
Genotype by Environment Interaction in Grain Iron and Zinc Concentration and Yield of Maize Hybrids under Low Nitrogen and Optimal Conditions
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Genotype and Environmental Variance
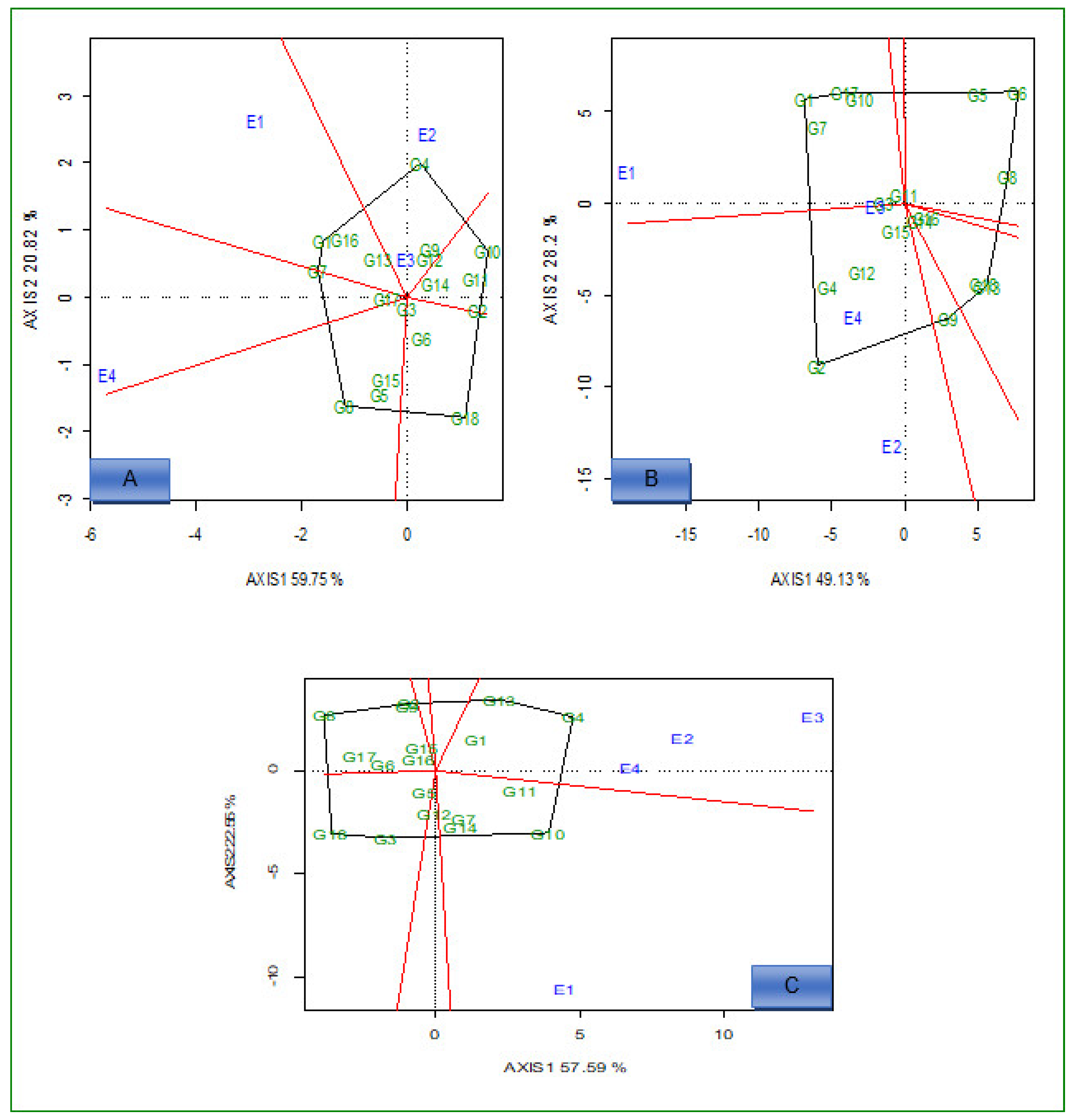
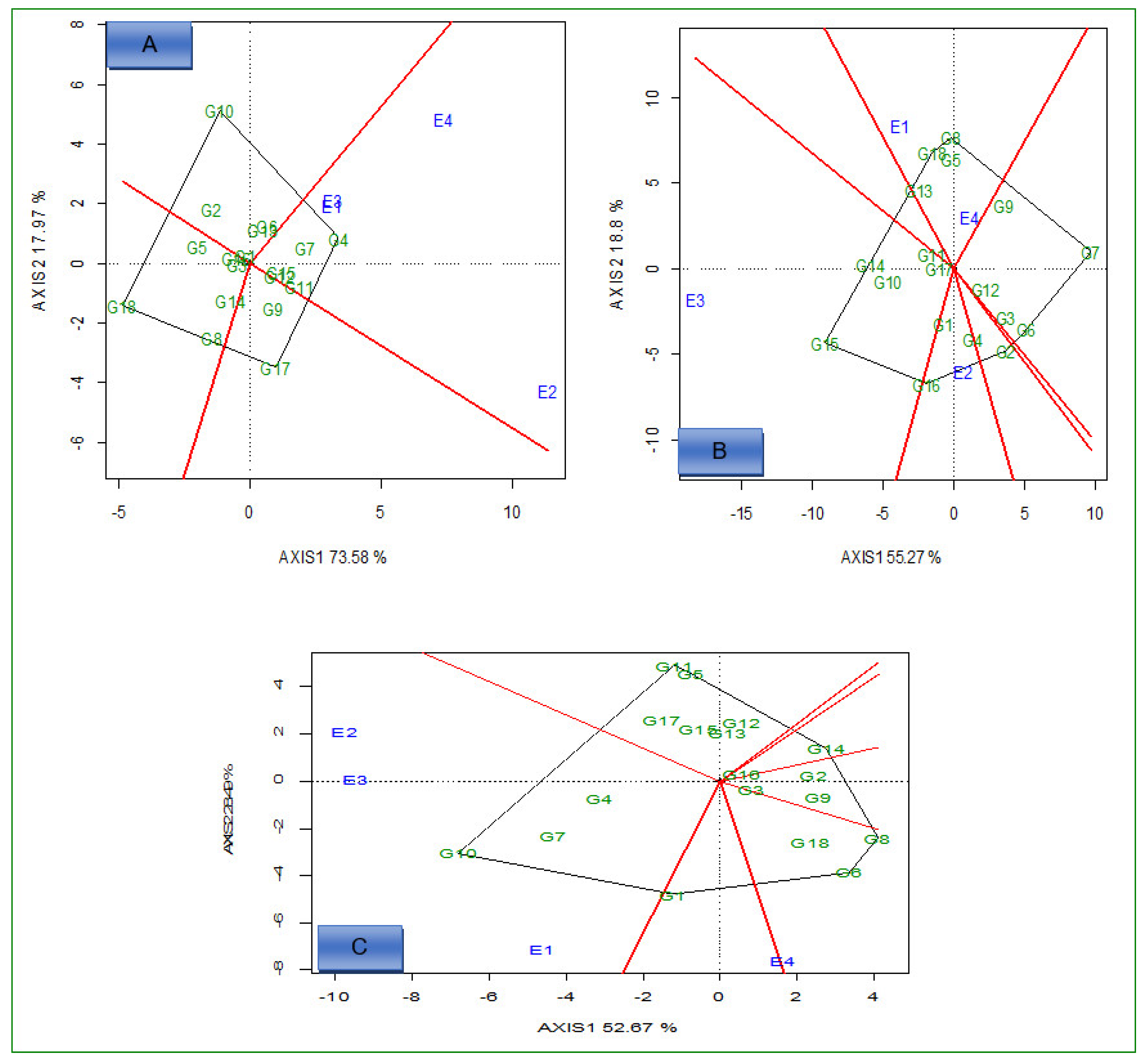
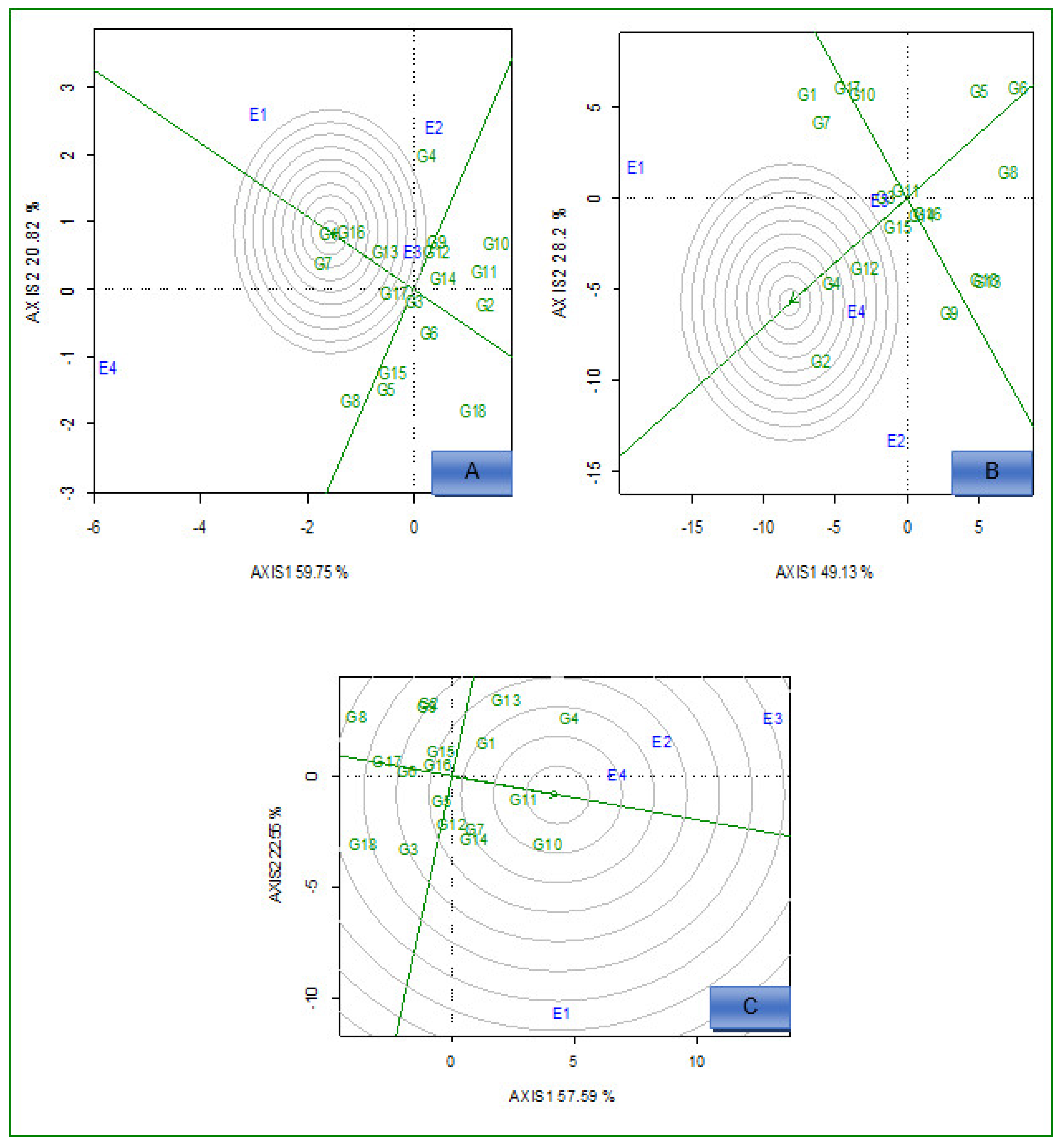
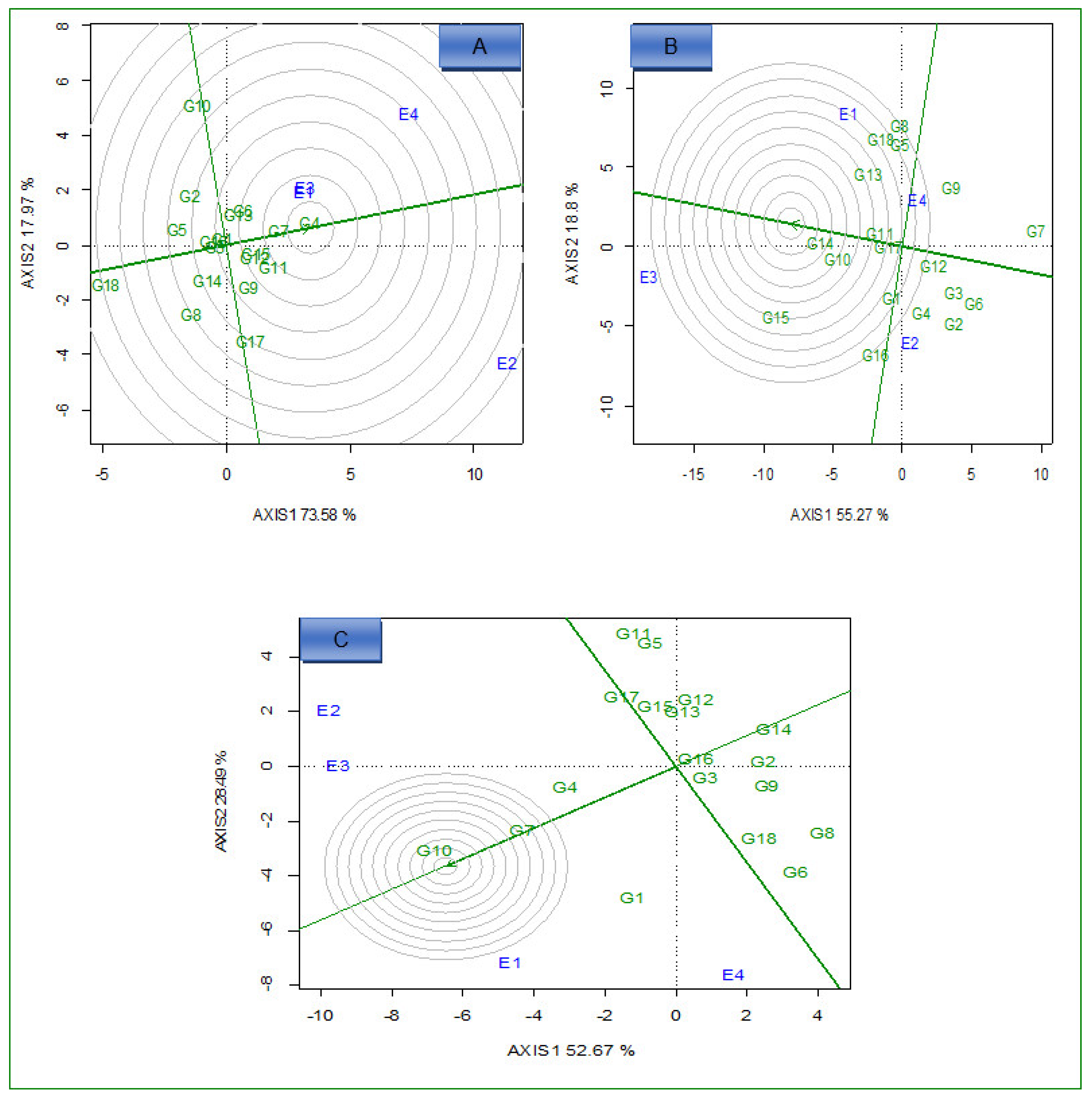
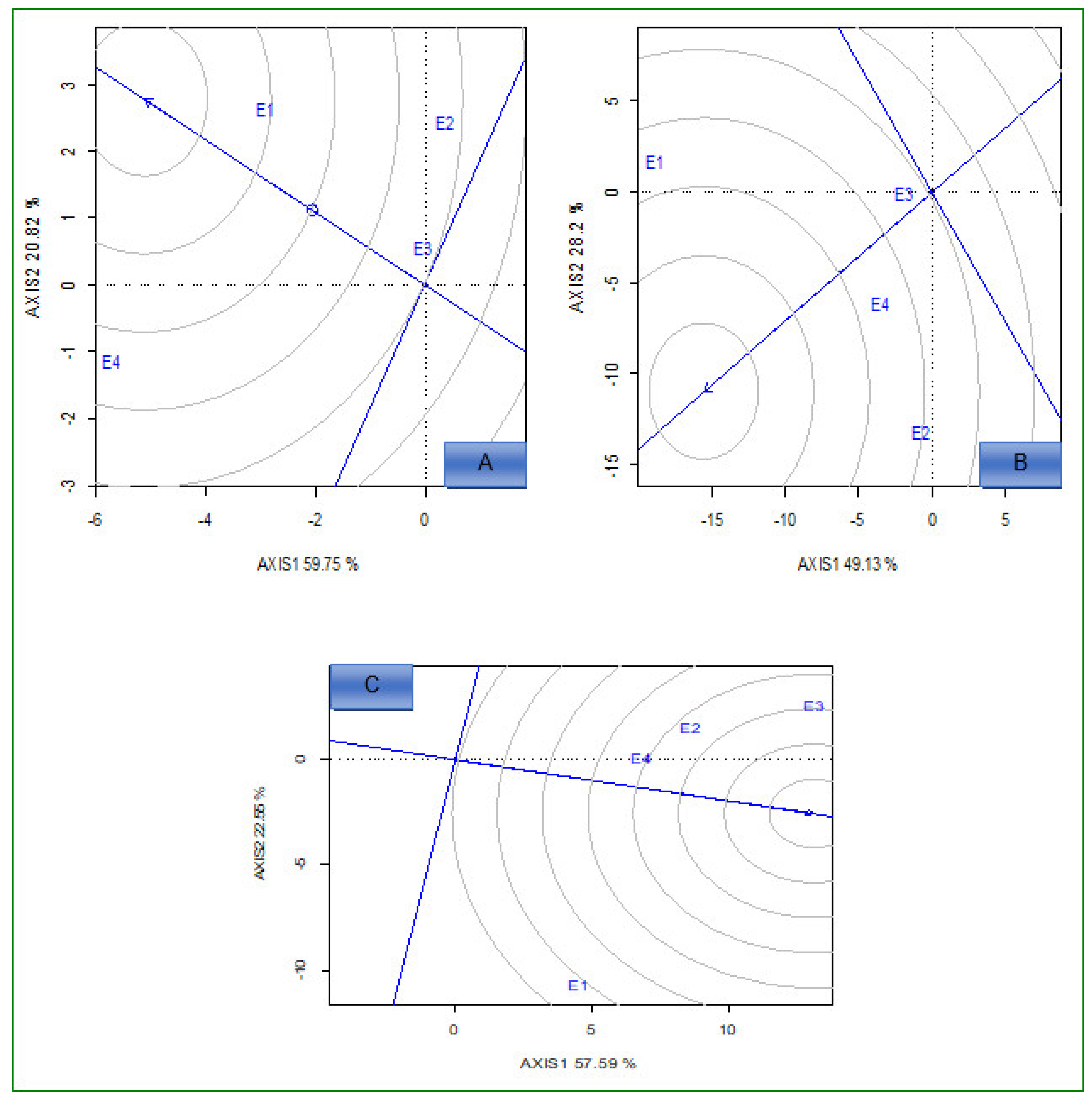
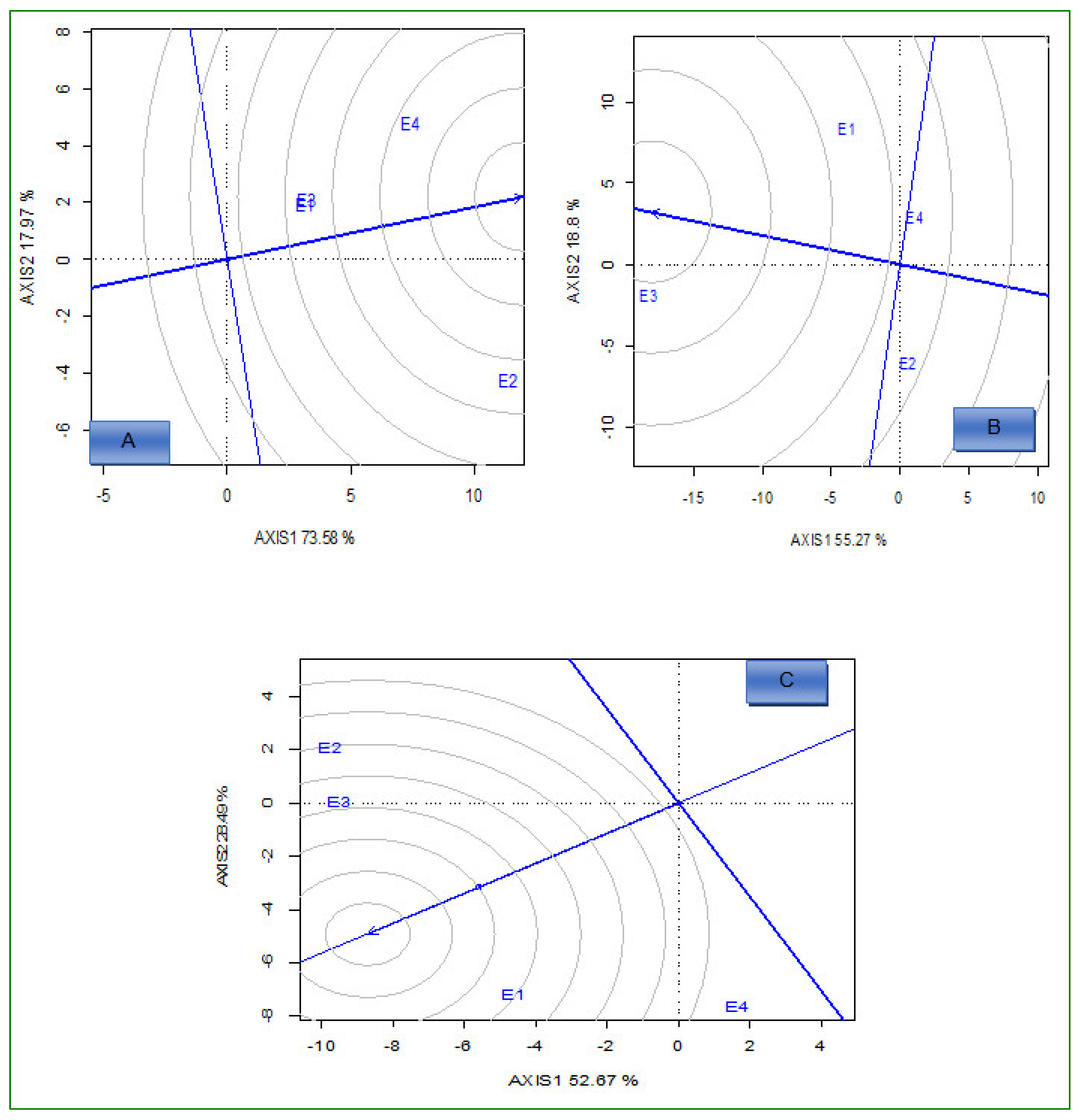
2.2. AMMI Analysis
2.3. Delineation of Mega Environment and Superior Genotype for Grain Yield, Iron, and Zinc
2.3.1. Which-Won-Where and What
2.3.2. Ideal Genotypes
2.3.3. Ideal Environments
2.4. Stability Analysis Using IPCA, AMMI Stability Value (ASV), and Yield Stability Index (YSI), Iron Stability Index (FSI), and Zinc Stability Index (ZSI)
3. Discussion
4. Materials and Methods
4.1. Descriptions of the Study Area
4.2. Plant Materials
4.3. Experimental Design and Procedures
4.4. Data Collection
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chomba, E.; Westcott, C.M.; Westcott, J.E.; Mpabalwani, E.M.; Krebs, N.F.; Patinkin, Z.W.; Palacios, N.; Michael, H.K. Zinc absorption from biofortified maize meets the requirements of young rural Zambian children. J. Nutr. 2015, 145, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Osthoff, G.; Mashingaidze, K.; Labuschagne, M. Iron and zinc in maize in the developing world: Deficiency, availability, and breeding. Crop. Sci. 2018, 58, 2200–2213. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Akinwale, R.O. Identification of early maturing maize inbred lines based on multiple traits under drought and low N environments for hybrid development and population improvement. Can. J. Plant Sci. 2011, 91, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Langade, D.M.; Shahi, J.P.; Agrawal, V.K.; Sharma, A. Maize as emerging source of oil in India: An overview. Maydica 2013, 58, 224–230. [Google Scholar]
