
Optimizing Alternative Substrate for Tomato Production in Arid Zone: Lesson from Growth, Water Relations, Chlorophyll Fluorescence, and Photosynthesis

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Growth Parameters
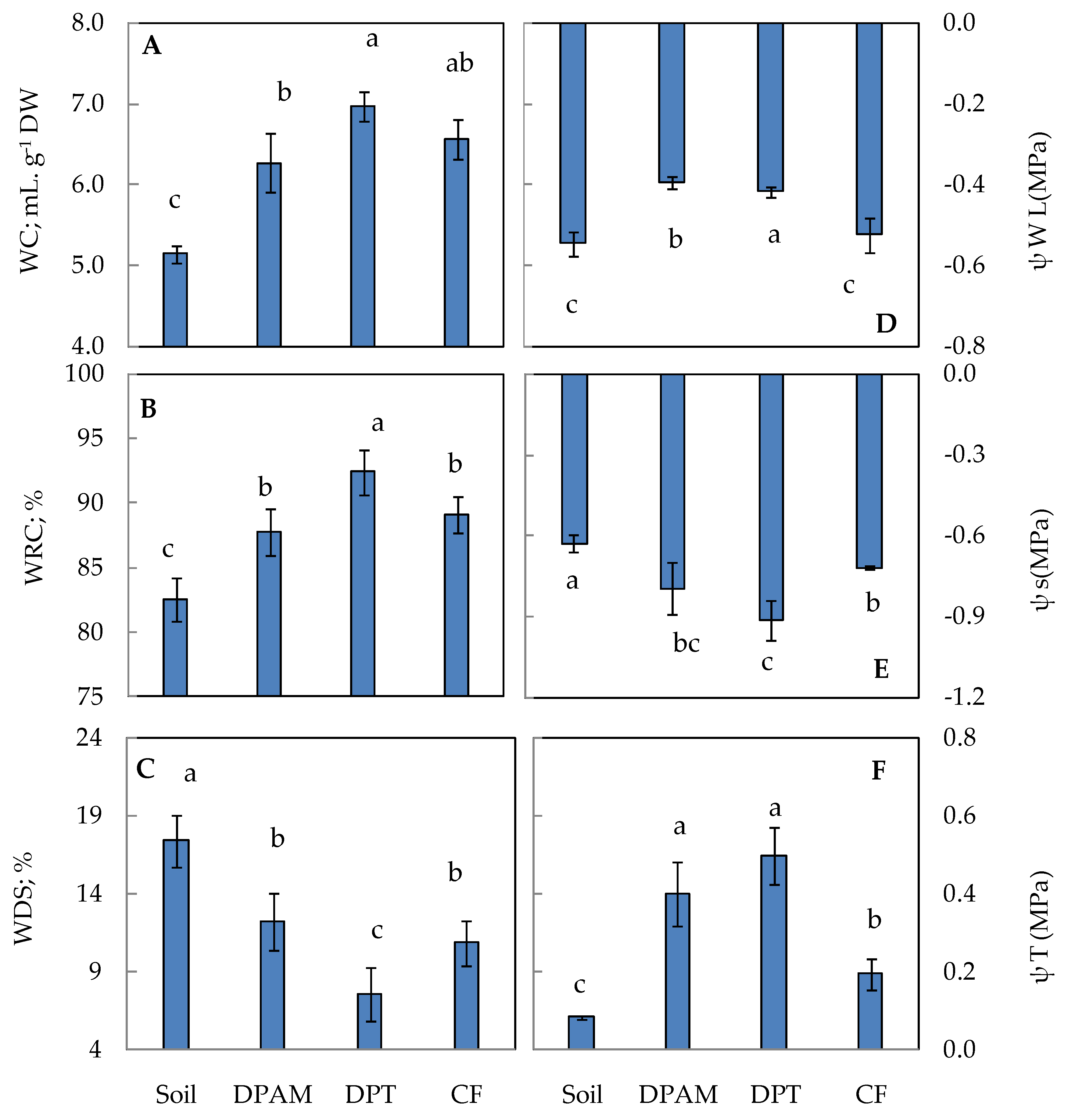
2.2. Water Status of Tomato Leaves
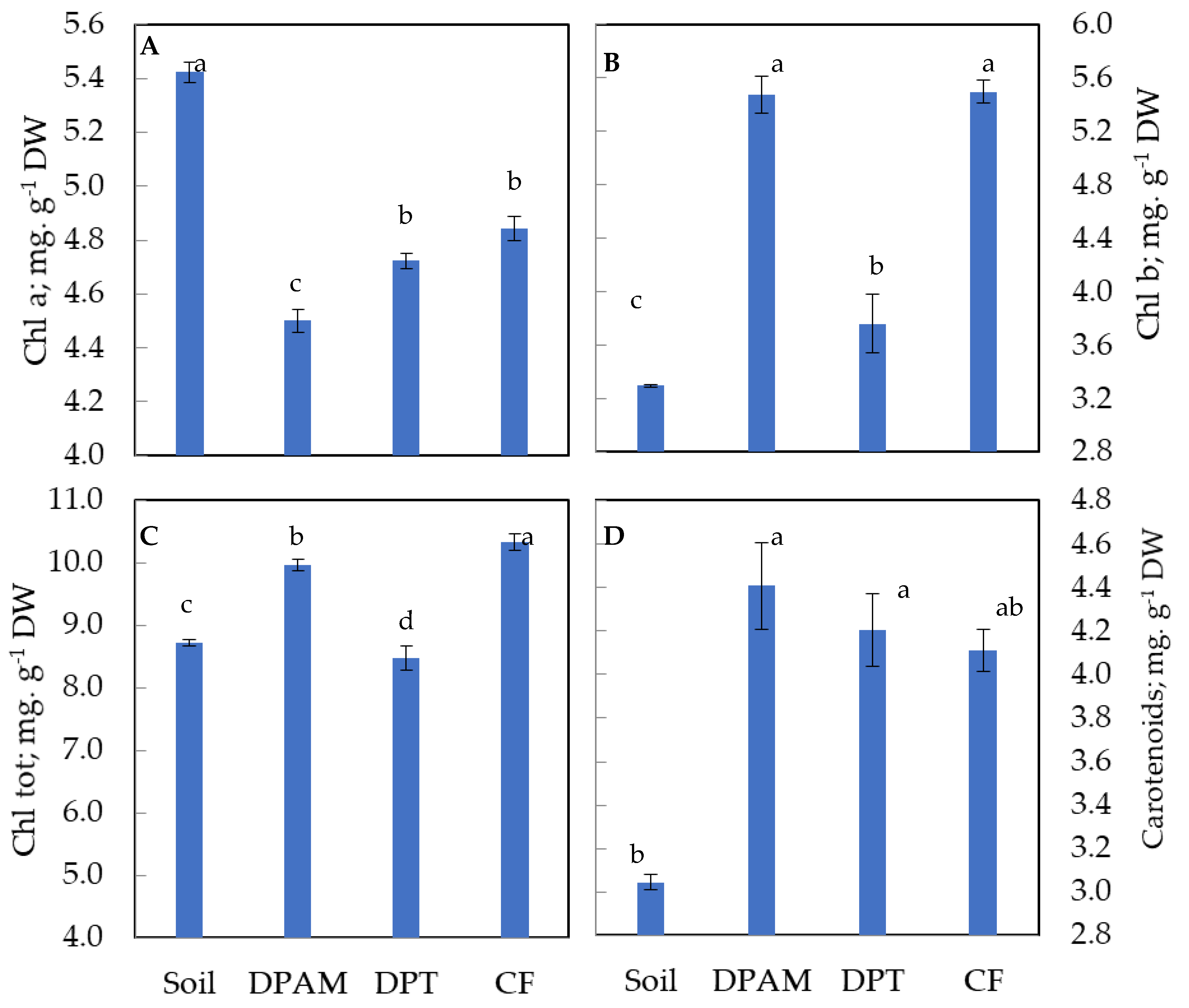
2.3. Leaves Pigment Content Assessment
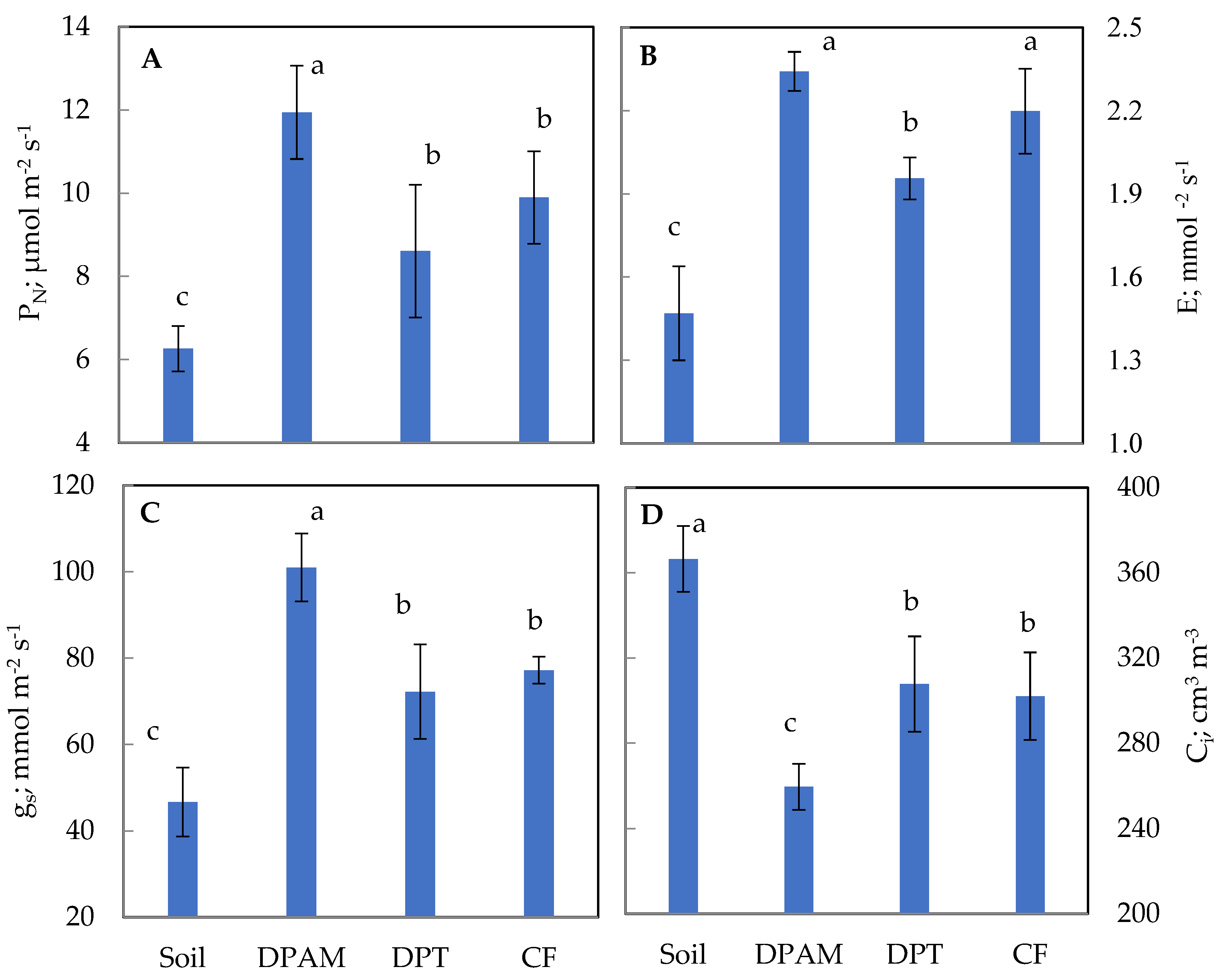
2.4. Photosynthetic Gas Exchange
2.5. Chlorophyll Fluorescence
2.6. Physico-Chemical Parameters of Tomato Fruit
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Plant Material
4.2. Physicochemical Properties of the Substrates
4.3. Plant Growth
4.4. Leaves Water Parameters
4.5. Photosynthetic Parameters:
4.5.1. Extraction, Separation, and Quantification of Pigments
4.5.2. Gas Exchanges
4.5.3. Chlorophyll Fluorescence
4.6. Physicochemical Analyses of Fruits
4.7. Preparation of Methanol Extracts
4.8. Total Polyphenols (TPC)and Flavonoids Content (TFC)
4.9. Total Antioxidant Activity (TAC)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arah, I.K.; Kumah, E.K.; Anku, E.K.; Amaglo, H. An overview of post-harvest losses in tomato production in Africa: Causes and possible prevention strategies. J. Biol. Agric. Healthc. 2015, 5, 78–88. [Google Scholar]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandić, A.; Noris, E.; Matić, S. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the Mediterranean basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- Gruda, N.S. Increasing sustainability of growing media constituents and stand-alone substrates in soilless culture systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Salhi, A.; Benabdelouahab, S.; Martin-Vide, J. Statistical analysis of long-term precipitation in the Maghreb reveals significant changes in timing and intensity. Theor. Appl. Climatol. 2022, 150, 1369–1384. [Google Scholar] [CrossRef]
