This study advances our understanding of endozoochory of seeds by non-frugivorous birds, providing information on parameters influencing effective dispersal, such as seed survival, retention time and germinability. It is the first experimental study of gulls (Laridae) to address their important role as vectors of dry-fruited plants [
25]. Like other waterbirds such as ducks, geese and storks, gulls are important vectors of weeds in rice fields and other agricultural habitats [
13,
26]. Our research contributes to the understanding of mechanisms of dispersal of weeds and invasive plants, most of which have been assigned to abiotic dispersal syndromes and have been assumed to rely on humans for long-distance dispersal [
17,
27].
3.1. Retrieval, Viability and Retention Time of Seeds in Relation to Plant Traits
We found that the five weed species (four dry fruited and one fleshy fruited) had high rates of seed survival after gut passage, exceeding 72%. Similar experiments with mallards
Anas platyrhynchos and other Anatidae have found some plant species to have much lower rates of seed survival [
16,
18]. We may even have underestimated seed survival since our experiment ended after 33 h, and some seeds have been recovered after longer periods from Anatidae [
19]. Species with the highest seed survival were dry fruited and of intermediate size (
P. monspeliensis and
C. difformis). In experiments with Anatidae, smaller seeds generally showed higher survival because they escape from the gizzard into the intestines more quickly, although other characteristics such as hardness, seed shape and phylogeny were also important predictors of seed survival [
16], see also [
28,
29].
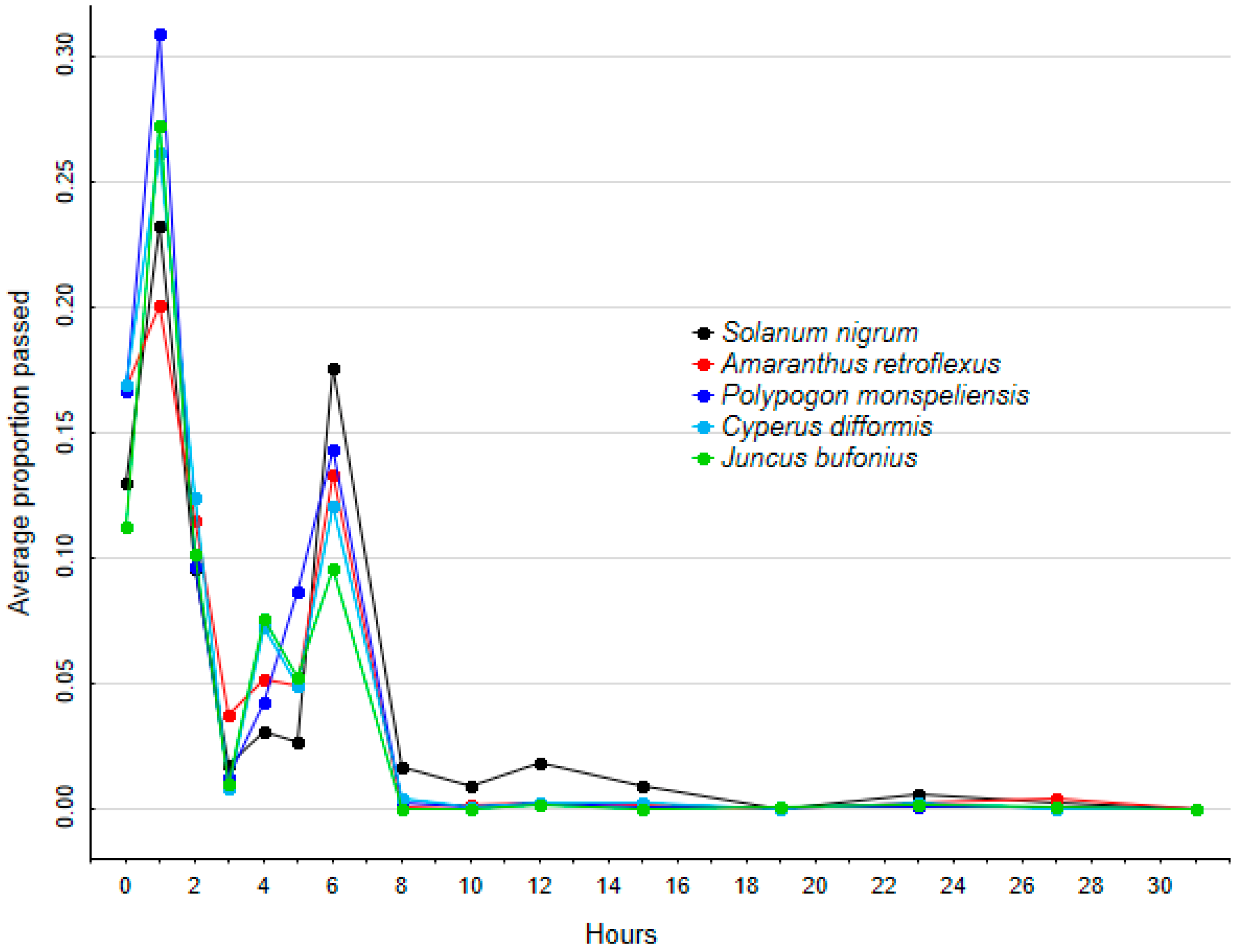
Longer retention times may benefit plants by increasing seed dispersal distances, making it more likely that they can reach and colonize previously unoccupied habitat patches. Given the generally high rates of seed survival, our finding that average retention times were strongly related to seed size was to be expected, with larger seeds taking longer to pass into the intestines and then be egested. In Anatidae experiments where species with larger seeds were found to have shorter retention times, this is likely because these species had low seed survival, and the only seeds to be egested were the fraction that passed quickly into the intestines [
