganic nitrogen form on growth, morphology, N uptake, and nutrient allocation in hybrid Napier grass (Pennisetum purpureum × Pennisetum americanum cv. Pakchong1). Ecol. Eng. 2014, 73, 653–658. [Google Scholar] [CrossRef]
- Boschiero, B.N.; Mariano, E.; Azevedo, R.A.; Trivelin, P.C.O. Influence of nitrate-ammonium ratio on the growth, nutrition, and metabolism of sugarcane. Plant Physiol. Biochem. 2019, 139, 246–255. [Google Scholar] [CrossRef]
- Garnica, M.; Houdusse, F.; Yvin, J.C.; Garcia-Mina, J.M. Nitrate modifies urea root uptake and assimilation in wheat seedlings. J. Sci. Food Agric. 2009, 89, 55–62. [Google Scholar] [CrossRef]
- Borysiuk, K.; Ostaszewska-Bugajska, M.; Vaultier, M.-N.; Hasenfratz-Sauder, M.-P.; Szal, B. Enhanced formation of methylglyoxal-derived advanced glycation end products in Arabidopsis under ammonium nutrition. Front. Plant Sci. 2018, 9, 667. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Beatty, P.H.; Klein, M.S.; Fischer, J.J.; Lewis, I.A.; Muench, D.G.; Good, A.G. Understanding plant nitrogen metabolism through metabolomics and computational approaches. Plants 2016, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Cruz, J.L.; Alves, A.A.; LeCain, D.R.; Ellis, D.D.; Morgan, J.A. Effect of elevated CO2 concentration and nitrate: Ammonium ratios on gas exchange and growth of cassava (Manihot esculenta Crantz). Plant Soil 2014, 374, 33–43. [Google Scholar] [CrossRef]
- Zebarth, B.J.; Tai, H.; Luo, S.; Millard, P.; De Koeyer, D.; Li, X.-Q.; Xiong, X. Effect of nitrogen form on gene expression in leaf tissue of greenhouse grown potatoes during three stages of growth. Am. J. Potato Res. 2012, 89, 315–327. [Google Scholar] [CrossRef]
- Podgorska, A.; Gieczewska, K.; Łukawska-Kuźma, K.; Rasmusson, A.G.; Gardeström, P.; Szal, B. Long-term ammonium nutrition of A rabidopsis increases the extrachloroplastic NAD (P) H/NAD (P)+ ratio and mitochondrial reactive oxygen species level in leaves but does not impair photosynthetic capacity. Plant Cell Environ. 2013, 36, 2034–2045. [Google Scholar]
- Vuletić, M.; Hadži-Tašković Šukalović, V.; Marković, K.; Dragišić Maksimović, J. Antioxidative system in maize roots as affected by osmotic stress and different nitrogen sources. Biol. Plant. 2010, 54, 530–534. [Google Scholar] [CrossRef]
- Carr, N.F.; Boaretto, R.M.; Mattos, D., Jr. Coffee seedlings growth under varied NO3−: NH4+ ratio: Consequences for nitrogen metabolism, amino acids profile, and regulation of plasma membrane H+-ATPase. Plant Physiol. Biochem. 2020, 154, 11–20. [Google Scholar] [CrossRef]
