
Genome Size Variation across a Cypriot Fabeae Tribe Germplasm Collection
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
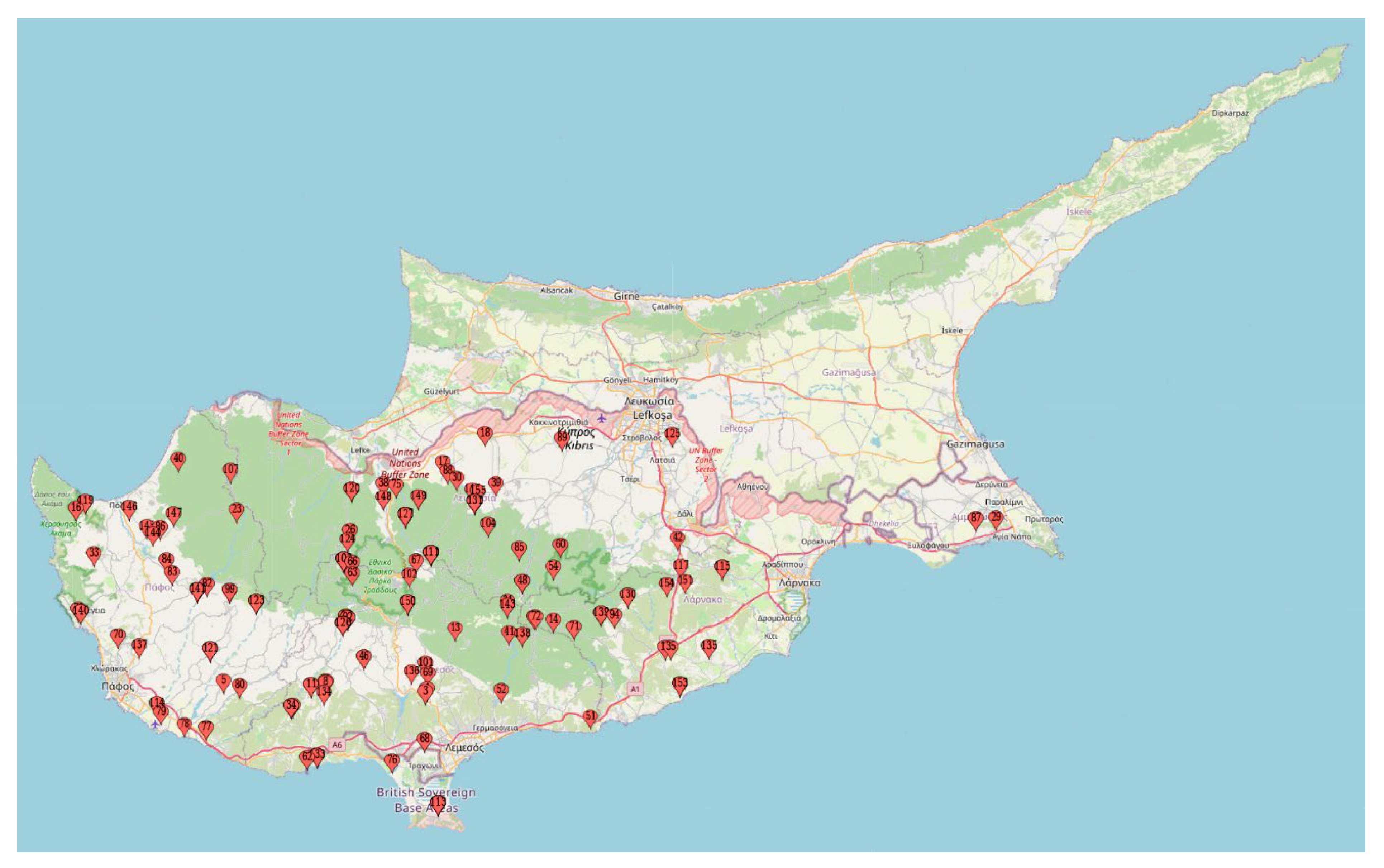
2.1. Plant Material
2.2. Sample Preparation
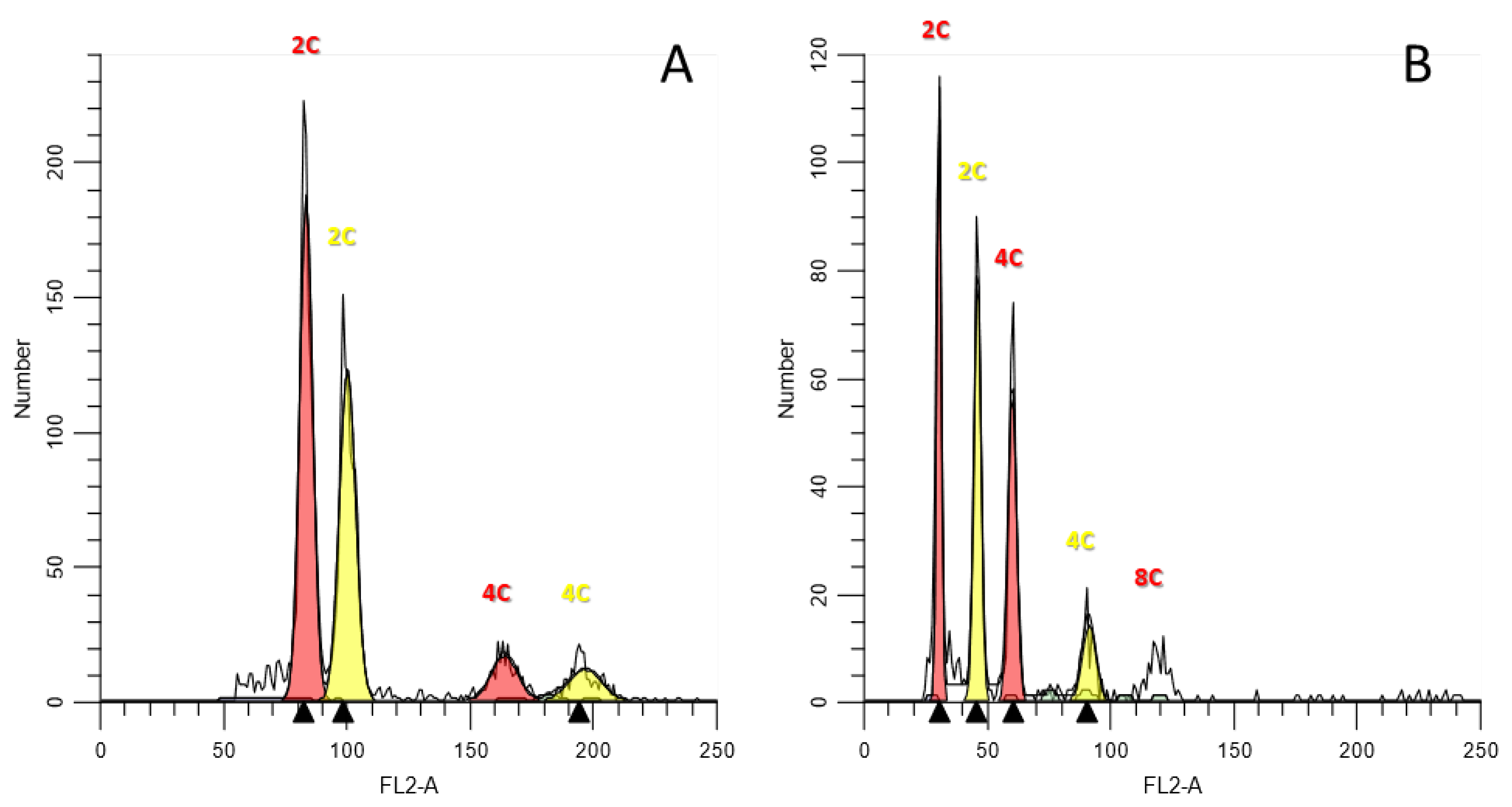
2.3. Flow Cytometry
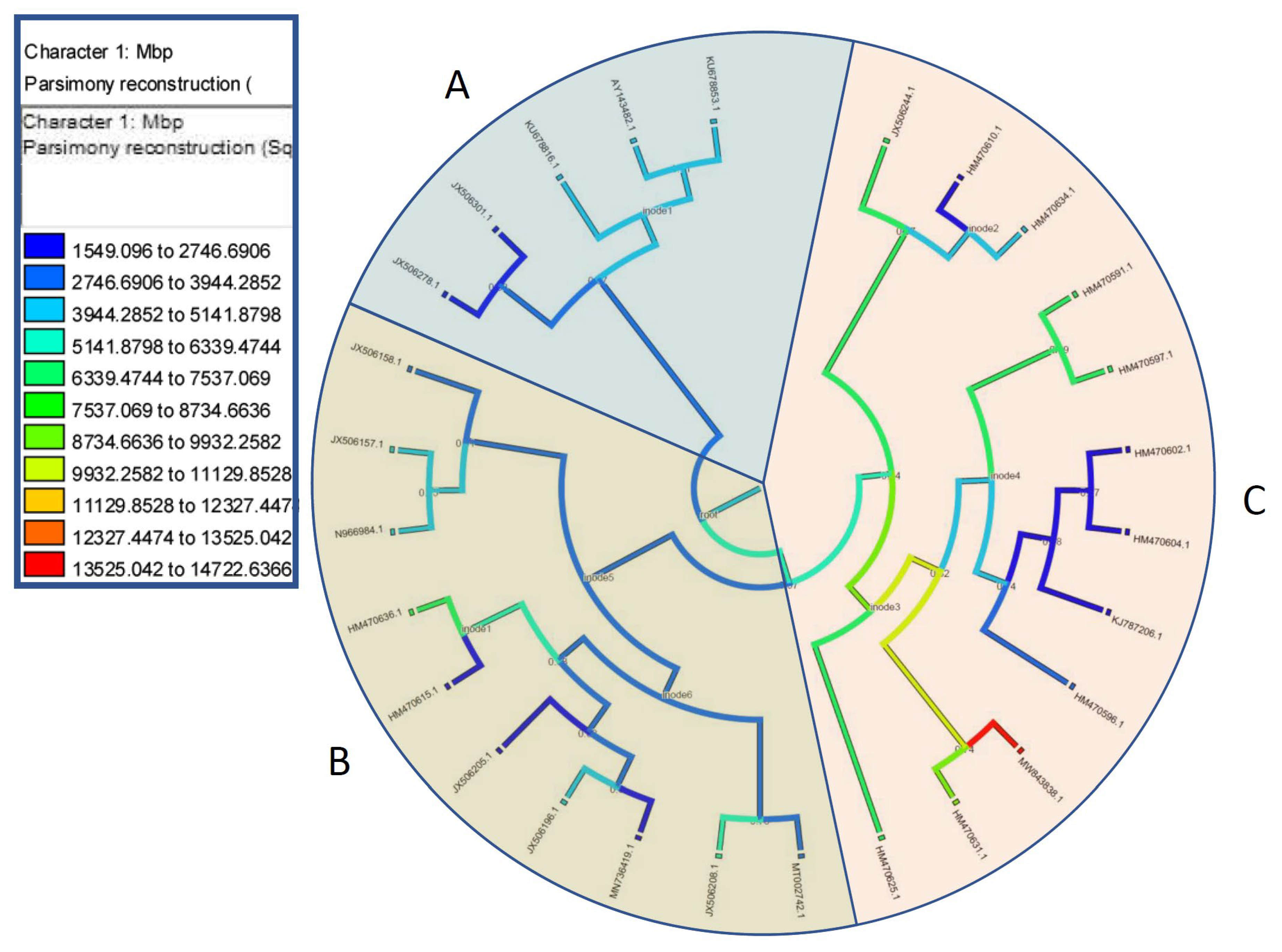
2.4. C-Value Character Evolution
2.5. Statistical Analyses
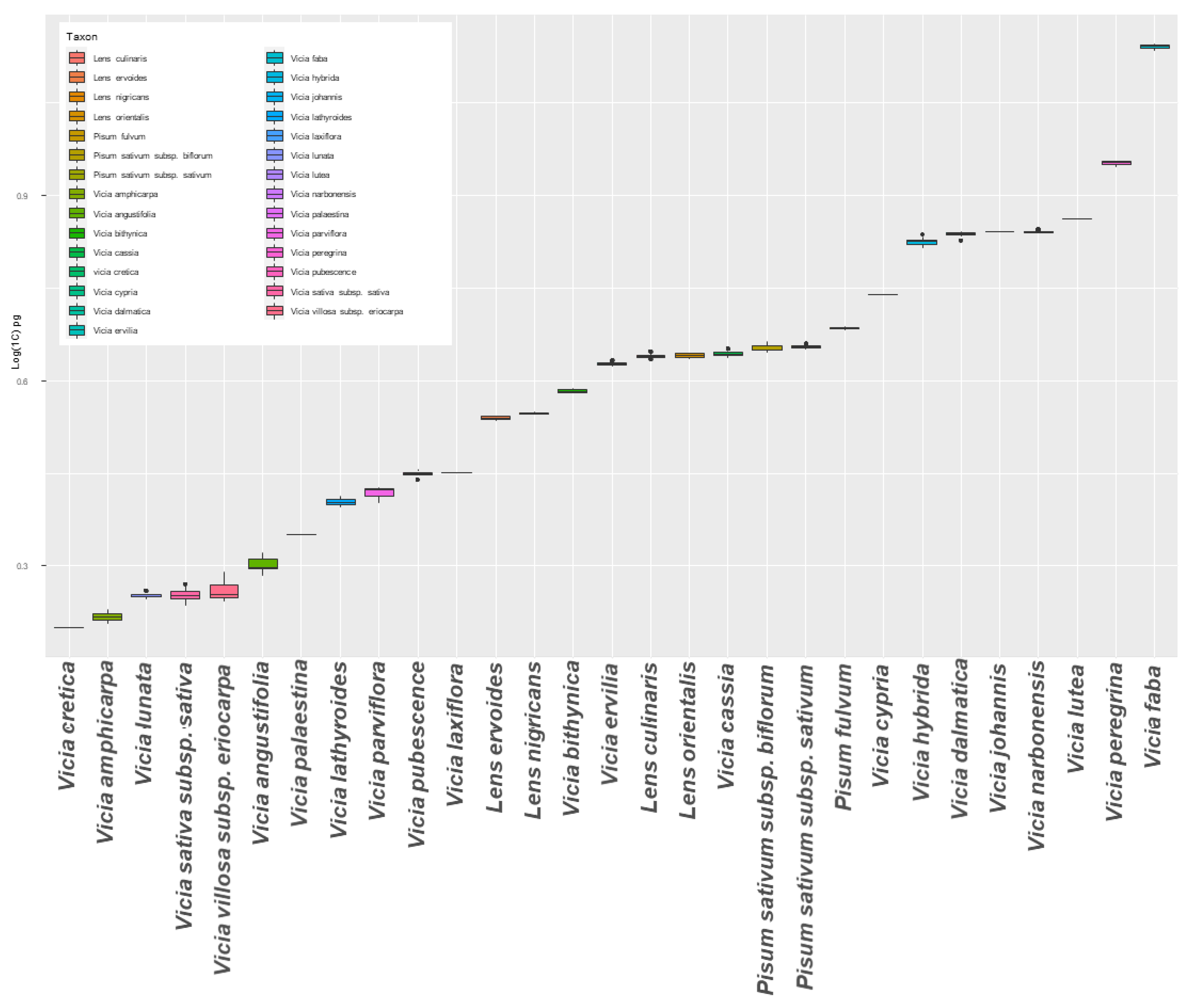
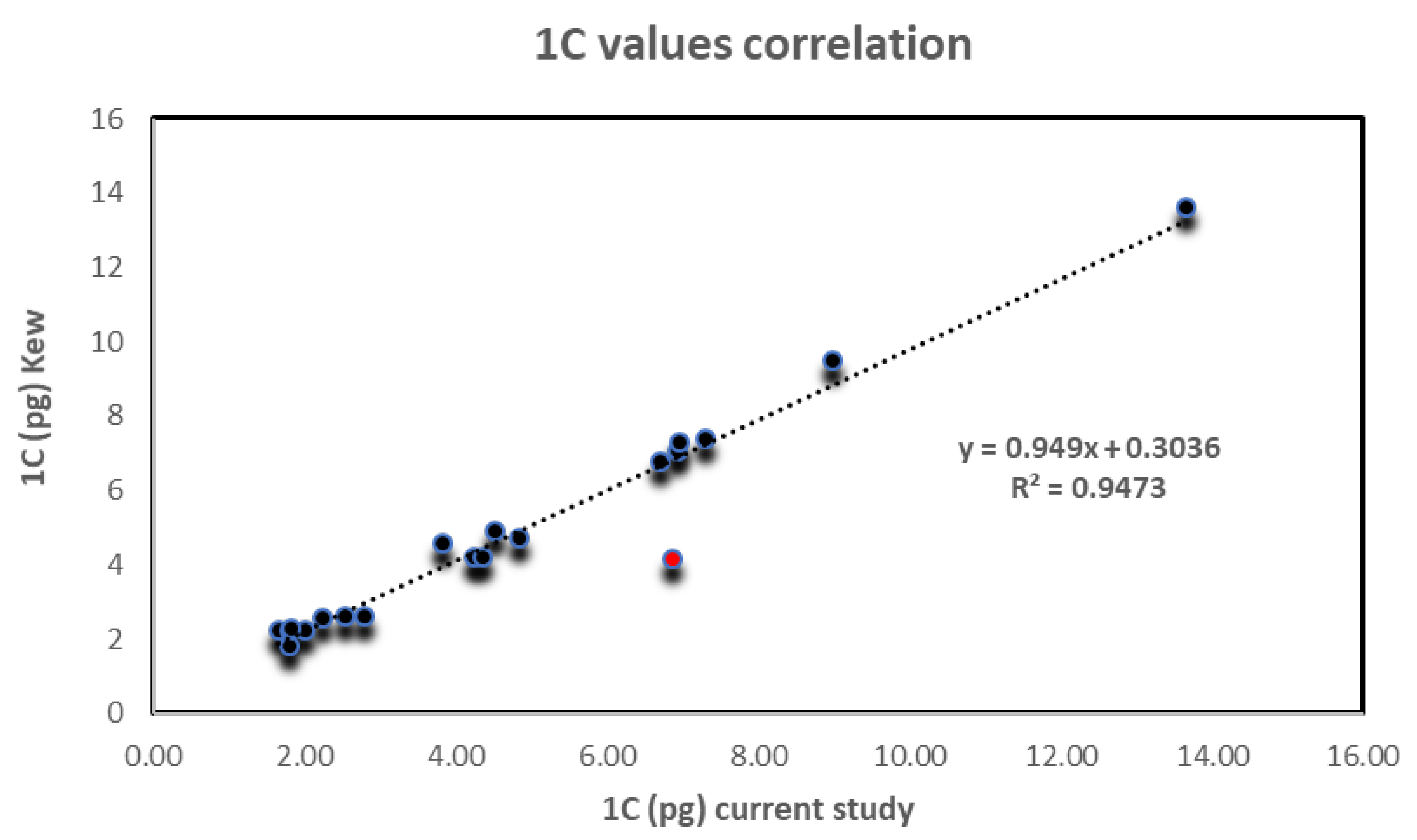
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Thomas, C.A., Jr. The genetic organization of chromosomes. Annu. Rev. Genet. 1971, 5, 237–256. [Google Scholar] [CrossRef]
- Klein, S.P.; Anderson, S.N. The evolution and function of transposons in epigenetic regulation in response to the environment. Curr. Opin. Plant Biol. 2022, 69, 102277. [Google Scholar] [CrossRef] [PubMed]
- Macas, J.; Novak, P.; Pellicer, J.; Cizkova, J.; Koblizkova, A.; Neumann, P.; Fukova, I.; Dolezel, J.; Kelly, L.J.; Leitch, I.J. In depth characterization of repetitive DNA in 23 plant genomes reveals sources of genome size variation in the legume tribe fabeae. PLoS ONE 2015, 10, e0143424. [Google Scholar] [CrossRef]
- Fleischmann, A.; Michael, T.P.; Rivadavia, F.; Sousa, A.; Wang, W.; Temsch, E.M.; Greilhuber, J.; Müller, K.F.; Heubl, G. Evolution of genome size and chromosome number in the carnivorous plant genus Genlisea (Lentibulariaceae), with a new estimate of the minimum genome size in angiosperms. Ann. Bot. 2014, 114, 1651–1663. [Google Scholar] [CrossRef] [Green Version]
- Pellicer, J.; Fay, M.F.; Leitch, I.J. The largest eukaryotic genome of them all? Bot. J. Linn. Soc. 2010, 164, 10–15. [Google Scholar]
- Talukdar, D. Leguminosae. Brenner’s Encycl. Genet. Second Ed. 2013, 2, 212–216. [Google Scholar]
- Vižintin, L.; Javornik, B.; Bohanec, B. Genetic characterization of selected Trifolium species as revealed by nuclear DNA content and ITS rDNA region analysis. Plant Sci. 2006, 170, 859–866. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J.B.; Heslop-Harrison, J.S.; Riley, R. Nuclear DNA amounts in angiosperms. Proc. R. Soc. London. Ser. B Biol. Sci. 1997, 216, 179–199. [Google Scholar]