- Mpandeli, S.; Nhamo, L.; Moeletsi, M.; Masupha, T.; Magidi, J.; Tshikolomo, K.; Liphadzi, S.; Naidoo, D.; Mabhaudhi, T. Assessing climate change and adaptive capacity at local scale using observed and remotely sensed data. Weather Clim. Extrem. 2019, 26, 100240. [Google Scholar] [CrossRef]
- Semerci, A.; Semerci, H.; Çalişkan, B.; Cicek, N.; Ekmekçi, Y.; Mencuccini, M. Morphological and physiological responses to drought stress of European provenances of Scots pine. Eur. J. For. Res. 2017, 136, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Waiba, K.M.; Sharma, P.; Sharma, A.; Chadha, S.; Kaur, M. Soil-less vegetable cultivation: A review. J. Pharm. Phytochem. 2020, 9, 631–636. [Google Scholar]
- Gruda, N.S. Advances in Soilless Culture and Growing Media in Today’s Horticulture—An Editorial. Agronomy 2022, 12, 2773. [Google Scholar] [CrossRef]
- Hussien, M.A.N.; Mohamed, F.H. The Impacts of different Substrates on the Growth, Yield and Fruit Quality of Greenhouse-Grown Cherry Tomato. Hortsci. J. Suez Canal Univ. 2021, 10, 73–76. [Google Scholar] [CrossRef]
- Nerlich, A.; Dannehl, D. Soilless cultivation: Dynamically changing chemical properties and physical conditions of organic substrates influence the plant phenotype of lettuce. Front. Plant Sci. 2021, 11, 601455. [Google Scholar] [CrossRef] [PubMed]
- Ghehsareh, A.M.; Hematian, M.; Kalbasi, M. Comparison of date-palm wastes and perlite as culture substrates on growing indices in greenhouse cucumber. Int. J. Recycl. Org. Waste Agric. 2012, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Dhen, N.; Abed, S.B.; Zouba, A.; Haouala, F.; Al Mohandes Dridi, B. The challenge of using date branch waste as a peat substitute in container nursery production of lettuce (Lactuca sativa L.). Int. J. Recycl. Org. Waste Agric. 