- Babu, R.; Palacios, N.; Prasanna, B.M. Biofortified Maize—A Genetic Avenue for Nutritional Security. In Translational Genomics for Crop Breeding (Abiotic Stress, Yield and Quality), 2nd ed.; Varshney, R.K., Tuberosa, R., Eds.; John Wiley and Sons Inc.: New York, NY, USA, 2013; pp. 161–176. [Google Scholar]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Stein, A.J. Global impacts of human mineral malnutrition. Plant Soil 2010, 335, 133–154. [Google Scholar] [CrossRef]
- Hindu, V.; Palacios-Rojas, N.; Raman, B.; Suwarno, W.; Rashid, Z.; Usha, R.; Gajanan, S.R.; Nair, S. Identification and validation of genomic regions influencing kernel zinc and iron in maize. Theor. Appl. Genet. 2018, 131, 1443–1457. [Google Scholar] [CrossRef] [Green Version]
- Wessells, K.R.; Brown, K.H. Estimating the global prevalence of zinc deficiency: Results based on zinc availability in national food supplies and the prevalence of stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef] [Green Version]
- Goredema-Matongera, N.; Ndhlela, T.; Magorokosho, C.; Kamutando, C.N.; van Biljon, A.; Labuschagne, M. Multinutrient biofortification of maize (Zea mays L.) in Africa: Current status, opportunities and limitations. Nutrients 2021, 13, 1039. [Google Scholar] [CrossRef]
- Cakmak, I.; Kutman, U.B. Agronomic biofortification of cereals with zinc: A review. Eur. J. Soil Sci. 2018, 69, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Ning, S.; Zeller, M.P. Management of iron deficiency. Hematology 2019, 1, 315–322. [Google Scholar] [CrossRef]
- Jamnok, J.; Sanchaisuriya, K.; Sanchaisuriya, P.; Fucharoen, G.; Fucharoen, S.; Ahmed, F. Factors associated with anaemia and iron deficiency among women of reproductive age in Northeast Thailand: A cross-sectional study. BMC Public Health 2020, 20, 102. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, L.; Di Gaudio, F.; Consentino, B.B.; Rouphael, Y.; El-Nakhel, C.; La Bella, S.; Vasto, S.; Mauro, R.P.; D’anna, F.; Iapichino, G.; et al. Eiodine biofortification counters micronutrient deficiency and improve functional quality of open field grown curly endive. Horticulturae 2021, 7, 58. [Google Scholar] [CrossRef]
- Gibson, R.S. Zinc deficiency and human health: Etiology, health consequences, and future solutions. Plant Soil 2012, 361, 291–299. [Google Scholar] [CrossRef]
