
Growth and Leaf Gas Exchange Upregulation by Elevated [CO2] Is Light Dependent in Coffee Plants

 , , ,
, , ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
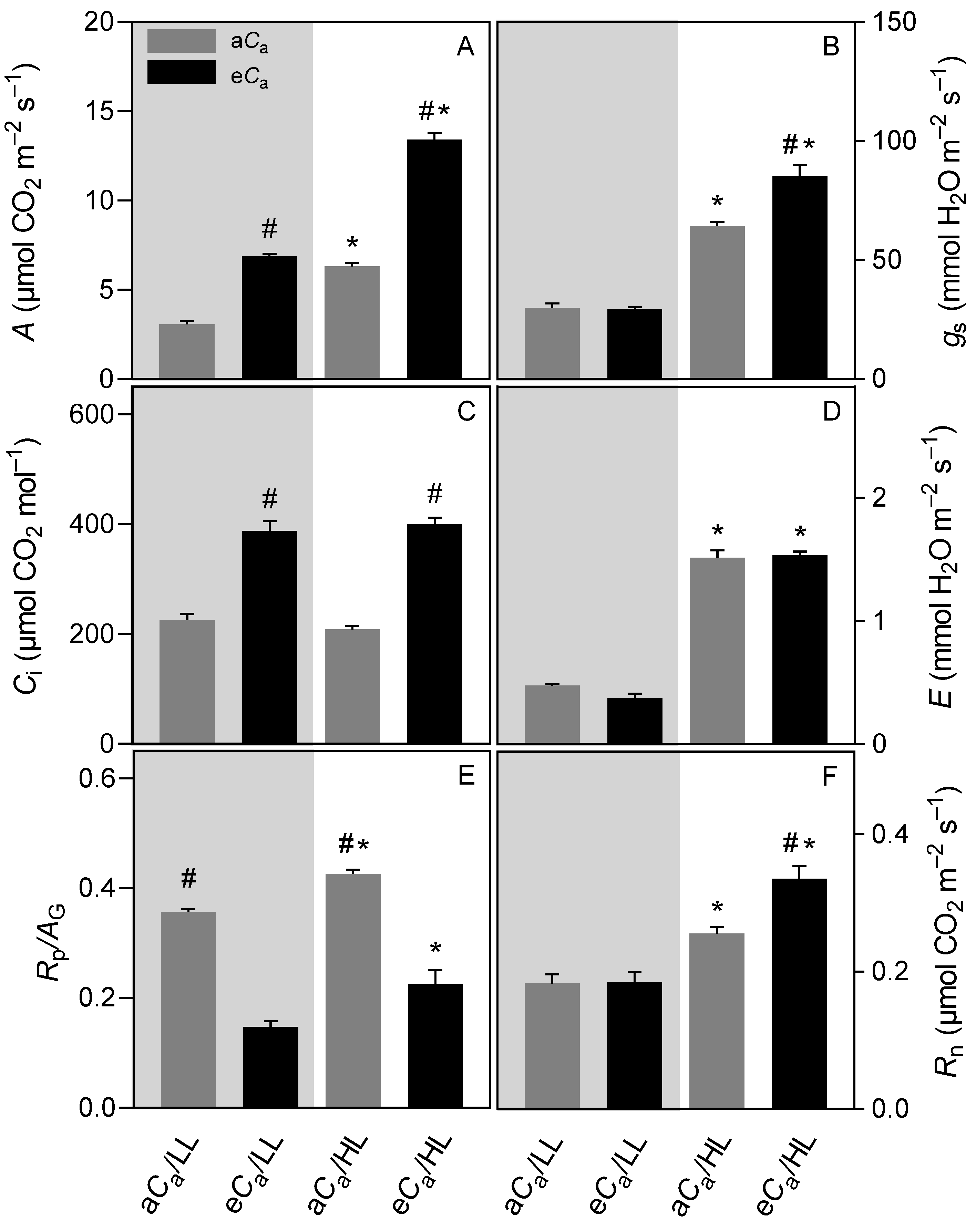
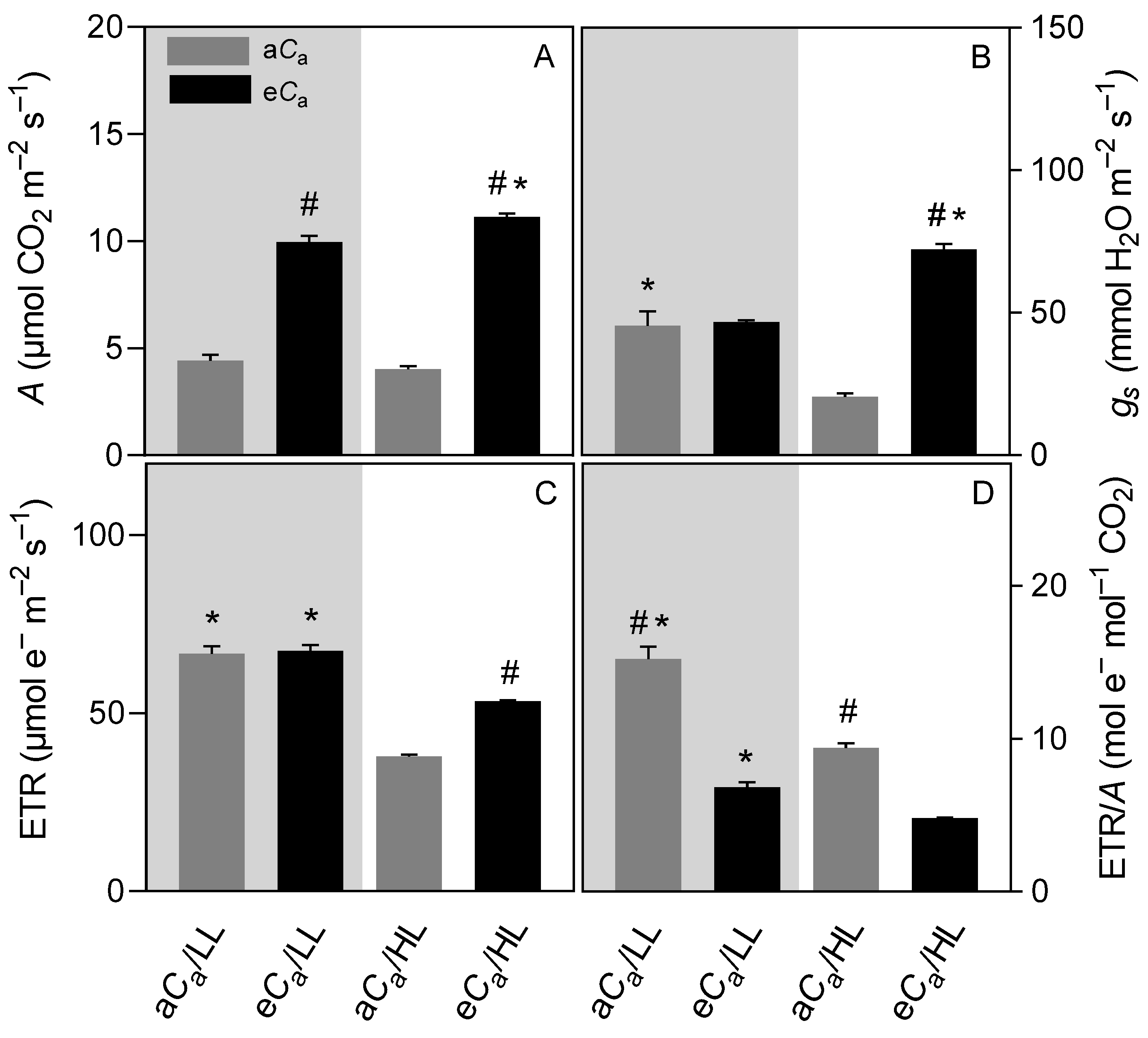
2.1. Gas Exchanges and Chlorophyll a Fluorescence Parameters
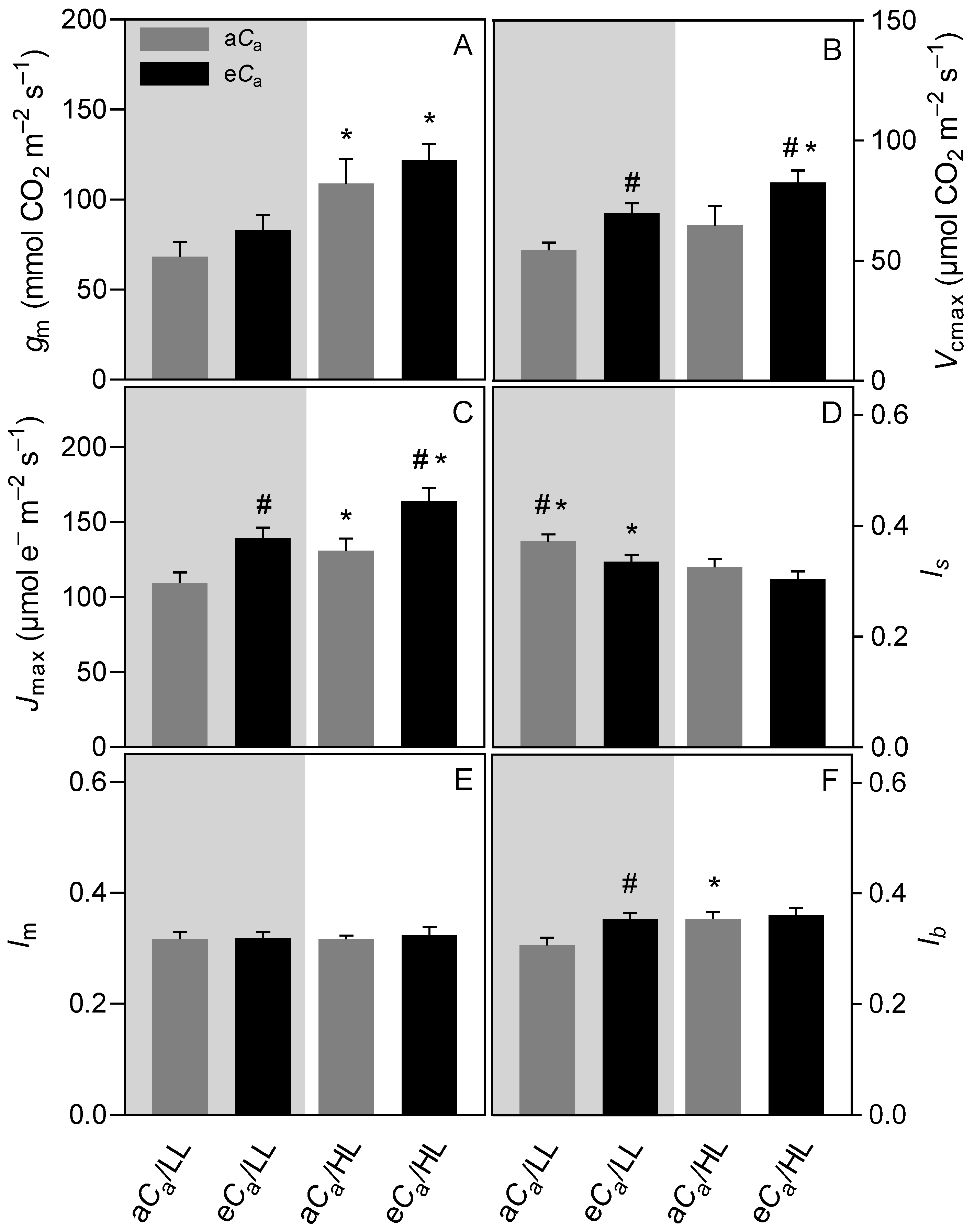
2.2. A/Ci Curves
2.3. Photosynthetic N-Use Efficiency and N Partitioning
2.4. Metabolites
2.5. Morphoanatomical Traits
2.6. Growth Traits and Biomass Partitioning
3. Discussion
3.1. The eCa Increases Stomatal Conductance Only in High-Light-Grown Plants
3.2. The eCa Improves Photosynthetic Performance by Decreasing Diffusional Limitations without Causing Photosynthetic Down-Regulation
3.3. The eCa Improves Photosynthetic-N Use Efficiency Irrespective of Light Levels via Increases in Photosynthetic Performance Rather Than Alterations in N Partitioning
3.4. The eCa Interacts with High Light to Promote Growth, but Changes in Biomass Partitioning Are Only Affected by the CO2 Supply
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Sampling
4.2. Gas Exchanges and Chlorophyll a Fluorescence Analysis
4.3. Nitrogen-Use Efficiency and N Partitioning
4.4. Metabolites
4.5. Morphoanatomical Analyses
4.6. Growth Attributes
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
Appendix A. Supplementary Data

{kind=link}
{kind=link}
{kind=link}
| Parameters | Factors | ||
|---|---|---|---|
| Light | Ca | Light x Ca Interaction | |
| Gas exchange | |||
| A | *** | *** | *** |
| gs | *** | * | *** |
| Ci | ns | *** | ns |
| E | *** | ns | ns |
| Rp/AG | *** | *** | ns |
| Rn | *** | * | ** |
| Gas exchange (light transition) | |||
| A | ns | *** | ** |
| gs | ns | *** | *** |
| ETR | *** | *** | *** |
| ETR/A | *** | *** | *** |
| Chlorophyll a fluorescence | |||
| Fv/Fm | ns | ns | ns |
| Fv′/Fm′ | *** | ns | ns |
| ΦPSII | *** | ns | ns |
| ETR | *** | *** | *** |
| ETR/A | *** | *** | ns |
| NPQ | *** | ** | ns |
| qp | *** | ns | ns |
| A/Ci curves | |||
| gm | *** | ns | ns |
| Vcmax | * | *** | ns |
| Jmax | * | *** | ns |
| ls | * | ** | ns |
| lm | ns | ns | ns |
| lb | * | * | ns |
| Nitrogen, use efficiency and partitioning | |||
| Nm (g kg−1) | ** | *** | ns |
| Na (g m−2) | *** | *** | * |
| PR | *** | ** | ns |
| PB | *** | ns | ns |
| PL | *** | ns | ns |
| PP | *** | * | ns |
| PNP | *** | * | ns |
| PNUE | ** | *** | ns |
| Metabolites | |||
| Chl a + b | *** | *** | ns |
| Chl/N | * | ns | ns |
| Strach | *** | ns | * |
| Glu | *** | ns | * |
| Fru | *** | ns | *** |
| Suc | ns | *** | ns |
| NSC | *** | ns | ns |
| TAA | *** | *** | ns |
| Proteins | ns | *** | ns |
| Morphoanatomical traits | |||
| SD | *** | ns | ns |
| SI | *** | ns | ns |
| SS | ns | ns | ns |
| SPD | ns | ns | ns |
| gwmax | *** | ** | ns |
| Growth attributes | |||