2018, 7, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, M.; Bahkali, A.H. Valorization of date palm (Phoenix dactylifera) fruit processing by-products and wastes using bioprocess technology—Review. Saudi J. Biol. Sci. 2013, 20, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Ghehsareh, A.M. Effect of date palm wastes and rice hull mixed with soil on growth and yield of cucumber in greenhouse culture. Int. J. Recycl. Org. Waste Agric. 2013, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Belgacem, C.; Serra-Parareda, F.; Tarrés, Q.; Mutjé, P.; Delgado-Aguilar, M.; Boufi, S. Valorization of date palm waste for plastic reinforcement: Macro and micromechanics of flexural strength. Polymers 2021, 13, 1751. [Google Scholar] [CrossRef] [PubMed]
- Heidari, S.; Mortazavi, S.N.; Reezi, S.; Nikbakht, A. Composted palm waste as an alternative of coco peat in growing media: Effects on growth and nutritional status of Lily cut flower (Lilium spp.). J. Hortic. Postharvest Res. 2021, 4, 49–66. [Google Scholar]
- Ravindran, B.; Lee, S.R.; Chang, S.W.; Nguyen, D.D.; Chung, W.J.; Balasubramanian, B.; Mupambwa, H.A.; Arasu, M.V.; Al-Dhabi, N.A.; Sekaran, G. Positive effects of compost and vermicompost produced from tannery waste-animal fleshing on the growth and yield of commercial crop-tomato (Lycopersicon esculentum L.) plant. J. Environ. Manag. 2019, 234, 154–158. [Google Scholar] [CrossRef]
- Larounga, T.; Kossi, S.; Gbénonchi, M.; Priscilla, T.B. Effects of compost from urban solid household waste on the respiration of soil microbial flora and the yield of tomato (Lycopersicon esculentum) at the agronomic experimental station of Lome in Togo. GSC Adv. Res. Rev. 2022, 12, 042–050. [Google Scholar] [CrossRef]
- Meiliana, F.; Jumaria, N. Response to the growth of tomato plant (Solanum lycopersicum L.) to giving variation of organic fertilizers. J. Pertan. Trop. 2020, 7, 284–288. [Google Scholar]