- Shahzad, Z.; Rouached, H.; Rakha, A. Combating mineral malnutrition through iron and zinc biofortification of cereals. Compr. Rev. Food Sci. Food Saf. 2014, 13, 329–346. [Google Scholar] [CrossRef]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef]
- Walker, N.J.; Schulze, R.E. An assessment of sustainable maize production under different management and climate scenarios for smallholder agro-ecosystems in KwaZulu-Natal, South Africa. Phys. Chem. Earth 2006, 31, 995–1002. [Google Scholar] [CrossRef]
- Haarhoff, S.J.; Kotzé, T.N.; Swanepoel, P.A. A prospectus for sustainability of rainfed maize production systems in South Africa. Crop. Sci. 2020, 60, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Ajala, S.O.; Olayiwola, M.O.; Ilesanmi, O.J.; Gedil, M.; Job, A.O.; Olaniyan, A.B. Assessment of genetic diversity among low-nitrogen-tolerant early generation maize inbred lines using SNP markers. S. Afr. J. Plant Soil 2019, 36, 181–188. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Cui, S.; Chang, S.X.; Jia, C.; Zhang, Q. A global synthesis of the effect of water and nitrogen input on maize (Zea mays) yield, water productivity and nitrogen use efficiency. Agric. For. Meteorol. 2019, 268, 136–145. [Google Scholar] [CrossRef]
- Semba, R.D. The rise and fall of protein malnutrition in global health. An. Nutr. Metab. 2016, 69, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amegbor, I.K.; Abe, A.; Adjebeng-Danquah, J.; Adu, G.B. Genetic analysis and yield assessment of maize hybrids under low and optimal nitrogen environments. Heliyon 2022, 8, e09052. [Google Scholar] [CrossRef]
- Liu, C.; Ma, C.; Lü, J.; Ye, Z. Yield stability analysis in maize hybrids of southwest china under genotype by environment interaction using GGE biplot. Agronomy 2022, 12, 1189. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Zhu, H.; Yu, F. Maize genotypes with different zinc efficiency in response to low zinc stress and heterogeneous zinc supply. Front. Plant Sci. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Singhal, T.; Satyavathi, C.T.; Singh, S.P.; Kumar, A.; Sankar, S.M.; Bhardwaj, C.; Mallik, M.; Bhat, J.; Anuradha, N.; Singh, N. Multi-environment quantitative trait loci mapping for grain iron and zinc content using bi-parental recombinant inbred line mapping population in pearl millet. Front. Plant Sci. 2021, 12, 736658. [Google Scholar] [CrossRef]