- Tacke, R.; Ecke, W.; Höfer, M.; Sass, O.; Link, W. Fine-mapping of the major locus for vicine and convicine in faba bean (Vicia faba) and marker-assisted breeding of a novel, low vicine and convicine winter faba bean population. Plant Breed. 2022, 141, 644–657. [Google Scholar]
- Aguilar-Benitez, D.; Casimiro-Soriguer, I.; Maalouf, F.; Torres, A.M. Linkage mapping and QTL analysis of flowering time in faba bean. Sci. Rep. 2021, 11, 13716. [Google Scholar]
- Carrillo-Perdomo, E.; Vidal, A.; Kreplak, J.; Duborjal, H.; Leveugle, M.; Duarte, J.; Desmetz, C.; Deulvot, C.; Raffiot, B.; Marget, P.; et al. Development of new genetic resources for faba bean (Vicia faba L.) breeding through the discovery of gene-based SNP markers and the construction of a high-density consensus map. Sci. Rep. 2020, 10, 6790. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Jiang, J.; Zhang, H.; Liu, R.; Strelkov, S.; Hwang, S.-F.; Chang, K.-F.; Yang, F.; Miao, Y.; He, Y.; et al. Density enhancement of a faba bean genetic linkage map (Vicia faba) based on simple sequence repeats markers. Plant Breed. 2019, 138, 207–215. [Google Scholar] [CrossRef]
- Ma, L.; Wang, X.; Yan, M.; Liu, F.; Zhang, S.; Wang, X. Genome survey sequencing of common vetch (Vicia sativa L.) and genetic diversity analysis of Chinese germplasm with genomic SSR markers. Mol. Biol. Rep. 2022, 49, 313–320. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, Q.; Yan, B.; Zhang, J.; Xue, C.; Chen, J.; Lin, Y.; Zhang, X.; Shen, W.; Chen, X. Single-Molecule Real-Time and Illumina-Based RNA Sequencing Data Identified Vernalization-Responsive Candidate Genes in Faba Bean (Vicia faba L.). Front. Genet. 2021, 12, 656137. [Google Scholar] [CrossRef] [PubMed]