| TDM | *** | *** | *** |
| TLA | *** | *** | *** |
| SLA | *** | ns | ns |
| LMR | ns | *** | * |
| SMR | ns | ns | ns |
| OMR | ** | ns | ns |
| PMR | ** | ** | ns |
| RMR | ns | *** | ns |
| LAR | *** | * | ns |
| Parameters | LL | HL | ||
|---|---|---|---|---|
| aCa | eCa | aCa | eCa | |
| Fv/Fm | 0.80 ± 0.002 | 0.79 ± 0.003 | 0.80 ± 0.004 | 0.80 ± 0.004 |
| Fv ′/Fm′ | 0.62 ± 0.009 | 0.60 ± 0.005 | 0.57 ± 0.008 * | 0.57 ± 0.004 * |
| ΦPSII | 0.46 ± 0.003 | 0.47 ± 0.004 | 0.18 ± 0.006 * | 0.20 ± 0.001 * |
| ETR (µmol e− m−2 s−1) | 41.6 ± 0.3 | 42.6 ± 0.2 | 84.3 ± 2.8 * | 103.3 ± 2.9 #* |
| ETR/A (mol e− mol−1 CO2) | 12.8 ± 0.6 | 6.2 ± 0.2 # | 13.9 ± 0.4 * | 7.9 ± 0.4 #* |
| NPQ | 1.52 ± 0.04 | 1.34 ± 0.04 # | 2.00 ± 0.05 * | 1.79 ± 0.08 #* |
| qp | 0.70 ± 0.004 | 0.69 ± 0.003 | 0.33 ± 0.012 * | 0.37 ± 0.012 #* |
References
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Poorter, H.; Knopf, O.; Wright, I.J.; Temme, A.A.; Hogewoning, S.W.; Graf, A.; Cernusak, L.A.; Pons, T.L. A meta-analysis of responses of C3 plants to atmospheric CO2: Dose–response curves for 85 traits ranging from the molecular to the whole-plant level. New Phytol. 2022, 233, 1560–1596. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Global Change Biol. 2021, 27, 27–49. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.D.; Tomaz, M.A.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C. Combined effects of elevated [CO2] and high temperature on leaf mineral balance in Coffea spp. plants. Clim. Change 2014, 126, 365–379. [Google Scholar] [CrossRef]
- Kitao, M.; Tobita, H.; Kitaoka, S.; Harayama, H.; Yazaki, K.; Komatsu, M.; Koike, T. Light energy partitioning under various environmental stresses combined with elevated CO2 in three deciduous broadleaf tree species in Japan. Climate 2019, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Birami, B.; Nägele, T.; Gattmann, M.; Preisler, Y.; Gast, A.; Arneth, A.; Ruehr, N.K. Hot drought reduces the effects of elevated CO2 on tree water use efficiency and carbon metabolism. New Phytol. 2020, 226, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Bloom, A.J.; Asensio, J.S.R.; Randall, L.; Rachmilevitch, S.; Cousins, A.B.; Carlisle, E.A. CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants. Ecology 2012, 93, 355–367. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M. Ecophysiological constraints on the production of shaded and unshaded coffee: A review. Field Crops Res. 2004, 86, 99–114. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Change 2019, 152, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.C.V.; Galmés, J.; Cavatte, P.C.; Pereira, L.F.; Ventrella, M.C.; DaMatta, F.M. Understanding the low photosynthetic rates of sun and shade coffee leaves: Bridging the gap on the relative roles of hydraulic, diffusive and biochemical constraints to photosynthesis. PLoS ONE 2014, 9, e95571. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Avila, R.T.; Cardoso, A.A.; Martins, S.C.; Ramalho, J.C. Physiological and agronomic performance of the coffee crop in the context of climate change and global warming: A review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- Koutouleas, A.; Sarzynski, T.; Bertrand, B.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Léran, S.; Markussen, B.; et al. Shade effects on yield across different Coffea arabica cultivars—How much is too much? A meta-analysis. Agron. Sustain. Dev. 2022, 42, 55. [Google Scholar] [CrossRef]
- Koutouleas, A.; Sarzynski, T.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Rigal, C.; Vaast, P.; Ramalho, J.C.; et al. Shaded-coffee: A nature-based strategy for coffee production under climate change? A review. Front. Sustain. Food Syst. 2022, 6, 877476. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle Rivera, O.; Kirschke, D. A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Clim. Change 2015, 129, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Moat, J.; Williams, J.; Baena, S.; Wilkinson, T.; Gole, T.W.; Challa, Z.K.; Demissew, S.; Davis, A.P. Resilience potential of the Ethiopian coffee sector under climate change. Nat. Plants 2017, 3, 17081. [Google Scholar] [CrossRef]
- Gomes, L.C.; Bianchi, F.J.J.A.; Cardoso, I.M.; Fernandes, R.B.A.; Fernandes Filho, E.I.; Schulte, R.P.O. Agroforestry systems can mitigate the impacts of climate change on coffee production: A spatially explicit assessment in Brazil. Agric. Ecosyst. Environ. 2020, 294, 106858. [Google Scholar] [CrossRef]
- Martins, M.Q.; Rodrigues, W.P.; Fortunato, A.S.; Leitão, A.E.; Rodrigues, A.P.; Pais, I.P.; Martins, L.D.; Silva, M.J.; Reboredo, F.H.; Partelli, F.L.; et al. Protective response mechanisms to heat stress in interaction with high [CO2] conditions in Coffea spp. Front. Plant Sci. 2016, 7, 947. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; et al. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supraoptimal temperatures in tropical Coffea arabica and C. canephora species. Global Change Biol. 2016, 22, 415–431. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Pais, I.P.; Ribeiro-Barros, A.I.; Martins, L.D.; Tomaz, M.A.; Rodrigues, W.P.; Campostrini, E.; Semedo, J.N.; Fortunato, A.S.; Martins, M.Q.; et al. Lipid profile adjustments may contribute to warming acclimation and to heat impact mitigation by elevated [CO2] in Coffea spp. Environ. Exp. Bot. 2019, 167, 103856. [Google Scholar] [CrossRef]