- Yan, W. GGE biplot—A windows application for graphical analysis of multienvironment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Gauch, H.G.; Zobel, R.W. Predictive and postdictive success of statistical analyses of yield trials. Theor. Appl. Genet. 1988, 76, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Purchase, J.L.; Hatting, H.; van Deventer, C.S. Genotype × environment interaction of winter wheat (Triticum aestivum L.) in South Africa: I. AMMI analysis of yield performance. S. Afr. J. Plant Soil 2000, 17, 95–100. [Google Scholar] [CrossRef]
- Farshadfar, E.; Mahmodi, N.; Yaghotipoor, A. AMMI stability value and simultaneous estimation of yield and yield stability in bread wheat (Triticum aestivum L.). Aust. J. Crop. Sci. 2011, 5, 1837–1844. [Google Scholar]
- Katuuramu, D.N.; Luyima, G.B.; Nkalubo, S.T.; Wiesinger, J.A.; Kelly, J.D.; Cichy, K.A. On-farm multi-location evaluation of genotype by environment interactions for seed yield and cooking time in common bean. Sci. Rep. 2020, 10, 3628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutari, B.; Sibiya, J.; Gasura, E.; Kondwakwenda, A.; Matova, P.M.; Chirwa, R. Genotype x environment interaction and stability analyses of grain yield and micronutrient (Fe and Zn) concentrations in navy bean (Phaseolus vulgaris L.) genotypes under varied production environments. Field Crops Res. 2022, 286, 108607. [Google Scholar] [CrossRef]
- Oikeh, S.O.; Menkir, A.; Maziya-Dixon, B.; Welch, R.; Glahn, R.P. Assessment of concentrations of iron and zinc and bioavailable iron in grains of early-maturing tropical maize varieties. J. Agric. Food Chem. 2003, 51, 3688–3694. [Google Scholar] [CrossRef]