- Martos-Fuentes, M.; Egea-Gilabert, C.; Mezaka, I.; Fernández, J.A.; Egea-Cortines, M.; Weiss, J. Distance analysis among northern and southern European legume accessions using next-generation sequencing reveal discrepancies between geographic and genetic origins. Sci. Hortic. 2019, 243, 498–505. [Google Scholar]
- Varshney, R.K.; Glaszmann, J.-C.; Leung, H.; Ribaut, J.-M. More genomic resources for less-studied crops. Trends Biotechnol. 2010, 28, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.T. Preface: The Evolution of the Evolution of the Genome. In The Evolution of the Genome; Academic Press: Cambridge, MA, USA, 2005; pp. vii–xi. [Google Scholar]
- Pellicer, J.; Leitch, I.J. The Plant DNA C-values database (release 7.1): An updated online repository of plant genome size data for comparative studies. New Phytol. 2020, 226, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, H.; Cameron, F.M.; Hazarika, M.H.; Jones, G.H. Nuclear Variation between Diploid Angiosperms. Nature 1966, 211, 828–830. [Google Scholar] [CrossRef] [PubMed]
- Soldano, A. Riproposizione di taxa sottospecifici prioritari dovuti a botanici italiani. Natura Bresciana 1992, 27, 51–56. [Google Scholar]
- Maxted, N.; Ambrose, M. Peas (Pisum L.). In Plant Genetic Resources of Legumes in the Mediterranean. Current Plant Science and Biotechnology in Agriculture; Springer: Dordrecht, The Netherlands, 2001; Volume 39, pp. 181–190. [Google Scholar]
- Wong, M.M.L.; Gujaria-Verma, N.; Ramsay, L.; Yuan, H.Y.; Caron, C.; Diapari, M.; Vandenberg, A.; Bett, K.E. Classification and Characterization of Species within the Genus Lens Using Genotyping-by-Sequencing (GBS). PLoS ONE 2015, 10, e0122025. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.-F.; Gao, Q.; Liu, F.; Wang, Z.; Wang, J.-L.; Wang, X.-G. DNA barcoding evaluation of Vicia (Fabaceae): Comparative efficacy of six universal barcode loci on abundant species. J. Syst. Evol. 2020, 58, 77–88. [Google Scholar] [CrossRef]