15,
18]. We compared seeds from fleshy-fruited and dry-fruited plants fed simultaneously to birds, and found that seed size, rather than fruit fleshiness, is the main determinant of retention time. However, when comparing separate experiments, the retention times of seeds are generally much lower for frugivorous than for non-frugivorous birds, as a consequence of the effects of fruit pulp on gut activity [
30]. This suggests that non-frugivorous vectors have a greater potential for long-distance seed dispersal than frugivorous ones.
If we had not removed the pulp from
S. nigrum seeds prior to our experiments, we may have found different results, with a change in how this species compared with dry-fruited ones. However, our interest was in comparing how the seeds per se compared in their response to gut passage. We found no evidence that, apart from being the largest of the five species, there was anything special about
S. nigrum seeds and their gut passage in comparison with the other species assigned to barochory or epizoochory syndromes. This is consistent with Costea et al. [
31], who found that there is a diversity of seed architectures amongst angiosperms that allow different species to survive gut passage, and that there is no evidence for a fundamental difference between seeds from fleshy- or dry-fruited species. When combined with the growing body of field data showing how endozoochory is a frequent and often predictable dispersal mechanism for many species assigned to other dispersal syndromes [
9], our results provide further evidence that seeds of dry-fruited species can have adaptations (or exaptations) for non-classical endozoochory. Species such as
A. retroflexus and
J. bufonius have been repeatedly shown to disperse by endozoochory in field studies of gulls, Anatidae or other waterbirds [
12,
32].