- Ávila, R.T.; Almeida, W.L.; Costa, L.C.; Machado, K.L.; Barbosa, M.L.; de Souza, R.P.; Martino, P.B.; Juárez, M.A.T.; Marçal, D.M.S.; Martins, S.C.V.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- Ávila, R.T.; Martins, S.C.; Sanglard, L.M.; Dos Santos, M.S.; Menezes-Silva, P.E.; Detman, K.C.; Sanglard, M.L.; Cardoso, A.A.; Morais, L.E.; Vital, C.E.; et al. Starch accumulation does not lead to feedback photosynthetic downregulation in girdled coffee branches under varying source-to-sink ratios. Trees 2020, 34, 1–16. [Google Scholar] [CrossRef]
- Catarino, I.A.C.; Monteiro, G.B.; Ferreira, M.J.; Torres, L.M.B.; Domingues, D.S.; Centeno, D.C.; Lobo, A.K.M.; Silva, E.A. Elevated [CO2] mitigates drought effects and increases leaf 5-o-caffeoylquinic acid and caffeine concentrations during the early growth of Coffea arabica plants. Front. Sustain. Food Syst. 2021, 5, 676207. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Jorge, T.; Osorio, S.; Pott, D.M.; Lidon, F.C.; DaMatta, F.M.; Marques, I.; Ribeiro-Barros, A.I.; Ramalho, J.C.; António, C. Primary metabolite profile changes in Coffea spp. promoted by single and combined exposure to drought and elevated CO2 concentration. Metabolites 2021, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, M.J.; Martins, S.; Semedo, M.C.; et al. Intrinsic non-stomatal resilience to drought of the photosynthetic apparatus in Coffea spp. is strengthened by elevated air [CO2]. Tree Physiol. 2021, 41, 708–727. [Google Scholar] [CrossRef]
- Marques, I.; Rodrigues, A.P.; Gouveia, D.; Lidon, F.C.; Martins, S.; Semedo, M.C.; Gaillard, J.C.; Pais, I.P.; Semedo, J.N.; Scotti-Campos, P.; et al. High-resolution shotgun proteomics reveals that increased air [CO2] amplifies the acclimation response of Coffea species to drought regarding antioxidative, energy, sugar, and lipid dynamics. J. Plant Physiol. 2022, 276, 153788. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, J.C.; Rodrigues, A.P.; Semedo, J.N.; Pais, I.P.; Martins, L.D.; Simões-Costa, M.C.; Leitão, A.E.; Fortunato, A.S.; Batista-Santos, P.; Palos, I.M.; et al. Sustained photosynthetic performance of Coffea spp. under long-term enhanced [CO2]. PLoS ONE 2013, 8, e82712. [Google Scholar] [CrossRef] [Green Version]
- Ghini, R.; Torre-Neto, A.; Dentzien, A.F.M.; Guerreiro-Filho, O.; Iost, R.; Patrício, F.R.A.; Prado, J.S.M.; Thomaziello, R.A.; Bettiol, W.; DaMatta, F.M. Coffee growth, pest and yield responses to free-air CO2 enrichment. Clim. Change 2015, 132, 307–320. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Godoy, A.G.; Menezes-Silva, P.E.; Martins, S.C.V.; Sanglard, L.M.V.P.; Morais, L.E.; Torre-Neto, A.; Ghini, R. Sustained enhancement of photosynthesis in coffee trees grown under free-air CO2 enrichment conditions: Disentangling the contributions of stomatal, mesophyll, and biochemical limitations. J. Exp. Bot. 2016, 67, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Rakočević, M.; Ribeiro, R.V.; Marchiori, P.E.R.; Filizola, H.F.; Batista, E.R. Structural and functional changes in