- Schaefer, H.; Hechenleitner, P.; Santos-guerra, A.; De Sequeira, M.M.; Pennington, R.T.; Kenicer, G.; Carine, M.A. Systematics, biogeography, and character evolution of the legume tribe Fabeae with special focus on the middle-Atlantic island lineages. BMC Evol. Biol. 2012, 12, 250. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, G.M.A.; Carvalho, C.R.; Soares, F.A.F. Flow cytometry and cytogenetic tools in eucalypts: Genome size variation × karyotype stability. Tree Genet. Genomes 2017, 13, 106. [Google Scholar] [CrossRef]
- Van De Wouw, M.; Maxted, N.; Ford-Lloyd, B.V. A multivariate and cladistic study of Vicia L. ser. Vicia (Fabaceae) based on analysis of morphological characters. Plant Syst. Evol. 2003, 237, 19–39. [Google Scholar] [CrossRef]
- Bennett, S.J.; Maxted, N. An ecogeographic analysis of the Vicia narbonensis complex. Genet. Resour. Crop Evol. 1997, 44, 411–428. [Google Scholar] [CrossRef]
- Tabur, S.; Cesur, A.; Özkul, H. Karyology of seven fabaceae taxa from Turkey. J. Appl. Biol. Sci. 2009, 3, 45–49. [Google Scholar]
- Naranjo, C.A.; Ferrari, M.R.; Palermo, A.M.; Poggio, L. Karyotype, DNA content and meiotic behaviour in five South American species of Vicia (Fabaceae). Ann. Bot. 1998, 82, 757–764. [Google Scholar] [CrossRef] [Green Version]
- El-Bok, S.; Zoghlami-Khelil, A.; Ben-Brahim, T.; Ouji, A.; Hassen, H.; Lamine, O.; Jabri, C.; Douggari, R.; El-Gazzah, M. Chromosome number and karyotype analysis of some taxa of Vicia genus (fabaceae): Revision and description. Int. J. Agric. Biol. 2014, 16, 1067–1074. [Google Scholar]
- Castiglione, M.R.; Frediani, M.; Gelati, M.T.; Venora, G.; Giorgetti, L.; Caputo, P.; Cremonini, R. Cytological and molecular characterization of Vicia barbazitae Ten. & Guss. Protoplasma 2012, 249, 779–788. [Google Scholar]
- Castiglione, M.R.; Frediani, M.; Gelati, M.T.; Ravalli, C.; Venora, G.; Caputo, P.; Cremonini, R. Cytology of Vicia species. X. Karyotype evolution and phylogenetic implication in Vicia species of the sections Atossa, Microcarinae, Wiggersia and Vicia. Protoplasma 2011, 248, 707–716. [Google Scholar] [CrossRef]