- Penella, C.; Nebauer, S.G.; San Bautista, A.; López-Galarza, S.; Calatayud, Á. Rootstock alleviates PEG-induced water stress in grafted pepper seedlings: Physiological responses. J. Plant Physiol. 2014, 171, 842–851. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.-M.; Lu, Y.-T.; Qiu, Q.-L.; Fan, D.-M.; Wang, X.-C.; Zheng, X.-Q. Influence of different nitrogen sources on carbon and nitrogen metabolism and gene expression in tea plants (Camellia sinensis L.). Plant Physiol. Biochem. 2021, 167, 561–566. [Google Scholar] [CrossRef]
- Forde, B.G. The role of long-distance signalling in plant responses to nitrate and other nutrients. J. Exp. Bot. 2002, 53, 39–43. [Google Scholar]
- Samuelson, M.E.; Larsson, C.-M. Nitrate regulation of zeation riboside levels in barley roots: Effects of inhibitors of N assimilation and comparison with ammonium. Plant Sci. 1993, 93, 77–84. [Google Scholar] [CrossRef]
- Tian, Q.; Chen, F.; Liu, J.; Zhang, F.; Mi, G. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots. J. Plant Physiol. 2008, 165, 942–951. [Google Scholar] [CrossRef]
- Mercier, H.; Kerbauy, G.; Sotta, B.; Miginiac, E. Effects of NO3−, NH4+ and urea nutrition on endogenous levels of IAA and four cytokinins in two epiphytic bromeliads. Plant Cell Environ. 1997, 20, 387–392. [Google Scholar] [CrossRef]
- Strik, B.C.; Bryla, D.R. Uptake and partitioning of nutrients in blackberry and raspberry and evaluating plant nutrient status for accurate assessment of fertilizer requirements. HortTechnology 2015, 25, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.M.; Wilkerson, F.P.; Dugdale, R.C.; Raven, J.A.; Dupont, C.L.; Leavitt, P.R.; Parker, A.E.; Burkholder, J.M.; Kana, T.M. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 2016, 61, 165–197. [Google Scholar] [CrossRef]
- Moschou, P.N.; Wu, J.; Cona, A.; Tavladoraki, P.; Angelini, R.; Roubelakis-Angelakis, K.A. The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 2012, 63, 5003–5015. [Google Scholar] [CrossRef] [Green Version]
- Camalle, M.; Standing, D.; Jitan, M.; Muhaisen, R.; Bader, N.; Bsoul, M.; Ventura, Y.; Soltabayeva, A.; Sagi, M. Effect of salinity and nitrogen sources on the leaf quality, biomass, and metabolic responses of two ecotypes of Portulaca oleracea. Agronomy 2020, 10, 656. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Sun, S.; Liu, W.; Zhu, L.; Yan, X. Different Forms and Proportions of Exogenous Nitrogen Promote the Growth of Alfalfa by Increasing Soil Enzyme Activity. Plants 2022, 11, 1057. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.H.; Hulme, S.P.; Rees, M.; Ripley, B.S.; Woodward, F.I.; Osborne, C.P. Ecophysiological traits in C3 and C4 grasses: A phylogenetically controlled screening experiment. New Phytol. 2010, 185, 780–791. [Google Scholar] [CrossRef]
- Guerrero, M.G.; Vega, J.M.; Losada, M. The assimilatory nitrate-reducing system and its regulation. Annu. Rev. Plant Biol. 1981, 32, 169–204. [Google Scholar] [CrossRef]