- Mahendrakar, M.D.; Kumar, S.; Singh, R.B.; Rathore, A.; Potupureddi, G.; Kavi Kishor, P.B.; Gupta, R.; Srivastava, R.K. Genetic variability, genotype × environment interaction and correlation analysis for grain iron and zinc contents in recombinant inbred line population of pearl millet [Pennisetum glaucum (L). R. Br.]. Indian J. Genet. Plant Breed. 2019, 79, 545–551. [Google Scholar] [CrossRef]
- Shojaei, S.H.; Mostafavi, K.; Bihamta, M.R.; Omrani, A.; Mousavi, S.M.N.; Illés, Á.; Bojtor, C.; Nagy, J. Stability on maize hybrids based on GGE Biplot graphical technique. Agronomy 2022, 12, 394. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Mazumdar, S.; Chakraborti, M.; Hossain, F.; Manjaiah, K.M.; Agrawal, P.K.; Guleria, S.K.; Gupta, H.S. Genetic variability and genotype × environment interactions for kernel Fe and Zn concentrations in maize (Zea mays) genotypes. Indian J. Agric. Sci. 2011, 81, 81. [Google Scholar]
- Yan, W.; Rajcan, I. Biplot analysis of test sites and trait relations of soybean in Ontario. Crop. Sci. 2002, 42, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Sankar, S.M.; Singh, S.P.; Prakash, G.; Satyavathi, C.T.; Soumya, S.L.; Yadav, Y.; Sharma, L.D.; Rao, A.R.; Singh, N.; Srivastava, R.K. Deciphering genotype by environment interaction for target environmental delineation and identification of stable resistant sources against foliar blast disease of Pearl Millet. Front. Plant Sci. 2021, 12, 656158. [Google Scholar] [CrossRef]
- Agyeman, A.; Ewool, M. Genotype by environment interaction analysis of grain yield and yield components in provitamin A maize. J. Crop. Improv. 2022, 36, 1–24. [Google Scholar] [CrossRef]
- Dia, M.; Wehner, T.C.; Hassell, R.; Price, D.S.; Boyhan, G.E.; Olson, S.; King, S.; Davis, A.R.; Tolla, G.E. Genotype × environment interaction and stability analysis for watermelon fruit yield in the United States. Crop. Sci. 2016, 56, 1645–1661. [Google Scholar] [CrossRef]
- Zali, H.; Farshadfar, E.; Sabaghpour, S.H. Evaluation of genotype × environment interaction in chickpea using measures of stability from AMMI model. Ann. Biol. Res. 2012, 3, 3126–3136. [Google Scholar]
- AOAC Official Methods of Analysis. In Official Methods 985.01, 17th ed.; Association of Official Analytical Chemists: Rockville, MD, USA, 2000; Volume 1.