coffee trees after 4 years under free air CO2 enrichment. Ann. Bot. 2018, 21, 1065–1078. [Google Scholar] [CrossRef] [PubMed]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.J.; Iba, K.; Schroeder, J.I. CO2 sensing and CO2 regulation of stomatal conductance: Advances and open questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marçal, D.M.; Avila, R.T.; Quiroga-Rojas, L.F.; de Souza, R.P.; Junior, C.C.G.; Ponte, L.R.; Barbosa, M.L.; Oliveira, L.A.; Martins, S.C.V.; Ramalho, J.D.C.; et al. Elevated [CO2] benefits coffee growth and photosynthetic performance regardless of light availability. Plant Physiol. Biochem. 2021, 158, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Cavatte, P.C.; Oliveira, A.A.G.; Morais, L.E.; Martins, S.C.V.; Sanglard, L.M.V.P.; DaMatta, F.M. Could shading reduce the negative impacts of drought on coffee? A morphophysiological analysis. Physiol. Plant. 2012, 144, 11–22. [Google Scholar] [CrossRef]
- Rodríguez-López, N.F.; Martins, S.C.; Cavatte, P.C.; Silva, P.E.; Morais, L.E.; Pereira, L.F.; Reis, J.V.; Ávila, R.T.; Godoy, A.G.; Lavinski, A.O.; et al. Morphological and physiological acclimations of coffee seedlings to growth over a range of fixed or changing light supplies. Environ. Exp. Bot. 2014, 102, 1–10. [Google Scholar] [CrossRef]
- Franks, P.J.; Farquhar, G.D. The mechanical diversity of stomata and its significance in gas exchange control. Plant Physiol. 2007, 143, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, C.; Batke, S.P.; Yiotis, C.; Caballero, R.; Soh, W.K.; Murray, M.; McElwain, J.C. Increasing stomatal conductance in response to rising atmospheric CO2. Ann. Bot. 2018, 121, 1137–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, S.P.; Taylor, S.H.; Burgess, S.J.; Carmo-Silva, E.; Lawson, T.; Souza, A.P.; Leonelli, L.; Wang, Y. Into the shadows and back into sunlight: Photosynthesis in fluctuating light. Annu. Rev. Plant Biol. 2022, 73, 617–648. [Google Scholar] [CrossRef] [PubMed]
- Matos, F.S.; Wolfgramm, R.; Gonçalves, F.V.; Cavatte, P.C.; Ventrella, M.C.; DaMatta, F.M. Phenotypic plasticity in response to light in the coffee tree. Environ. Exp. Bot. 2009, 67, 421–427. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Clarke, V.C. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Vera, P.; Blanco-Flores, H.; Francisco, A.M.; Castillo, J.; Tezara, W. Silicon dioxide nanofertilizers improve photosynthetic capacity of two Criollo cocoa clones (Theobroma cacao L.). Exp. Agric. 2021, 57, 85–102. [Google Scholar] [CrossRef]