- Esteban, R.; Royo, B.; Urarte, E.; Zamarreño, A.; Garcia-Mina, J.; Moran, J. Both free indole-3-acetic acid and the photosynthetic efficiency play a relevant role in the response of Medicago truncatula to urea and ammonium nutrition under axenic conditions. Front. Plant Sci. 2016, 7, 00140. [Google Scholar] [CrossRef] [Green Version]
- Grechi, I.; Vivin, P.; Hilbert, G.; Milin, S.; Robert, T.; Gaudillère, J. Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Environ. Exp. Bot. 2007, 59, 139–149. [Google Scholar] [CrossRef]
- Dickson, R.W.; Fisher, P.R.; Argo, W.R.; Jacques, D.J.; Sartain, J.B.; Trenholm, L.E.; Yeager, T.H. Solution ammonium: Nitrate ratio and cation/anion uptake affect acidity or basicity with floriculture species in hydroponics. Sci. Hortic. 2016, 200, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Heil, J.; Vereecken, H.; Brüggemann, N. A review of chemical reactions of nitrification intermediates and their role in nitrogen cycling and nitrogen trace gas formation in soil. Eur. J. Soil Sci. 2016, 67, 23–39. [Google Scholar] [CrossRef]
- Xiao, W.-L.; Wang, Z.-X.; Wu, F.-Z.; Zhou, X.-G. Effects of soil improvement technology on soil quality in solar greenhouse. Environ. Sci. Pollut. Res. 2018, 25, 24093–24100. [Google Scholar] [CrossRef]
- Yan, L.P.; Liu, C.L.; Liang, H.M.; Mao, X.H.; Wang, F.; Pang, C.H.; Shu, J.; Xia, Y. Physiological responses to salt stress of T2 alfalfa progenies carrying a transgene for betaine aldehyde dehydrogenase. Plant Cell Tissue Organ Cult. 2012, 108, 191–199. [Google Scholar] [CrossRef]
- Hessini, K.; Jeddi, K.; Siddique, K.H.M.; Moran, J.F. Can nitrate-based fertilization be recommended for the cultivation of ammonium-preferring species in a salty ecosystem? The case for Spartina alterniflora. Arab. J. Geosci. 2021, 14, 1259. [Google Scholar] [CrossRef]
- Hessini, K. Nitrogen form differently modulates growth, metabolite profile, and antioxidant and nitrogen metabolism activities in roots of Spartina alterniflora in response to increasing salinity. Plant Physiol. Biochem. 2022, 174, 35–42. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Modulation of antioxidant defense system is associated with combined drought and heat stress tolerance in citrus. Front. Plant Sci. 2017, 8, 953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanu, A.S.; Ashraf, U.; Mo, Z.; Sabir, S.-u.-R.; Baggie, I.; Charley, C.S.; Tang, X. Calcium amendment improved the performance of fragrant rice and reduced metal uptake under cadmium toxicity. Environ. Sci. Pollut. Res. 2019, 26, 24748–24757. [Google Scholar] [CrossRef]
- Hassanein, R.A.; El Khawas, S.A.; Khafaga, H.S.; Abd El-Nabe, A.S.; Abd Elrady, A. Amelioration of drought stress on physiological performance of pearl millet (Pennisetum americanum) plant grown under saline condition using potassium humate and silicon source. Egypt. J. Exp. Biol. 2017, 13, 57–68. [Google Scholar] [CrossRef]