- Inceer, H.; Hayirlioglu-Ayaz, S.; Beyazoglu, O. Cytotaxonomic investigations on some taxa of the genus Vicia L. from north-eastern Anatolia. Acta Bot. Gall. 2002, 149, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Callimassia, M.A.; Bennett, M.D. Cytotaxonomic studies of Eastern Mediterranean Vicia species (Leguminosae). Plant Syst. Evol. 1991, 177, 221–234. [Google Scholar] [CrossRef]
- Tabur, S.; Civelek, S.; Bagci, E. Cytotaxonomic studies on some Vicia L. species growing in eastern Mediterranean and southern Aegean regions, I. Acta Bot. Gall. 2001, 148, 159–174. [Google Scholar] [CrossRef]
- Caputo, P.; Frediani, M.; Gelati, M.T.; Venora, G.; Cremonini, R.; Ruffini Castiglione, M. Karyological and molecular characterisation of subgenus Vicia (Fabaceae). Plant Biosyst. 2013, 147, 1242–1252. [Google Scholar] [CrossRef]
- Arslan, E.; Ertuǧrul, K.; Öztürk, A.B. Karyological studies of some species of the genus Vicia L. (Leguminosae) in Turkey. Caryologia 2012, 65, 106–113. [Google Scholar] [CrossRef]
- McLeish, J.; Sunderland, N. Measurements of deoxyribosenucleic acid (DNA) in higher plants by Feulgen photometry and chemical methods. Exp. Cell Res. 1961, 24, 527–540. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Vrána, J.; Cápal, P.; Bednářová, M.; Doležel, J. Flow cytometry in plant research: A success story. Plant Cell Monogr. 2014, 22, 395–430. [Google Scholar]
- Galbraith, D.W.; Lambert, G.M. High-throughput monitoring of plant nuclear DNA contents via flow cytometry. Methods Mol. Biol. 2012, 918, 311–325. [Google Scholar] [PubMed]
- Doležel, J.; Greilhuber, J.; Suda, J. Flow Cytometry with Plants: An Overview. In Flow Cytometry with Plant Cells: Analysis of Genes, Chromosomes and Genomes; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 41–65. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press: Oxford, UK, 2012; ISBN 0199549060. [Google Scholar]