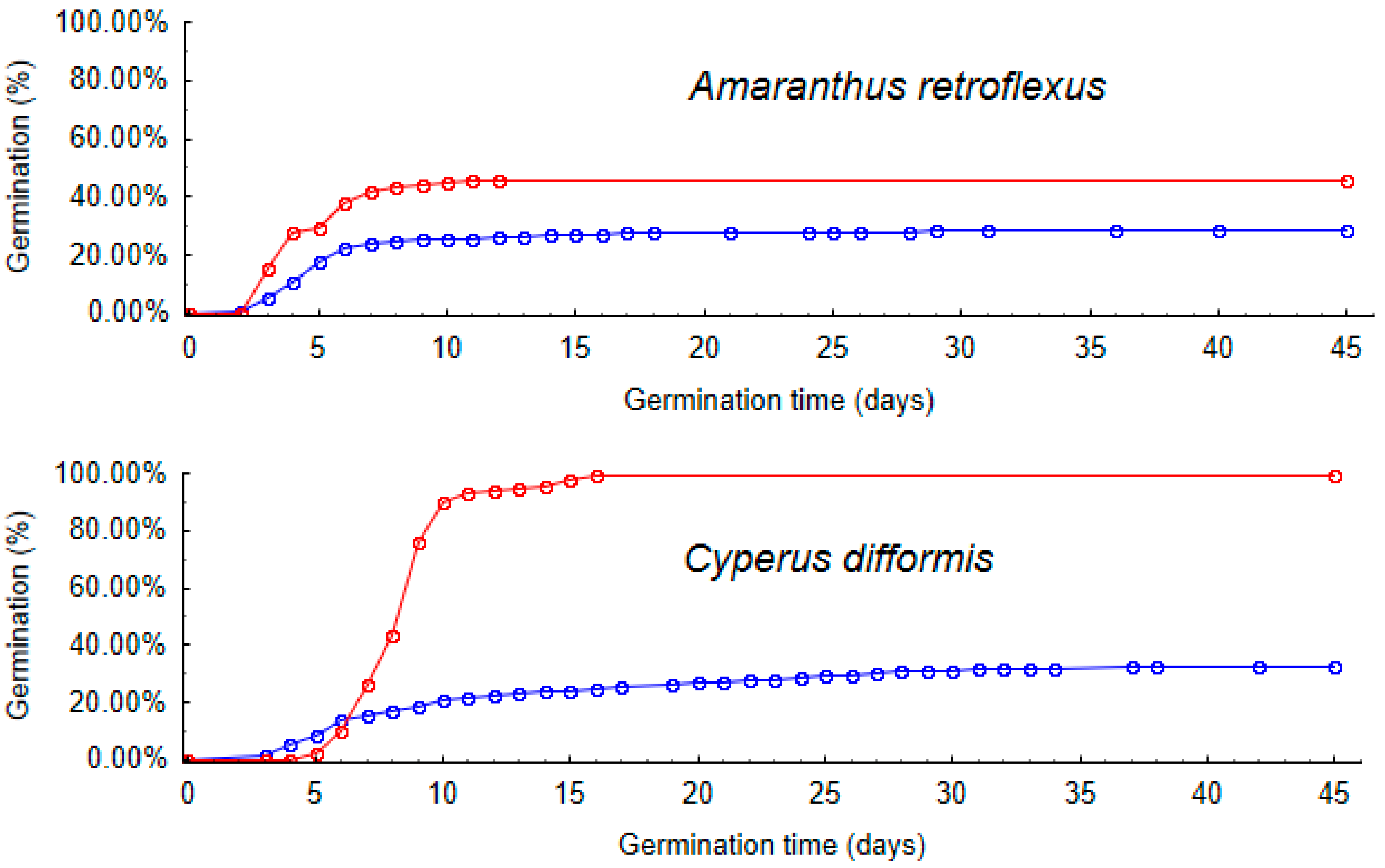
Germination experiments confirmed the viability of the egested seeds, although we found germinability to be higher in control seeds in all five species. This result may have changed if the seeds had not been kept at cold temperatures between collection and our experiments, since this stratification may have broken dormancy in control seeds. If the seeds had been kept at ambient temperature, gut passage may have increased germinability compared to control seeds, as observed in some previous studies [
19,
33]. This is supported by a separate finding that storage time between faeces egestion and seed extraction was correlated with germinability (
Figure S1), although this result was confounded with retention time (see Methods). With hindsight, we should have processed faecal samples in a random order with respect to retention time.