- Wu, Z.; Jiang, Q.; Yan, T.; Zhang, X.; Xu, S.; Shi, H.; Deng, T.; Li, F.; Du, Y.; Du, R. Ammonium nutrition mitigates cadmium toxicity in rice (Oryza sativa L.) through improving antioxidase system and the glutathione-ascorbate cycle efficiency. Ecotoxicol. Environ. Saf. 2020, 189, 110010. [Google Scholar] [CrossRef]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Reddy, K.R.; Kakani, V.G.; Reddy, V. Nitrogen deficiency effects on plant growth, leaf photosynthesis, and hyperspectral reflectance properties of sorghum. Eur. J. Agron. 2005, 22, 391–403. [Google Scholar] [CrossRef]
- Guo, S.; Brück, H.; Sattelmacher, B. Effects of supplied nitrogen form on growth and water uptake of French bean (Phaseolus vulgaris L.) plants. Plant Soil 2002, 239, 267–275. [Google Scholar] [CrossRef]
- Raab, T.K.; Terry, N. Carbon, nitrogen, and nutrient interactions in Beta vulgaris L. as influenced by nitrogen source, NO3-versus NH4+. Plant Physiol. 1995, 107, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Kazuto, S.; Hans, L.; Kyozo, C. Change in Uptake, Transport and Accumulation of Ions in Nerium oleander (Rosebay) as Affected by Different Nitrogen Sources and Salinity. Ann. Bot. 2008, 102, 735–746. [Google Scholar]
- Ottow, E.A.; Brinker, M.; Teichmann, T.; Fritz, E.; Kaiser, W.; Brosché, M.; Kangasjarvi, J.; Jiang, X.; Polle, A. Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress. Plant Physiol. 2005, 139, 1762–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann. Bot. 2007, 99, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakakibara, H.; Takei, K.; Hirose, N. Interactions between nitrogen and cytokinin in the regulation of metabolism and development. Trends Plant Sci. 2006, 11, 440–448. [Google Scholar] [CrossRef]
- Kiba, T.; Kudo, T.; Kojima, M.; Sakakibara, H. Hormonal control of nitrogen acquisition: Roles of auxin, abscisic acid, and cytokinin. J. Exp. Bot. 2011, 62, 1399–1409. [Google Scholar] [CrossRef]
- Meier, M.; Liu, Y.; Lay-Pruitt, K.S.; Takahashi, H.; von Wirén, N. Auxin-mediated root branching is determined by the form of available nitrogen. Nat. Plants 2020, 6, 1136–1145. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S.; Arif, M.S.; Kausar, R. Nitrogen nutrition and adaptation of glycophytes to saline environment: A review. Arch. Agron. Soil Sci. 2018, 64, 1181–1206. [Google Scholar] [CrossRef]
- Deng, W.-W.; Wang, S.; Chen, Q.; Zhang, Z.-Z.; Hu, X.-Y. Effect of salt treatment on theanine biosynthesis in Camellia sinensis seedlings. Plant Physiol. Biochem. 2012, 56, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Cao, X.; Bai, Z.; Zhang, J.; Zhu, L.; Huang, J.; Jin, Q. Nitrogen metabolism correlates with the acclimation of photosynthesis to short-term water stress in rice (Oryza sativa L.). Plant Physiol. Biochem. 2018, 125, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Caspersen, S.; Svensson, B.; Hkansson, T.; Winter, C.; Asp, H. Blueberry—Soil interactions from an organic perspective. Sci. Hortic. 2016, 208, 78–91. [Google Scholar] [CrossRef]