- Stoknes, K.; Wojciechowska, E.; Jasinska, A.; Noble, R. Amelioration of composts for greenhouse vegetable plants using pasteurisedAgaricus mushroom substrate. Sustainability 2019, 11, 6779. [Google Scholar] [CrossRef] [Green Version]
- Darimani, H.S.; Adams, A.; Rahaman, A.B.A.; Nwang, J.; Yunus, R. Bio-sompost as a soil supplement to improve growth and yield of tomato (Lycopersicum esculentum). J. Agric. Chem. Environ. 2022, 11, 67–82. [Google Scholar]
- Tahir, M.A.; Akhtar, M.S.; Nakashima, Y.; Sabah, N.U.; Hassan, S.H.U. Transformation of organic waste materials as H2O gel and its influence on growth of maize (Zea mays L.). Commun. Soil Sci. Plant Anal. 2022, 53, 129–139. [Google Scholar] [CrossRef]
- Aydi, S.; Sassi Aydi, S.; Rahmani, R.; Bouaziz, F.; Souchard, J.P.; Merah, O.; Abdelly, C. Date-Palm compost as soilless substrate improves plant growth, photosynthesis, yield and phytochemical quality of greenhouse melon (Cucumis melo L.). Agronomy 2023, 13, 212. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; He, Q.; Zhou, H. Stomatal limitations to photosynthesis and their critical water conditions in different growth stages of maize under water stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Fascella, G.; Montoneri, E.; Rouphael, Y. Biowaste-derived humic-like substances improve growth and quality of orange jasmine (Murraya paniculata L. Jacq.) plants in soilless potted culture. Resources 2021, 10, 80. [Google Scholar] [CrossRef]
- Coker, C.S.; Ozores-Hampton, M. Compost tea foliar disease suppression in horticulture crops. In Compost Utilization in Production of Horticultural Crops; CRC Press: Boca Raton, FL, USA, 2021; pp. 123–134. [Google Scholar]
- Mohamed, M.H.; Sami, R.; Al-Mushhin, A.A.; Ali, M.M.E.; El-Desouky, H.S.; Ismail, K.A.; Zewail, R.M. Impacts of effective microorganisms, compost tea, fulvic acid, yeast extract, and foliar spray with seaweed extract on sweet pepper plants under greenhouse conditions. Plants 2021, 10, 1927. [Google Scholar] [CrossRef]