We did not expect to find the observed positive correlation between seed size and the germinability of egested seeds relative to control seeds, especially as larger seeds had longer average retention times, which would be expected to translate into reduced viability [
19,
33]. The impact of gut passage on germination time was unrelated to seed size, since the smallest and largest seeds (of
J. bufonius and
S. nigrum) were the only two for which seeds germinated faster compared to the controls.
J. bufonius is easily the most abundant seed in LBBG faeces and pellets from rice fields of the Guadalquivir estuary [
12,
24], and its rapid germination after gut passage may promote endozoochory given intense intraspecific competition for this weed, which is highly abundant in rice fields, especially between harvest and sowing. It has smaller seeds than the other weeds we studied, but this allows production of more seeds than other species [
34,
35], with 34,000 seeds/ramet [
36].
The relationship between seed size and germinability or germination time is clearly complex and confounded by other seed traits such as dormancy strategies. Kleyheeg et al. 2018 [
37] tested 30 wetland plant species and found that germinability increased with seed size for control seeds, but this relationship was less pronounced after artificial digestion. They did not find a significant influence of seed size on time to germination. In contrast, we found that size was more related to germinability
after gut passage than
before (
Figure 3), although this difference was not significant. Some experimental studies have found a negative correlation between seed size and germinability within plant species [
29].
Our experimental estimations of seed survival and retention time might differ from real values in natural conditions. Avian diet can have an important influence on these parameters [
38], although our birds were fed on an animal diet, which reflects their preference for crayfish in the field. Captivity can change seed survival and retention time due to the limited mobility of the birds compared with those active in the field. Kleyheeg, et al. [
39] found that seed survival increased significantly with forced activity in ducks. LBBGs spend long periods of the day inactive, roosting in rice fields or neighbouring habitats [
40], so our experiment may reflect field conditions quite well.
Martín-Vélez et al. [
13] developed a spatial seed dispersal model based on GPS data for LBBGs and for field and experimental data for four weed species used in our study (
J. bufonius,
C. difformis,
P. monspeliensis and
A. retroflexus). These authors estimated median endozoochory dispersal distances of 690 to 940 m, with the maximum exceeding 150 km. Their model found that seeds would be deposited in suitable habitats for weed establishment such as other rice fields, natural wetlands and other irrigated crops. These geographic distance estimations are longer than those calculated for abiotic dispersal mechanisms such as wind [
41]. For example, the maximum dispersal distance estimated for
J. bufonius via wind was 100 m [
42].
3.2. Characteristics of LBBG That Make Them Good Seed Dispersal Vectors
The LBBG is a long-distance migrant with a broad flyway, moving from North Europe to northwest Africa [
43,
44]. It typically combines migration with foraging during frequent and long-migratory stopovers, the so-called fly-and-forage migration strategy [
44,
45,
46]. Although it does not achieve long daily distances compared with other migratory birds [average instantaneous speed of 38.6 km/h; 44 km/day in autumn and 98 km/day in spring; 44] (average instantaneous speed of 38.6 km/h; 44 km/day in autumn and 98 km/day in spring), the LBBG is still faster and can achieve longer daily distances than other fly-and-forage migrants such as raptors [
45]. On the other hand, LBBG have the ability to find suitable habitats for feeding nearly everywhere along their migratory route due to their opportunistic feeding in inland as well as marine habitats, increasing the possibility that seeds are egested in favourable habitats [
22]. The LBBG may thus be a good vector for long-distance dispersal of weeds, aliens and other plants in a step-by-step manner. Throughout the winter, the LBBG performs daily movements between feeding and roosting sites of up to 80 km [
22] favouring dispersal of seeds from highly anthropized habitats to more natural environments. Due to their common use of landfills, agricultural fields and other human-transformed habitats, the LBBG may be a particularly suitable vector for weeds and invasive species (this study; [
22]).