- Athinodorou, F.; Foukas, P.; Tsaniklidis, G.; Kotsiras, A.; Chrysargyris, A.; Delis, C.; Kyratzis, A.C.; Tzortzakis, N.; Nikoloudakis, N. Morphological diversity, genetic characterization, and phytochemical assessment of the cypriot tomato germplasm. Plants 2021, 10, 1698. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Ioannidou, S.; Nikoloudakis, N.; Seraphides, N.; Papayiannis, L.C.; Kyratzis, A.C. Physicochemical characterization and trait stability in a genetically diverse ex situ collection of pomegranate (Punica granatum L.) germplasm from Cyprus. Sci. Hortic. 2020, 263, 109116. [Google Scholar] [CrossRef]
- Kyratzis, A.C.; Nikoloudakis, N.; Katsiotis, A. Genetic variability in landraces populations and the risk to lose genetic variation. The example of landrace “Kyperounda” and its implications for ex situ conservation. PLoS ONE 2019, 14, e0224255. [Google Scholar] [CrossRef] [Green Version]
- Kyratzis, A.C.; Antoniou, C.; Papayiannis, L.C.; Graziani, G.; Rouphael, Y.; Kyriacou, M.C. Pod Morphology, Primary and Secondary Metabolite Profiles in Non-grafted and Grafted Carob Germplasm Are Configured by Agro-Environmental Zone, Genotype, and Growing Season. Front. Plant Sci. 2021, 11, 2199. [Google Scholar] [CrossRef] [PubMed]
- Anestiadou, K.; Nikoloudakis, N.; Hagidimitriou, M.; Katsiotis, A. Monumental olive trees of Cyprus contributed to the establishment of the contemporary olive germplasm. PLoS ONE 2017, 12, e0187697. [Google Scholar] [CrossRef] [Green Version]
- Grigoriou, A.; Tsaniklidis, G.; Hagidimitriou, M.; Nikoloudakis, N. The cypriot indigenous grapevine germplasm is a multi-clonal varietal mixture. Plants 2020, 9, 1034. [Google Scholar] [CrossRef] [PubMed]
- Meikle, R.D. Flora Cyprus; The Bentham-Moxon Trust: Kew, UK, 1977; Volume 1. [Google Scholar]
- Hand, R.; Hadjikyriakou, G.N.; Christodoulou, C.S. Continuously Updated: Flora of Cyprus—A Dynamic Checklist. 2011. Available online: http//www.flora-of-cyprus.eu (accessed on 3 March 2023).