- Nemie-Feyissa, D.; Olafsdottir, S.M.; Heidari, B.; Lillo, C. Nitrogen depletion and small R3-MYB transcription factors affecting anthocyanin accumulation in Arabidopsis leaves. Phytochemistry 2014, 98, 34–40. [Google Scholar] [CrossRef]
- Alcântara, B.K.; Rizzi, V.; Gaziola, S.A.; Azevedo, R.A. Soluble amino acid profile, mineral nutrient and carbohydrate content of maize kernels harvested from plants submitted to ascorbic acid seed priming. An. Acad. Bras. Cienc. 2017, 89, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, P.; Huang, F.L.; Guo, Z.H.; Xiao, X.Y.; Peng, C. Physiological responses of Morus alba L. in heavy metal(loid)-contaminated soil and its associated improvement of the microbial diversity. Environ. Sci. Pollut. Res. 2020, 27, 4294–4308. [Google Scholar] [CrossRef]
- Wang, A.G.; Luo, G.H. Quantitative Relation between the Reaction of Hydroxylamine and Superoxide Anion Radicals in Plants. Plant Physiol. Commun. 1990, 84, 2895–2898. [Google Scholar]
- Dhindsa, R.S.; Pamela, P.D.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Li, H.-X.; Xiao, Y.; Cao, L.-L.; Yan, X.; Li, C.; Shi, H.-Y.; Wang, J.-W.; Ye, Y.-H. Cerebroside C increases tolerance to chilling injury and alters lipid composition in wheat roots. PLoS ONE 2013, 8, e73380. [Google Scholar] [CrossRef] [Green Version]
- Asami, D.K.; Hong, Y.J.; Barrett, D.M.; Mitchell, A.E. Comparison of the total phenolic and ascorbic acid content of freeze-dried and air-dried marionberry, strawberry, and corn grown using conventional, organic, and sustainable agricultural practices. J. Agric. Food Chem. 2003, 51, 1237–1241. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol. 1985, 113, 548–555. [Google Scholar] [PubMed]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Potein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Maghsoudlou, P. Enzyme-linked immunosorbent assay (ELISA): The basics. Br. J. Hosp. Med. 2016, 77, C98–C101. [Google Scholar] [CrossRef]
- Zhou, M.; Hua, T.; Ma, X.; Sun, H.; Xu, L. Protein content and amino acids profile in 10 cultivars of ginkgo (Ginkgo biloba L.) nut from China. R. Soc. Open Sci. 2019, 6, 181571. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CK | NH4+–N | NO3−–N | Urea | |||||
|---|---|---|---|---|---|---|---|---|
| Element | Weight% | Atomic% | Weight% | Atomic% | Weight% | Atomic% | Weight% | Atomic% |
| C | 37.21 | 45.92 | 35.26 | 44.64 | 39 | 49.57 | 38.98 | 50.21 |
| O | 51.33 | 47.55 | 51.17 | 48.64 | 45.72 | 43.63 | 43.76 | 42.31 |
| Na | 8.35 | 5.38 | 5.82 | 3.85 | 6.82 | 4.53 | 6.28 | 4.22 |