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons Inc.: New York, NY, USA, 1984. [Google Scholar]
- SAS. Statistical Analysis System; Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2013; p. 476. [Google Scholar]
- Pacheco, A.; Vargas, M.; Alvarado, G.; Rodríguez, F.; López, M.; Crossa, J.; Juan, B. User’s Manual GEA-R (Genotype by Environment Analysis with R ). In GEA-R; CIMMYT: El Batán, México, 2016; pp. 1–42. [Google Scholar]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G. Statistical analysis of a yield trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Ding, M.; Tier, B.; Yan, W. Application of the GGE biplot to evaluate genotype, environment and GxE interaction on Pinus radiata: A case study GGE biplot application. N. Z. J. For. Sci. 2007, 38, 132–142. [Google Scholar]
- Yan, W. GGE Biplot vs. AMMI graphs for genotype-by-environment data analysis. J. Indian Soc. Agric. Stat. 2011, 65, 183–193. [Google Scholar]
- Yan, W.; Hunt, L.A.; Sheng, Q.; Szlavnics, Z. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop. Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Tumuhimbise, R.; Melis, R.; Shanahan, P.; Kawuki, R. Genotype × environment interaction effects on early fresh storage root yield and related traits in cassava. Crop. J. 2014, 2, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Phuke, R.M.; Anuradha, K.; Radhika, K.; Jabeen, F.; Anuradha, G.; Ramesh, T.; Hariprasanna, K.; Mehtre, S.P.; Deshpande, S.P.; Anil, G.; et al. Genetic variability, genotype × environment interaction, correlation, and GGE biplot analysis for grain iron and zinc concentration and other agronomic traits in RIL population of sorghum (Sorghum bicolor L. Moench). Front. Plant Sci. 2017, 8, 712. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grain Yield | Iron | Zinc | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sum of Squares Explained (%) | Sum of Squares Explained (%) | Sum of Squares Explained (%) | |||||||||||
| Source of Variation | DF | MS | Total VE | GEI E | GEI Cum | MS | Total VE | GEI E | GEI Cum | MS | Total VE | GEI E | GEI Cum |
| Treatments | 71 | 25.04 | 22.86 | 20.61 | |||||||||
| Genotypes | 17 | 19.23 ** | 17.87 | 19.85 ** | 20.31 | 19.67 ** | 22.03 | ||||||
| Environments | 3 | 408.28 ** | 66.95 | 118.82 ** | 21.45 | 222.15 ** | 43.92 | ||||||
| Replication | 1 | 0.29 | 1.00 | 0.59 | |||||||||
| Interactions | 51 | 4.43 ** | 12.37 | 18.22 ** | 55.92 | 9.06 ** | 30.46 | ||||||
| IPCA 1 | 19 | 7.93 ** | 66.78 | 66.78 | 28.79 ** | 58.88 | 58.88 | 15.86 ** | 65.17 | 65.17 | |||
| IPCA 2 | 17 | 2.97 ** | 22.32 | 89.11 | 13.87 ** | 25.38 | 84.26 | 7.47 ** | 27.47 | 92.65 | |||
| Residuals | 15 | 1.64 | 9.75 | 2.27 | |||||||||
| Error | 68 | 0.75 | 0.51 | 0.77 | |||||||||
| Grain Yield | Iron | Zinc | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sum of Squares Explained (%) | Sum of Squares Explained (%) | Sum of Squares Explained (%) | |||||||||||
| Source of Variation | DF | MS | Total VE | GEI E | GEI Cum | MS | Total VE | GEI E | GEI Cum | MS | Total VE | GEI E | GEI Cum |
| Treatments | 71 | 2.49 | 32.83 | 17.66 | |||||||||
| Genotypes | 17 | 2.93 ** | 24.58 | 30.42 ** | 21.87 | 33.22 ** | 39.87 | ||||||
| Environments | 3 | 12.91 ** | 19.77 | 268.06 ** | 34.01 | 57.21 ** | 12.11 | ||||||
| Replication | 1 | 0.50 | 0.18 | 3.79 | |||||||||
| Interactions | 51 | 1.76 ** | 49.71 | 19.80 ** | 42.71 | 10.15 ** | 36.54 | ||||||
| IPCA 1 | 19 | 3.10 ** | 65.62 | 65.62 | 31.41 ** | 59.09 | 59.09 | 14.73 ** | 54.07 | 54.07 | |||
| IPCA 2 | 17 | 1.33 ** | 25.19 | 90.81 | 17.84 ** | 30.02 | 89.12 | 11.21 ** | 36.79 | 90.87 | |||
| Residuals | 15 | 0.53 | 7.33 | 3.15 | |||||||||
| Error | 68 | 0.26 | 0.48 | 2.18 | |||||||||
| Grain Yield | Fe Concentration in Grain | Zn Concentration in Grain | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gen | IPCA1 | IPCA2 | ASV | rASV | GY | rGY | YSI | IPCA1 | IPCA2 | ASVF | rASV | Fe | rFe | FSI | IPCA1 | IPCA2 | ASVZ | rASV | Zn | rZn | ZSI |
| G1 | −0.59 | −0.23 | 1.55 | 13 | 3.68 | 2 | 15 | −1.94 | 0.18 | 3.83 | 18 | 16.34 | 8 | 26 | 0.68 | 0.00 | 1.00 | 8 | 19.48 | 7 | 15 |
| G2 | 0.61 | 0.03 | 1.60 | 14 | 1.81 | 17 | 31 | −0.11 | 1.30 | 1.32 | 8 | 19.44 | 1 | 9 | 0.80 | −0.30 | 1.22 | 12 | 17.39 | 13 | 25 |