- Ghouili, E.; Hidri, Y.; M’Hamed, H.C.; Somenahally, A.; Xue, Q.; Znaïdi, I.E.A.; Sassi, K. Date palm waste compost promotes plant growth and nutrient transporter genes expression in barley (Hordeum vulgare L.). S. Afr. J. Bot. 2022, 149, 247–257. [Google Scholar] [CrossRef]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus × domestica Borkh). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Jawad, R.; Nawaz, A.; Ejaz, S.; Ali, S.; Saleem, M.S.; Hammad, H.M. Zeolite amendment reduces lead accumulation and improves growth and yield in tomato plants irrigated with sewage water. Environ. Sci. Pollut. Res. 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Merah, O. Carbon isotope discrimination and mineral composition of three organs in durum wheat genotypes grown under Mediterranean conditions. Comptes Rendus l’Acad. Sci.-Ser. III-Sci. Vie 2001, 324, 355–363. [Google Scholar] [CrossRef]
- Monneveux, P.; Rekika, D.; Acevedo, E.; Merah, O. Leaf gas exchange, carbon isotope discrimination, transpiration efficiency and productivity in durum wheat. Plant Sci. 2006, 170, 867–872. [Google Scholar] [CrossRef]
- Xiong, J.; Tian, Y.; Wang, J.; Liu, W.; Chen, Q. Comparison of coconut coir, rockwool, and peat cultivations for tomato production: Nutrient balance, plant growth and fruit quality. Front. Plant Sci. 2017, 8, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merah, O.; Deléens, E.; Souyris, I.; Nachit, M.M.; Monneveux, P. Stability of carbon isotope discrimination and yield across different Mediterranean environments in durum wheat. Crop Sci. 2001, 41, 677–681. [Google Scholar] [CrossRef]
- Najar, R.; Aydi, S.; Sassi Aydi, S.; Zarai, A.; Abdelly, C. Effect of salt stress on photosynthesis and chlorophyll fluorescence in Medicago truncatula. Plant Biosyst. 2019, 153, 88–97. [Google Scholar] [CrossRef]