| Mg | 0.65 | 0.4 | 1.27 | 0.79 | − | − | 0.53 | 0.34 |
| K | − | − | − | − | − | − | 0.59 | 0.23 |
| Ca | 0.98 | 0.36 | 3.45 | 1.31 | 0.89 | 0.34 | 1.74 | 0.67 |
| Mn | 0.9 | 0.24 | 1.32 | 0.37 | - | - | - | - |
| Fe | 0.44 | 0.12 | − | − | 2.65 | 0.73 | 1.63 | 0.45 |
| Cu | 0.14 | 0.03 | − | − | 1.57 | 0.41 | 4.05 | 0.99 |
| Zn | − | − | 1.71 | 0.4 | 2.1 | 0.5 | 2.44 | 0.58 |
| Mo | − | − | − | − | 1.25 | 0.29 | − | − |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Treatment | pH | EC (mS/cm) | SOM (%) | SOC (%) | AHN (mg/kg) |
|---|---|---|---|---|---|
| CK | 5.37 ± 0.01 b | 0.96 ± 0.01 c | 74.02 ± 0.70 b | 42.93 ± 0.41 b | 402.27 ± 14.09 d |
| NH₄+–N | 4.45 ± 0.01 d | 1.83 ± 0.02 a | 72.26 ± 1.39 b | 41.91 ± 0.80 b | 571.20 ± 12.83 a |
| NO₃−–N | 6.25 ± 0.01 a | 0.97 ± 0.017 c | 72.02 ± 1.20 b | 41.78 ± 0.69 b | 481.60 ± 2.80 c |
| Urea | 5.00 ± 0.02 c | 1.25 ± 0.07 b | 80.72 ± 0.68 a | 46.82 ± 0.39 a | 522.67 ± 9.00 b |
| Root/mg·(100 g)−1 DW | Leaf/mg·(100 g)−1 DW | |||||||
|---|---|---|---|---|---|---|---|---|
| Amino Acid | CK | NH₄+–N | NO₃−–N | Urea | CK | NH₄+–N | NO₃−–N | Urea |
| Asp | 1.30 ± 0.11 c | 1.82 ± 0.02 b | 2.33 ± 0.04 a | 1.40 ± 0.09 c | 1.22 ± 0.18 d | 6.81 ± 0.55 a | 4.53 ± 0.40 b | 3.86 ± 0.17 c |
| Thr | 0.91 ± 0.08 c | 1.23 ± 0.05 b | 1.52 ± 0.01 a | 0.84 ± 0.11 c | 1.00 ± 0.15 c | 3.24 ± 0.38 a | 2.39 ± 0.32 b | 3.36 ± 0.12 a |
| Ser | 5.08 ± 0.45 c | 11.46 ± 0.65 a | 11.28 ± 0.10 a | 7.77 ± 0.78 b | 1.48 ± 0.15 d | 19.80 ± 1.99 a | 10.20 ± 0.97 c | 15.57 ± 0.39 b |
| Glu | 1.56 ± 0.16 c | 1.92 ± 0.06 b | 3.60 ± 0.18 a | 1.82 ± 0.19 bc | 1.99 ± 0.08 b | 2.29 ± 0.25 ab | 2.75 ± 0.24 a | 2.31 ± 0.12 ab |
| Gly | 0.94 ± 0.18 a | 0.51 ± 0.06 b | 0.66 ± 0.13 b | 0.46 ± 0.08 b | 0.30 ± 0.06 c | 0.92 ± 0.05 a | 0.39 ± 0.03 c | 0.68 ± 0.03 b |
| Ala | 2.32 ± 0.19 c | 2.93 ± 0.08 b | 3.47 ± 0.01 a | 2.10 ± 0.23 c | 4.04 ± 0.42 d | 16.30 ± 1.31 a | 10.78 ± 1.19 c | 13.63 ± 0.42 b |
| Cys | 0.08 ± 0.01 b | 0.16 ± 0.01 a | 0.17 ± 0.01 a | 0.15 ± 0.02 a | 0.08 ± 0.01 c | 0.66 ± 0.03 a | 0.40 ± 0.03 b | 0.64 ± 0.04 a |
| Val | 0.76 ± 0.07 d | 1.48 ± 0.08 b | 1.80 ± 0.02 a | 1.01 ± 0.08 c | 1.34 ± 0.18 c | 6.10 ± 0.77 a | 5.05 ± 0.26 b | 6.02 ± 0.18 a |
| Met | 0.15 ± 0.03 c | 0.38 ± 0.01 a | 0.33 ± 0.02 b | 0.16 ± 0.01 c | 0.11 ± 0.01 c | 0.88 ± 0.12 a | 0.57 ± 0.06 b | 0.71 ± 0.01 b |
| Ile | 0.45 ± 0.03 c | 0.93 ± 0.04 a | 1.05 ± 0.00 a | 0.57 ± 0.08 b | 1.11 ± 0.14 | 5.12 ± 0.46 | 3.7 ± 0.64 | 5.03 ± 0.13 |
| Leu | 0.33 ± 0.04 c | 0.60 ± 0.05 b | 1.00 ± 0.05 a | 0.48 ± 0.11 bc | 1.24 ± 0.16 c | 5.40 ± 0.50 a | 4.34 ± 0.33 b | 5.59 ± 1.18 a |
| Tyr | 0.74 ± 0.09 a | 0.60 ± 0.05 b | 0.78 ± 0.02 a | 0.50 ± 0.03 b | 1.39 ± 0.05 c | 4.96 ± 0.32 a | 4.09 ± 0.44 b | 3.97 ± 0.09 b |