| G3 | −0.10 | −0.17 | 0.31 | 2 | 2.43 | 12 | 14 | −0.49 | 1.20 | 1.54 | 9 | 15.40 | 12 | 21 | −1.29 | 0.89 | 2.09 | 15 | 17.69 | 12 | 27 |
| G4 | 0.46 | −0.87 | 1.49 | 12 | 2.84 | 5 | 17 | −0.49 | 1.29 | 1.61 | 10 | 18.22 | 2 | 12 | 1.55 | −0.09 | 2.29 | 18 | 22.15 | 2 | 20 |
| G5 | −0.54 | 0.49 | 1.49 | 11 | 2.52 | 9 | 20 | 0.12 | −0.25 | 0.35 | 1 | 12.60 | 17 | 18 | −0.37 | −1.07 | 1.20 | 11 | 18.34 | 9 | 20 |
| G6 | 0.21 | 0.75 | 0.93 | 7 | 2.61 | 6 | 13 | 0.61 | −0.54 | 1.31 | 7 | 11.80 | 18 | 25 | −0.24 | 0.81 | 0.88 | 5 | 17.07 | 15 | 20 |
| G7 | −0.66 | 0.12 | 1.73 | 15 | 3.74 | 1 | 16 | −1.23 | −1.63 | 2.92 | 15 | 17.93 | 3 | 18 | −0.56 | −1.04 | 1.33 | 13 | 19.72 | 6 | 19 |
| G8 | −1.04 | 0.15 | 2.70 | 17 | 2.61 | 7 | 24 | 1.11 | −0.47 | 2.23 | 11 | 13.56 | 16 | 27 | 0.14 | −0.77 | 0.80 | 4 | 15.11 | 18 | 22 |
| G9 | 0.22 | −0.66 | 0.88 | 6 | 2.32 | 14 | 20 | 1.31 | 0.16 | 2.58 | 13 | 16.71 | 6 | 19 | 0.77 | −0.29 | 1.17 | 10 | 17.32 | 14 | 24 |
| G10 | 1.16 | 0.54 | 3.08 | 18 | 2.40 | 13 | 31 | −1.03 | −1.43 | 2.48 | 12 | 16.25 | 9 | 21 | −0.30 | 0.19 | 0.48 | 3 | 22.39 | 1 | 4 |
| G11 | 0.75 | 0.02 | 1.95 | 16 | 2.04 | 16 | 32 | 0.06 | −0.40 | 0.41 | 2 | 16.43 | 7 | 9 | 0.14 | 0.96 | 0.98 | 7 | 21.34 | 3 | 10 |
| G12 | 0.32 | −0.22 | 0.87 | 5 | 2.51 | 10 | 15 | −0.08 | 0.66 | 0.68 | 4 | 17.78 | 4 | 8 | −0.68 | 0.52 | 1.12 | 9 | 19.03 | 8 | 17 |
| G13 | 0.00 | 0.26 | 0.26 | 1 | 3.30 | 4 | 5 | 1.49 | 0.02 | 2.93 | 16 | 14.96 | 15 | 31 | 1.41 | 0.44 | 2.12 | 16 | 19.89 | 5 | 21 |
| G14 | 0.22 | −0.35 | 0.66 | 3 | 2.20 | 15 | 18 | 0.21 | 0.51 | 0.66 | 3 | 15.52 | 11 | 14 | −0.72 | −1.70 | 2.00 | 14 | 19.92 | 4 | 18 |
| G15 | −0.46 | 0.34 | 1.25 | 10 | 2.48 | 11 | 21 | 0.26 | −0.93 | 1.06 | 6 | 17.20 | 5 | 11 | 0.28 | 0.88 | 0.97 | 6 | 17.98 | 10 | 16 |
| G16 | −0.43 | −0.37 | 1.17 | 9 | 3.40 | 3 | 12 | 0.40 | −0.31 | 0.84 | 5 | 15.81 | 10 | 15 | 0.12 | −0.06 | 0.19 | 1 | 17.86 | 11 | 12 |
| G17 | −0.32 | −0.38 | 0.93 | 8 | 2.58 | 8 | 16 | −1.61 | 0.57 | 3.23 | 17 | 15.00 | 14 | 31 | −0.23 | 0.19 | 0.39 | 2 | 16.38 | 16 | 18 |
| G18 | 0.18 | 0.56 | 0.73 | 4 | 1.55 | 18 | 22 | 1.42 | 0.08 | 2.80 | 14 | 15.01 | 13 | 27 | −1.52 | 0.46 | 2.28 | 17 | 16.13 | 17 | 34 |
| Grain Yield | Fe Concentration in Grain | Zn Concentration in Grain | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gen | IPCA1 | IPCA2 | ASV | rASV | GY | rGY | YSI | IPCA1 | IPCA2 | ASVF | rFSV | Fe | rFe | FSI | IPCA1 | IPCA2 | ASVZ | rZSV | Zn | rZn | ZSI |
| G1 | 0.17 | 0.07 | 0.52 | 4 | 6.51 | 11 | 15 | 0.72 | −1.02 | 1.95 | 13 | 21.36 | 1 | 14 | −0.79 | 0.66 | 1.98 | 11 | 22.52 | 3 | 14 |
| G2 | 0.78 | −0.43 | 2.38 | 14 | 5.45 | 15 | 29 | 0.72 | −0.89 | 1.89 | 11 | 17.10 | 13 | 24 | −0.53 | 0.64 | 1.42 | 7 | 19.23 | 15 | 22 |
| G3 | 0.09 | 0.01 | 0.27 | 1 | 6.07 | 12 | 13 | 0.87 | −0.61 | 2.11 | 14 | 17.57 | 12 | 26 | −0.27 | 0.23 | 0.68 | 1 | 20.08 | 6 | 7 |
| G4 | −0.79 | −0.88 | 2.52 | 15 | 9.33 | 1 | 16 | −0.05 | −0.63 | 0.64 | 2 | 16.91 | 15 | 17 | 0.57 | −0.59 | 1.49 | 8 | 21.92 | 4 | 12 |
| G5 | 0.59 | −0.16 | 1.78 | 10 | 4.79 | 17 | 27 | 0.20 | 1.10 | 1.20 | 6 | 19.11 | 7 | 13 | 1.11 | 0.08 | 2.64 | 15 | 19.31 | 14 | 29 |
| G6 | 0.1 | −0.37 | 0.48 | 3 | 7.18 | 6 | 9 | 0.96 | −0.61 | 2.31 | 15 | 16.53 | 16 | 31 | −1.59 | 0.54 | 3.82 | 18 | 20.01 | 8 | 26 |
| G7 | −0.55 | −0.82 | 1.83 | 12 | 8.2 | 2 | 14 | 2.20 | 0.08 | 5.11 | 18 | 15.92 | 18 | 36 | 0.51 | −0.42 | 1.28 | 6 | 22.99 | 2 | 8 |
| G8 | −0.07 | 1.41 | 1.43 | 9 | 5.2 | 16 | 25 | 0.05 | 1.43 | 1.43 | 7 | 18.66 | 8 | 15 | −1.40 | 0.10 | 3.33 | 17 | 19.05 | 17 | 34 |
| G9 | −0.59 | 0.32 | 1.80 | 11 | 7.09 | 7 | 18 | 0.67 | 0.73 | 1.72 | 9 | 16.96 | 14 | 23 | −0.66 | −0.38 | 1.61 | 9 | 19.12 | 16 | 25 |
| G10 | 1.75 | −0.3 | 5.24 | 18 | 6.53 | 10 | 28 | −0.83 | −0.21 | 1.95 | 12 | 20.14 | 3 | 15 | 0.93 | −0.80 | 2.34 | 13 | 24.34 | 1 | 14 |
| G11 | −0.61 | 0.27 | 1.83 | 13 | 8.16 | 3 | 16 | −0.71 | 0.34 | 1.68 | 8 | 17.60 | 11 | 19 | 1.11 | 1.68 | 3.13 | 16 | 20.02 | 7 | 23 |
| G12 | −0.31 | 0.39 | 1.00 | 7 | 7.58 | 4 | 11 | 0.03 | 0.00 | 0.06 | 1 | 16.33 | 17 | 18 | 0.34 | 0.52 | 0.96 | 4 | 19.55 | 13 | 17 |
| G13 | 0.14 | −0.46 | 0.63 | 6 | 7.09 | 8 | 14 | −0.26 | 0.76 | 0.98 | 4 | 19.80 | 4 | 8 | 0.38 | 0.19 | 0.91 | 3 | 19.66 | 10 | 13 |