- Lei, Z.Y.; Wang, H.; Wright, I.J.; Zhu, X.G.; Niinemets, Ü.; Li, Z.L.; Sun, D.S.; Dong, N.; Zhang, W.F.; Zhou, Z.L.; et al. Enhanced photosynthetic nitrogen use efficiency and increased nitrogen allocation to photosynthetic machinery under cotton domestication. Photosynth. Res. 2021, 150, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Xiao, L.X.; Zhang, Y.B.; Fahad, S.; Wang, F. dep1 improves rice grain yield and nitrogen use efficiency simultaneously by enhancing nitrogen and dry matter translocation. J. Integr. Agric. 2022, 21, 3185–3198. [Google Scholar] [CrossRef]
- Onoda, Y.; Wright, I.J.; Evans, J.R.; Hikosaka, K.; Kitajima, K.; Niinemets, Ü.; Poorter, H.; Tosens, T.; Westoby, M. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytol. 2017, 214, 1447–1463. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Xu, H.; Dong, S.; Chu, J.; Dai, X.; He, M. Optimized nitrogen allocation favors improvement in canopy photosynthetic nitrogen-use efficiency: Evidence from late-sown winter wheat. Environ. Exp. Bot. 2019, 159, 75–86. [Google Scholar] [CrossRef]
- Kanno, K.; Suzuki, Y.; Makino, A. A small decrease in Rubisco content by individual suppression of RBCS genes leads to improvement of photosynthesis and greater biomass production in rice under conditions of elevated CO2. Plant Cell Physiol. 2017, 58, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef] [Green Version]
- Apgaua, D.M.; Tng, D.Y.; Forbes, S.J.; Ishida, Y.F.; Vogado, N.O.; Cernusak, L.A.; Laurance, S.G. Elevated temperature and CO2 cause differential growth stimulation and drought survival responses in eucalypt species from contrasting habitats. Tree Physiol. 2019, 39, 1806–1820. [Google Scholar] [CrossRef]
- Dawson, W.; Rohr, R.P.; van Kleunen, M.; Fischer, M. Alien plant species with a wider global distribution are better able to capitalize on increased resource availability. New Phytol. 2012, 194, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Lambers, H.; Evans, J.R. Trait correlation networks: A whole-plant perspective on the recently criticized leaf economic spectrum. New Phytol. 2013, 201, 378–382. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 1–39. [Google Scholar]
- Rodeghiero, M.; Niinemets, U.; Cescatti, A. Major diffusion leaks of clampon leaf cuvettes still unaccounted: How erroneous are the estimates of Farquhar et al. model parameters? Plant Cell Environ. 2007, 30, 1006–1022. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, U.; Cescatti, A.; Rodeghiero, M.; Tosens, T. Complex adjustments of photosynthetic potentials and internal diffusion conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex. Plant Cell Environ. 2006, 29, 1159–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef]