The LBBG is an opportunistic feeder that includes many different food items in its diet [
47,
48]. This omnivorous diet may enable higher seed survival compared to granivorous bird species. Moreover, the structure of the gizzard, responsible for mechanical digestion, plays a key role in seed survival [
28]. In the case of ducks specialized in the digestion of seeds, the gizzard is very thick and muscular, with the presence of grit which improves mechanical digestion of seeds, although grit may also increase germination rate through scarification [
49]. Omnivorous birds such as LBBGs have relatively small gizzards compared to herbivorous or granivorous birds, and are thus more likely to allow seeds to survive gut passage [
37]. This is likely to explain the particularly high recovery rate of intact seeds in our experiment. Overall, gulls can be expected to provide effective seed dispersal with relatively high-quantity and quality components [
50], although further research into the survivorship of dispersed seeds and resulting seedlings is required.
3.3. Ecological and Management Implications of Weed Dispersal by LBBG
All five species we studied are noxious agricultural weeds in Europe and other parts of the world, causing important losses to food production globally [
51]. Some of them are also invasive aliens and outcompete native flora.
P. monspeliensis,
A. retroflexus and
C. difformis in particular have evolved under the use of intense herbicide treatments, developing herbicide resistance that results in significant economic costs [
52,
53]. In India,
P. monspeliensis is one of the main weeds of wheat and rice fields [
54], and is spreading rapidly in Spain [
55]. It has invaded saltmarshes in Southern California, outcompeting native
Salicornia virginica [
56] and leading to important restoration efforts [
57].
Amaranthus species are among the most serious agricultural weeds in the world [
58,
59]. Their life history traits and allelopathic chemicals allow them to outcompete other plants and significantly reduce grain production and quality. Both
A. retroflexus and
S. nigrum are well known for their allelopathic effects [
60,
61] which significantly reduce germination and growth of agricultural crops by toxic chemical production. Some weed species from our study have also been associated with livestock poisoning, e.g., via bacterial infection, such as
Clavibacter toxicus of seedheads in
P. monspeliensis, such as
Clavibacter toxicus of seedheads in
P. monspeliensis [
62] or chemical compounds in
A. retroflexus [
63,
64].
Solanum nigrum, native to Eurasia and invasive in Australia, is a spill over host of Tobamoviruses, which are amongst the most damaging viruses to horticultural crops [
65].
C. difformis is one of the world’s worst weed species [
66], particularly in rice-growing regions of Europe, US and Asia [
67]. In the Guadalquivir estuary, the main rice cultivation area in Spain, it is considered the worst weed, with resistance to the most commonly used herbicides. On the other hand, the globally distributed
J. bufonius is considered invasive in the US and has been recently recorded in Antarctica, where it may spread rapidly due to climate warming [
68]. Typically, the potential introduction vectors reported by Cuba-Díaz et al. [
68] in Antarctica were humans (tourists, scientists, etc.), with no mention of birds. Our results suggest that gulls and other migratory birds using Antarctica may have a key role transporting propagules of new species that may thrive under new, warmer conditions.
Most studies of weed science during the last two decades have focused on herbicide research. However, the 21st century requires a holistic approach to weed science, with more contributions to weed biology, ecology and genetics [
59,
69]. Understanding the spread of weeds by wildlife is particularly important. Knowledge about dispersal of weeds by birds may be an important contribution to the challenge of achieving weed control through a multidisciplinary perspective. For example, identifying the season when avian vectors are more abundant may facilitate effective integrated weed management and identify optimal timing for weed control. Ricefields in Spain and other parts of the Mediterranean region are major wintering sites for many species of waterbirds dispersing weeds, such as gulls, storks, geese, and other waterbirds [
20]. Proactive management activities in the winter could contribute to an integrated approach to weed management in rice fields. On the other hand, waterbirds reduce the density of weed seeds in rice fields through their foraging [
20].
Integrating genetic and genomic studies of weeds transported by birds and their herbicide resistance would be another important future field of research. Even a single herbicide-resistant weed seed may become enough to promote the colonization of a whole field [
70]. Globally, the role of waterbirds in the dispersal dynamics of weeds has been ignored to date, and recognizing it may help to predict their spread and to develop suitable management strategies for control.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}