| G14 | −0.18 | 0.03 | 0.54 | 5 | 5.67 | 14 | 19 | −1.51 | 0.20 | 3.50 | 16 | 19.25 | 6 | 22 | −0.29 | −0.28 | 0.74 | 2 | 18.50 | 18 | 20 |
| G15 | −0.37 | 0.15 | 1.11 | 8 | 7.55 | 5 | 13 | −2.13 | −0.71 | 4.98 | 17 | 20.44 | 2 | 19 | 0.64 | −0.60 | 1.62 | 10 | 19.82 | 9 | 19 |
| G16 | 0.13 | −0.09 | 0.39 | 2 | 6.07 | 13 | 15 | −0.53 | −1.21 | 1.73 | 10 | 18.60 | 9 | 19 | 0.05 | −1.01 | 1.02 | 5 | 19.63 | 11 | 16 |
| G17 | −1.29 | 0.14 | 3.87 | 17 | 6.62 | 9 | 26 | −0.38 | 0.10 | 0.89 | 3 | 18.12 | 10 | 13 | 0.86 | 0.30 | 2.07 | 12 | 20.33 | 5 | 17 |
| G18 | 1.00 | 0.73 | 3.07 | 16 | 2.38 | 18 | 34 | −0.02 | 1.15 | 1.16 | 5 | 19.72 | 5 | 10 | −0.97 | −0.86 | 2.45 | 14 | 19.62 | 12 | 26 |
| Genotypes | Code | Fe and Zn of Parents | Breeding Status of the Genetic Materials |
|---|---|---|---|
| CBY075 LM-1574 × CBY358 LM-1857 | G1 | High × high | Hybrid (H1) |
| CBY075 LM-1574 × CBY104 LM-1603 | G2 | High × intermediate | Hybrid (H1) |
| CBY075 LM-1574 × CBY013 LM-1512 | G3 | High × low | Hybrid (H1) |
| CBY101 LM-1600 × CBY358 LM-1857 | G4 | High × high | Hybrid (H1) |
| CBY101 LM-1600 × CBY104 LM-1603 | G5 | High × intermediate | Hybrid (H1) |
| CBY101 LM-1600 × CBY013 LM-1512 | G6 | High × low | Hybrid (H1) |
| CBY102 LM-1601 × CBY358 LM-1857 | G7 | Intermediate × high | Hybrid (H1) |
| CBY102 LM-1601 × CBY104 LM-1603 | G8 | Intermediate × intermediate | Hybrid (H1) |
| CBY102 LM-1601 × CBY013 LM-1512 | G9 | Intermediate × low | Hybrid (H1) |
| CBY359 LM-1858 × CBY358 LM-1857 | G10 | Intermediate × high | Hybrid (H1) |
| CBY359 LM-1858 × CBY104 LM-1603 | G11 | Intermediate × intermediate | Hybrid (H1) |
| CBY359 LM-1858 × CBY013 LM-1512 | G12 | Intermediate × low | Hybrid (H1) |
| CBY017 LM-1516 × CBY358 LM-1857 | G13 | Low × high | Hybrid (H1) |
| CBY017 LM-1516 × CBY104 LM-1603 | G14 | Low × intermediate | Hybrid (H1) |
| CBY017 LM-1516 × CBY013 LM-1512 | G15 | Low × low | Hybrid (H1) |
| CBY014 LM-1513 × CBY358 LM-1857 | G16 | Low × high | Hybrid (H1) |
| CBY014 LM-1513 × CBY104 LM-1603 | G17 | Low × intermediate | Hybrid (H1) |
| CBY014 LM-1513 × CBY013 LM-1512 | G18 | Low × low | Hybrid (H1) |
| Minerals | Soil Depth | Potchefstroom | Cedara | Vaalharts | |||
|---|---|---|---|---|---|---|---|
| Optimal | Low N | Optimal | Low N | Optimal | Low N | ||
| 2016–2017 | 2016–2017 | 2016–2017 | 2016–2017 | 2016–2017 | 2016–2017 | ||
| Fe (mg kg−1) | 30 cm | 11.9 | 10.0 | 13.5 | 9.6 | 7.0 | 5.9 |
| 60 cm | 10.6 | 8.4 | 11.9 | 10.1 | 6.6 | 5.9 | |
| Zn (mg kg−1) | 30 cm | 9.4 | 9.0 | 1.3 | 3.2 | 3.3 | 2.5 |
| 60 cm | 8.6 | 5.6 | 1.4 | 2.2 | 2.9 | 2.3 | |
| P (mg kg−1) | 30 cm | 27.9 | 15.5 | 11.7 | 12.8 | 52.3 | 32.4 |
| 60 cm | 35.7 | 12.6 | 10.5 | 10.1 | 44.7 | 29.3 | |
| K (mg kg−1) | 30 cm | 278.5 | 198.4 | 77.0 | 174.5 | 123 | 163 |
| 60 cm | 314.9 | 209.7 | 70.5 | 120.0 | 114 | 149 | |
| Ca (mg kg−1) | 30 cm | 830.0 | 666.0 | 513.0 | 699.0 | 436 | 535 |
| 60 cm | 952.0 | 887.0 | 511.0 | 694.0 | 402 | 500 | |
| Mg (mg kg−1) | 30 cm | 384.9 | 328.5 | 99.0 | 166.0 | 141 | 174 |
| 60 cm | 440.7 | 438.9 | 99.5 | 154.0 | 128 | 169 | |
| Mn (mg kg−1) | 30 cm | 38.9 | 35.1 | 3.6 | 3.4 | 11.1 | 13.2 |
| 60 cm | 43.8 | 26.9 | 3.6 | 2.3 | 9.2 | 13.1 | |
| Soil pH | 30 cm | 6.5 | 6.1 | 4.3 | 4.4 | 6.0 | 6.3 |
| 60 cm | 6.6 | 6.0 | 4.4 | 4.5 | 6.1 | 6.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhtar, S.; Mekonnen, T.W.; Osthoff, G.; Mashingaidz, K.; Labuschagne, M. Genotype by Environment Interaction in Grain Iron and Zinc Concentration and Yield of Maize Hybrids under Low Nitrogen and Optimal Conditions. Plants 2023, 12, 1463. https://doi.org/10.3390/plants12071463
Akhtar S, Mekonnen TW, Osthoff G, Mashingaidz K, Labuschagne M. Genotype by Environment Interaction in Grain Iron and Zinc Concentration and Yield of Maize Hybrids under Low Nitrogen and Optimal Conditions. Plants. 2023; 12(7):1463. https://doi.org/10.3390/plants12071463
Chicago/Turabian StyleAkhtar, Sajjad, Tesfaye Walle Mekonnen, Gernot Osthoff, Kingstone Mashingaidz, and Maryke Labuschagne. 2023. "Genotype by Environment Interaction in Grain Iron and Zinc Concentration and Yield of Maize Hybrids under Low Nitrogen and Optimal Conditions" Plants 12, no. 7: 1463. https://doi.org/10.3390/plants12071463