- Valentini, R.; Epron, D.; Deangelis, P.; Matteucci, G.; Dreyer, E. In-Situ estimation of net CO2 assimilation, photosynthetic electron flow and photorespiration in Turkey Oak (Q. cerris L.) leaves-diurnal cycles under different levels of water-supply. Plant Cell Environ. 1995, 18, 631–640. [Google Scholar] [CrossRef]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.C.V.; Galmés, J.; Molins, A.; DaMatta, F.M. Improving the estimation of mesophyll conductance to CO2: On the role of electron transport rate correction and respiration. J. Exp. Bot. 2013, 64, 3285–3298. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Bernacchi, C.J.; Portis, A.R.; Nakano, H.; Von Caemmerer, S.; Long, S.P. Temperature response of mesophyll conductance. Implications for the determination of Rubisco enzyme kinetics and for limitations to photosynthesis in vivo. Plant Physiol. 2012, 130, 1992–1998. [Google Scholar] [CrossRef] [Green Version]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Matos, E.S.; Mendonça, E.S. Nitrogênio total, amônio e nitrato. In Matéria Orgânica do Solo: Métodos de Análises; Mendonça, E.S., Matos, E.S., Eds.; Ed. UFV: Viçosa, Brasil, 2017; Volume 221, pp. 61–69. [Google Scholar]
- Niinemets, Ü.; Tenhunen, J.D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant Cell Environ. 1997, 20, 845–866. [Google Scholar] [CrossRef]
- Praxedes, S.C.; DaMatta, F.M.; Loureiro, M.E.; Ferrão, M.A.G.; Cordeiro, A.T. Effects of long-term soil drought on photosynthesis and carbohydrate metabolism in mature robusta coffee (Coffea canephora Pierre var. kouillou) leaves. Environ. Exp. Bot. 2006, 56, 263–273. [Google Scholar] [CrossRef]
- Ronchi, C.P.; DaMatta, F.M.; Batista, K.D.; Moraes, G.A.B.K.; Loureiro, M.E.; Ducatti, C. Growth and photosynthetic down-regulation in Coffea arabica in response to restricted root volume. Funct. Plant Biol. 2006, 33, 1013–1023. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Andrade, M.T.; Oliveira, L.A.; Pereira, T.S.; Cardoso, A.A.; Batista-Silva, W.; DaMatta, F.M.; Zsögön, A.; Martins, S.C.V. Impaired auxin signaling increases vein and stomatal density but reduces hydraulic efficiency and ultimately net photosynthesis. J. Exp. Bot. 2022, 558, 531–539. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, D.F. SISVAR: Um sistema computacional de análise estatística. Cienc. Agrotecnol. 2011, 35, 1039–1042. [Google Scholar] [CrossRef] [Green Version]



| Parameters | LL | HL | ||
|---|---|---|---|---|
| aCa | eCa | aCa | eCa | |
| Nm (g kg−1) | 30.6 ± 0.01 | 32.8 ± 0.1 | 32.6 ± 0.1 | 37.9 ± 0.1 #* |
| Na (g m−2) | 1.56 ± 0.04 | 1.65 ± 0.08 | 1.73 ± 0.08 | 2. 16 ± 0.08 #* |
| PR | 0.26 ± 0.00 | 0.29 ± 0.02 # | 0.22 ± 0.01 * | 0.23 ± 0.01 * |
| PB | 0.036 ± 0.003 | 0.037 ± 0.002 | 0.026 ± 0.001 * | 0.032 ± 0.001 #* |
| PL | 0.18 ± 0.00 | 0.19 ± 0.01 | 0.16 ± 0.01 | 0.15 ± 0.01 * |
| PP | 0.47 ± 0.01 | 0.52 ± 0.02 # | 0.41 ± 0.02 * | 0.42 ± 0.02 * |
| PNP | 0.52 ± 0.01 | 0.48 ± 0.01 # | 0.59 ± 0.02 * | 0.58 ± 0.02 * |
| PNUE (µmol CO2 g−1 N) | 2.85 ± 0.12 | 6.08 ± 0.27 # | 3.81 ± 0.15 * | 6.41 ± 0.34 # |
| Parameters | LL | HL | ||
|---|---|---|---|---|
| aCa | eCa | aCa | eCa | |