- Temsch, E.M.; Koutecký, P.; Urfus, T.; Šmarda, P.; Doležel, J. Reference standards for flow cytometric estimation of absolute nuclear DNA content in plants. Cytom. Part A 2022, 101, 710–724. [Google Scholar] [CrossRef]
- Doležel, J.; Binarová, P.; Lcretti, S. Analysis of Nuclear DNA content in plant cells by Flow cytometry. Biol. Plant. 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J.; Voglmayr, H.; Greilhuber, J.; Thomas, R.A. Nuclear DNA content and genome size of trout and human (multiple letters). Cytom. Part A 2003, 51, 127–129. [Google Scholar] [CrossRef]
- Nikoloudakis, N.; Aissat, A.; Katsiotis, A. Screening A. ventricosa populations for 2n gametes. Euphytica 2018, 214, 34. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [Green Version]
- Lock, J.M.; Maxted, N. Tribe Fabeae. In Legumes of the World; Royal Botanic Gardens: Kew, UK, 2005; pp. 505–509. [Google Scholar]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World; Clarendon Press: Oxford, UK, 1993. [Google Scholar]
- Flint-Hamilton, K.B. Legumes in Ancient Greece and Rome: Food, Medicine, or Poison? Hesperia J. Am. Sch. Class. Stud. Athens 1999, 68, 371–385. [Google Scholar] [CrossRef]
- Hand, R.; Hadjikyriakou, G.N.; Christodoulou, C.S. Updated numbers of the vascular flora of Cyprus including the endemism rate. Cypricola 2019, 13, 1–6. [Google Scholar]
- Greilhuber, J.; Temsch, E.M.; Loureiro, J.C.M. Nuclear DNA Content Measurement. In Flow Cytometry with Plant Cells: Analysis of Genes, Chromosomes and Genomes; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 67–101. [Google Scholar]
- Leitch, I.J.; Bennett, M.D. Genome Size and its Uses: The Impact of Flow Cytometry. In Flow Cytometry with Plant Cells: Analysis of Genes, Chromosomes and Genomes; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 153–176. [Google Scholar]
- Kolano, B.; Siwinska, D.; Pando, L.G.; Szymanowska-Pulka, J.; Maluszynska, J. Genome size variation in Chenopodium quinoa (Chenopodiaceae). Plant Syst. Evol. 2012, 298, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Martin, S.L.; Bekele, W.A.; Latta, R.G.; Diederichsen, A.; Peng, Y.; Tinker, N.A. Genome size variation in the genus Avena. Genome 2016, 59, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Venora, G.; Blangiforti, S.; Frediani, M.; Maggini, F.; Gelati, M.T.; Ruffini Castiglione, M.; Cremonini, R. Nuclear DNA contents, rDNAs, chromatin organization, and karyotype evolution in Vicia sect. faba. Protoplasma 2000, 213, 118–125. [Google Scholar] [CrossRef]
- Raina, S.N.; Bisht, M.S. DNA amounts and chromatin compactness in Vicia. Genetica 1988, 77, 65–77. [Google Scholar] [CrossRef]
- Raina, S.N.; Rees, H. DNA variation between and within chromosome complements of Vicia species. Heredity 1983, 51, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Siljak-Yakovlev, S.; Pustahija, F.; Šolić, E.M.; Bogunić, F.; Muratović, E.; Bašić, N.; Catrice, O.; Brown, S.C. Towards a genome size and chromosome number database of balkan flora: C-values in 343 taxa with novel values for 242. Adv. Sci. Lett. 2010, 3, 190–213. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Pyšek, P.; Lučanová, M.; Wigginton, S.; Tran, C.-T.; Cronin, J.T. Plant genome size influences stress tolerance of invasive and native plants via plasticity. Ecosphere 2020, 11, e03145. [Google Scholar] [CrossRef]
- Dai, S.-F.; Zhu, X.-G.; Hutang, G.-R.; Li, J.-Y.; Tian, J.-Q.; Jiang, X.-H.; Zhang, D.; Gao, L.-Z. Genome Size Variation and Evolution Driven by Transposable Elements in the Genus Oryza. Front. Plant Sci. 2022, 13, 921937. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S.P.; Schwarzacher, T.; Liu, Q. Polyploidy: Its consequences and enabling role in plant diversification and evolution. Ann. Bot. 2023, 131, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Papolu, P.K.; Ramakrishnan, M.; Mullasseri, S.; Kalendar, R.; Wei, Q.; Zou, L.; Ahmad, Z.; Vinod, K.K.; Yang, P.; Zhou, M. Retrotransposons: How the continuous evolutionary front shapes plant genomes for response to heat stress. Front. Plant Sci. 2022, 13, 1064847. [Google Scholar] [CrossRef] [PubMed]