| Phe | 1.77 ± 0.01 b | 1.28 ± 0.18 d | 2.09 ± 0.01 a | 1.53 ± 0.11 c | 3.37 ± 0.18 c | 10.21 ± 0.82 b | 12.49 ± 1.06 a | 11.99 ± 0.35 ab |
| His | 1.72 ± 0.02 c | 3.59 ± 0.08 a | 2.36 ± 0.06 b | 1.73 ± 0.13 c | 1.45 ± 0.15 c | 5.89 ± 0.42 a | 4.34 ± 0.47 b | 4.44 ± 0.36 b |
| Lys | 0.38 ± 0.01 d | 0.88 ± 0.02 a | 0.73 ± 0.02 b | 0.51 ± 0.05 c | 0.66 ± 0.13 c | 2.63 ± 0.27 ab | 2.53 ± 0.20 b | 3.07 ± 0.10 a |
| Arg | 3.61 ± 0.02 d | 34.61 ± 1.26 a | 8.1 ± 0.35 c | 11.08 ± 1.34 b | 0.39 ± 0.04 d | 3.39 ± 0.37 a | 1.74 ± 0.19 c | 2.64 ± 0.12 b |
| Pro | 1.08 ± 0.13 c | 1.64 ± 0.10 b | 2.2 ± 0.15 a | 1.02 ± 0.15 c | 1.31 ± 0.17 d | 13.82 ± 0.59 a | 6.28 ± 0.50 c | 8.50 ± 0.23 b |
| ∑EAA | 6.47 ± 0.28 b | 10.37 ± 0.16 a | 10.86 ± 0.12 a | 6.84 ± 0.46 b | 10.27 ± 1.02 c | 39.26 ± 2.21 a | 35.41 ± 2.65 b | 40.21 ± 1.23 a |
| ∑NEAA | 16.71 ± 1.14 d | 55.84 ± 0.74 a | 32.58 ± 0.67 b | 26.29 ± 2.78 c | 12.20 ± 1.12 d | 68.94 ± 4.79 a | 41.15 ± 3.74 c | 51.78 ± 1.26 b |
| ∑TAA | 23.17 ± 1.47 d | 66.01 ± 0.78 a | 43.45 ± 0.59 b | 33.13 ± 3.08 c | 22.46 ± 1.97 d | 108.20 ± 7.02 a | 76.56 ± 4.09 c | 91.99 ± 2.27 b |
| Trait | Component | |
|---|---|---|
| 1 | 2 | |
| MDA | −0.88 ** | −0.05 |
| O2·− | −0.77 ** | −0.14 |
| H2O2 | −0.74 ** | 0.60 |
| SOD | 0.89 ** | −0.42 |
| POD | 0.90 ** | 0.18 |
| CAT | 0.95 ** | −0.23 |
| AsA | −0.43 | 0.64 ** |
| GSH | −0.63 ** | 0.11 |
| SP | −0.94 ** | −0.06 |
| SPAD | 0.91 ** | 0.39 |
| NC | 0.95 ** | 0.31 |
| Pn | 0.98 ** | −0.01 |
| Tr | 0.95 ** | −0.22 |
| RSS | −0.48 | −0.87 ** |
| LSS | −0.69 | −0.72 ** |
| RCTK | −0.40 | 0.66 ** |
| LCTK | 0.73 ** | 0.19 |
| RIAA | −0.88 ** | 0.30 |
| LIAA | 0.88 ** | −0.15 |
| RTAA | 0.81 ** | −0.04 |
| LTAA | 0.96 ** | 0.25 |
| Total | 14.01 | 3.28 |
| % of variance | 66.72 | 15.64 |
| Cumulative % | 66.72 | 82.36 |
| Index | PC1 | PC2 | Plant Height | Main Stem Diameter | Root Dry Weight | Shoot Dry Weight | Whole Dry Weight |
|---|---|---|---|---|---|---|---|
| PC1 | 1 | 0 | 0.888 ** | 0.588 * | 0.556 * | 0.863 ** | 0.837 ** |
| PC2 | 0 | 1 | 0.353 | 0.057 | 0.770 ** | 0.476 | 0.517 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, Y.; Yang, H.; Yang, H.; Wei, Z.; Che, J.; Wu, W.; Lyu, L.; Li, W. Physiological and Morphological Responses of Blackberry Seedlings to Different Nitrogen Forms. Plants 2023, 12, 1480. https://doi.org/10.3390/plants12071480
Duan Y, Yang H, Yang H, Wei Z, Che J, Wu W, Lyu L, Li W. Physiological and Morphological Responses of Blackberry Seedlings to Different Nitrogen Forms. Plants. 2023; 12(7):1480. https://doi.org/10.3390/plants12071480
Chicago/Turabian StyleDuan, Yongkang, Haiyan Yang, Hao Yang, Zhiwen Wei, Jilu Che, Wenlong Wu, Lianfei Lyu, and Weilin Li. 2023. "Physiological and Morphological Responses of Blackberry Seedlings to Different Nitrogen Forms" Plants 12, no. 7: 1480. https://doi.org/10.3390/plants12071480