| Chl a + b (mg g−1) | 7.8 ± 0.3 | 6.9 ± 0.3 | 6.4 ± 0.9 * | 4.5 ± 0.2 #* |
| Chl/N (mmol mol−1) | 2.5 ± 0.1 | 2.1 ± 0.1 # | 2.0 ± 0.2 | 1.2 ± 0.1 #* |
| Strach (µmol eq. glucose g−1) | 568 ± 9 | 571 ± 10 | 684 ± 28 * | 641 ± 10 #* |
| Glucose (µmol g−1) | 24.9 ± 1.2 | 23.9 ± 0.6 | 28.8 ± 3.5 * | 34.7 ± 1.5 #* |
| Fructose (µmol g−1) | 29.6 ± 0.6 | 26.3 ± 0.6 # | 33.5 ± 1.6 * | 35.9 ± 0.5 #* |
| Sucrose (µmol g−1) | 64.9 ± 1.9 | 75.3 ± 2.1 # | 63.3 ± 1.9 | 79.5 ± 1.8 # |
| NSC | 687 ± 11 | 696 ± 12 | 810 ± 11 * | 791 ± 9 * |
| TAA (µmol g−1) | 11.3 ± 1.5 | 20.3 ± 0.9 # | 18.7 ± 3.29 * | 29.6 ± 1.2 #* |
| Proteins (mg g−1) | 95 ± 5 | 114 ± 4 # | 100 ± 12 | 121 ± 4 # |
| Parameters | LL | HL | ||
|---|---|---|---|---|
| aCa | eCa | aCa | eCa | |
| SD (mm−2) | 83 ± 6 | 87 ± 5 | 167 ± 10 * | 186 ± 6 * |
| SI (%) | 15.5 ± 0.4 | 16.0 ± 0.3 | 18.8 ± 0.5 * | 20.1 ± 0.7 * |
| SS (µm2) | 172 ± 11 | 164 ± 12 | 161 ± 13 | 193 ± 9 |
| gwmax (mol m−2 s−1) | 0.68 ± 0.05 | 0.70 ± 0.05 | 1.32 ± 0.08 * | 1.61 ± 0.02 #* |
| Parameters | LL | HL | ||
|---|---|---|---|---|
| aCa | eCa | aCa | eCa | |
| TB (g) | 25.3 ± 1.1 | 38.7 ± 1.3 # | 119 ± 7.7 * | 240 ± 7.4 #* |
| Height (cm) | 44 ± 2.1 | 55 ± 1.0 # | 84 ± 3.6 * | 103 ± 0.7 #* |
| TLA (m2) | 0.26 ± 0.01 | 0.37 ± 0.01 # | 0.78 ± 0.05 * | 1.26 ± 0.04 #* |
| LMR | 0.61 ± 0.02 | 0.55 ± 0.01 # | 0.65 ± 0.02 | 0.52 ± 0.02 # |
| SMR | 0.27 ± 0.02 | 0.27 ± 0.01 | 0.24 ± 0.01 | 0.26 ± 0.01 |
| OMR | 0.21 ± 0.01 | 0.21 ± 0.01 | 0.17 ± 0.01 * | 0.18 ± 0.01 |
| PMR | 0.056 ± 0.004 | 0.068 ± 0.005 # | 0.070 ± 0.039 * | 0.082 ± 0.003 #* |
| RMR | 0.12 ± 0.01 | 0.18 ± 0.02 # | 0.11 ± 0.01 | 0.22 ± 0.02 # |
| LAR (m2 kg−1) | 10.5 ± 0.8 | 9.5 ± 0.4 | 6.4 ± 0.2 * | 5.3 ± 0.3 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, A.H.; de Oliveira, U.S.; Oliveira, L.A.; de Carvalho, P.H.N.; de Andrade, M.T.; Pereira, T.S.; Gomes Junior, C.C.; Cardoso, A.A.; Ramalho, J.D.C.; Martins, S.C.V.; et al. Growth and Leaf Gas Exchange Upregulation by Elevated [CO2] Is Light Dependent in Coffee Plants. Plants 2023, 12, 1479. https://doi.org/10.3390/plants12071479
de Souza AH, de Oliveira US, Oliveira LA, de Carvalho PHN, de Andrade MT, Pereira TS, Gomes Junior CC, Cardoso AA, Ramalho JDC, Martins SCV, et al. Growth and Leaf Gas Exchange Upregulation by Elevated [CO2] Is Light Dependent in Coffee Plants. Plants. 2023; 12(7):1479. https://doi.org/10.3390/plants12071479
Chicago/Turabian Stylede Souza, Antonio H., Ueliton S. de Oliveira, Leonardo A. Oliveira, Pablo H. N. de Carvalho, Moab T. de Andrade, Talitha S. Pereira, Carlos C. Gomes Junior, Amanda A. Cardoso, José D. C. Ramalho, Samuel C. V. Martins, and et al. 2023. "Growth and Leaf Gas Exchange Upregulation by Elevated [CO2] Is Light Dependent in Coffee Plants" Plants 12, no. 7: 1479. https://doi.org/10.3390/plants12071479