- Carta, A.; Peruzzi, L. Testing the large genome constraint hypothesis: Plant traits, habitat and climate seasonality in Liliaceae. New Phytol. 2016, 210, 709–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxted, N. An Ecogeographical Study of Vicia Subgenus Vicia; IPGRI (International Plant Research Institute): Rome, Italy, 1995; ISBN 978-92-9043-240-1. [Google Scholar]
- Hollings, E.; Stace, C.A. Karyotype Variation and Evolution in the Vicia Sativ A Aggregate. New Phytol. 1974, 73, 195–208. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. of Accessions | DNA Content/Data from Kew (pg/1C) | Internal Standard 3 | Mbp 4 | Mean DNA Content ± SD (pg/1C) | %CV | Range of DNA Content (pg) | HSD Test 5 | LSD Test 5 |
|---|---|---|---|---|---|---|---|---|---|
| L. culinaris | 18 | 4.21 | Z. m | 4260.983 | 4.357 ± 0.05 | 1.148 | 4.209–4.448 | ij | k |
| L. ervoides1 | 5 | - | Z. m | 3391.313 | 3.468 ± 0.026 | 0.750 | 3.437–3.494 | l | n |
| L. nigricans1 | 3 | - | Z. m | 3452.992 | 3.531 ± 0.015 | 0.425 | 3.521–3.548 | l | n |
| L. orientalis1 | 2 | - | Z. m | 4280.706 | 4.377 ± 0.062 | 1.416 | 4.333–4.421 | hij | jk |
| P. fulvum | 2 | 4.7 | Z. m | 4735.476 | 4.842 ± 0.035 | 0.723 | 4.817–4.867 | g | g |
| P. sativum subsp. biflorum | 3 | 4.75 | Z. m | 4409.802 | 4.509 ± 0.091 | 2.018 | 4.437–4.611 | hi | hi |
| P. sativum subsp. sativum | 8 | 4.9 | Z. m | 4422.272 | 4.522 ± 0.026 | 0.575 | 4.493–4.572 | h | h |
| V. amphicarpa L. 2 | 2 | 1.65 | S. l | 1617.612 | 1.654 ± 0.059 | 3.567 | 1.612–1.696 | qr | t |
| V. angustifolia L. 2 | 7 | 2.01 | G. m | 1964.662 | 2.009 ± 0.061 | 3.036 | 1.929–2.094 | p | r |
| V. bithynica L. 2 | 3 | 3.83 | Z. m | 3749.978 | 3.834 ± 0.035 | 0.913 | 3.808–3.874 | k | m |
| V. cassia Boiss1 | 6 | - | Z. m | 4343.298 | 4.441 ± 0.128 | 2.882 | 4.356–4.694 | hi | ij |
| V. cretica Boiss. & Heldr.1 | 1 | - | S. l | 1549.096 | 1.584 ± 0.007 | 0.442 | 1.577–1.591 | r | t |
| V. cypria Kotschy1 | 1 | - | P. s | 5368.995 | 5.49 ± 0.022 | 0.401 | 5.467–5.51 | f | f |
| V. dalmatica A. Kern. | 6 | 6.87 | P. s | 6720.816 | 6.872 ± 0.081 | 1.179 | 6.716–6.94 | d | d |
| V. ervilia L. 2 | 8 | 4.2 | Z. m | 4154.789 | 4.248 ± 0.025 | 0.589 | 4.22–4.301 | j | l |
| V. faba L. | 14 | 13.62 | S. c | 13,525.042 | 13.829 ± 0.11 | 0.795 | 13.66–13.98 | a | a |
| V. hybrida L. 2 | 11 | 6.78 | P. s | 6546.110 | 6.693 ± 0.118 | 1.763 | 6.542–6.971 | e | e |
| V. johannis Tamamsh2 | 1 | 6.94 | P. s | 6782.479 | 6.935 ± 0.014 | 0.202 | 6.919–6.943 | d | d |
| V. lathyroides L. 2 | 3 | 2.53 | G. m | 2477.600 | 2.533 ± 0.05 | 1.974 | 2.484–2.583 | n | p |
| V. laxiflora Brot.1 | 2 | - | G. m | 2762.850 | 2.825 ± 0.003 | 0.106 | 2.823–2.827 | m | o |
| V. lunata (Boiss. & Balansa) Boiss.2 | 5 | 1.83 | S. l | 1750.033 | 1.789 ± 0.022 | 1.230 | 1.768–1.824 | q | s |
| V. lutea L. 2 | 1 | 7.4 | P. s | 7143.214 | 7.304 ± 0.011 | 0.151 | 7.291–7.312 | c | c |
| V. narbonensis L. 2 | 4 | 6.95 | P. s | 6795.878 | 6.949 ± 0.034 | 0.489 | 6.912–6.994 | d | d |
| V. palaestina Boiss.2 | 1 | 2.24 | G. m | 2191.757 | 2.241 ± 0.019 | 0.848 | 2.223–2.26 | o | q |
| V. parviflora Cav.1 | 3 | - | G. m | 2560.078 | 2.618 ± 0.08 | 3.056 | 2.526–2.673 | mn | p |
| V. peregrina L. 2 | 6 | 8.97 | P. s | 8773.149 | 8.971 ± 0.069 | 0.769 | 8.862–9.038 | b | b |
| V. pubescens (DC.) Link2 | 6 | 2.79 | G. m | 2730.739 | 2.792 ± 0.062 | 2.221 | 2.667–2.831 | m | o |
| V. sativa L. subsp. sativa | 17 | 2.25 | S. l | 1756.891 | 1.796 ± 0.063 | 3.508 | 1.726–1.999 | q | s |
| V. villosa subsp. eriocarpa (Hausskn.) P. W. Ball | 6 | 2.28 | S. l | 1782.568 | 1.823 ± 0.075 | 4.114 | 1.748–1.947 | q | s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charalambous, I.; Ioannou, N.; Kyratzis, A.C.; Kourtellarides, D.; Hagidimitriou, M.; Nikoloudakis, N. Genome Size Variation across a Cypriot Fabeae Tribe Germplasm Collection. Plants 2023, 12, 1469. https://doi.org/10.3390/plants12071469
Charalambous I, Ioannou N, Kyratzis AC, Kourtellarides D, Hagidimitriou M, Nikoloudakis N. Genome Size Variation across a Cypriot Fabeae Tribe Germplasm Collection. Plants. 2023; 12(7):1469. https://doi.org/10.3390/plants12071469
Chicago/Turabian StyleCharalambous, Iliana, Nektaria Ioannou, Angelos C. Kyratzis, Dimitrios Kourtellarides, Marianna Hagidimitriou, and Nikolaos Nikoloudakis. 2023. "Genome Size Variation across a Cypriot Fabeae Tribe Germplasm Collection" Plants 12, no. 7: 1469. https://doi.org/10.3390/plants12071469