- Foyer, C.H.; Hanke, G. ROS production and signaling in chloroplasts: Cornerstones and evolving concepts. Plant J. 2022, 111, 642–661. [Google Scholar] [CrossRef] [PubMed]
- Khanghahi, M.Y.; Leoni, B.; Crecchio, C. Photosynthetic responses of durum wheat to chemical/microbiological fertilization management under salt and drought stresses. Acta Physiol. Plant. 2021, 43, 123. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Salmon, Y.; Dietrich, L.; Sevanto, S.; Hölttä, T.; Dannoura, M.; Epron, D. Drought impacts on tree phloem: From cell-level responses to ecological significance. Tree Physiol. 2019, 39, 173–191. [Google Scholar] [CrossRef] [Green Version]
- Scarano, A.; Olivieri, F.; Gerardi, C.; Liso, M.; Chiesa, M.; Chieppa, M.; Rigano, M.M. Selection of tomato landraces with high fruit yield and nutritional quality under elevated temperatures. J. Sci. Food Agric. 2020, 100, 2791–2799. [Google Scholar] [CrossRef] [PubMed]
- Al-Kahtani, S.H.; Ahmed, M.A.; Al-Selwey, W.A.; Abdel-Razzak, H.S. Evaluation of composted agricultural crop wastes application on growth, mineral content, yield, and fruit quality of tomato. J. Exp. Biol. Agric. Sci. 2018, 6, 159–167. [Google Scholar]
- Yang, H.; Duan, Y.; Wei, Z.; Wu, Y.; Zhang, C.; Wu, W.; Lyu, L.; Li, W. Integrated physiological and metabolomic analyses reveal the differences in the fruit quality of the blueberry cultivated in three soilless substrates. Foods 2022, 11, 3965. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Q.; Su, L.; Cong, Y.; Chen, J.J.; Geng, Y.L.; Qian, C.L.; Xu, Q.; Chen, X.H.; Qi, X.H. Sugars enhance parthenocarpic fruit formation in cucumber by promoting auxin and cytokinin signaling. Sci. Hortic. 2021, 283, 110061. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Massa, D.; Vandecasteele, B. The tripartite of soilless systems, growing media, and plants through an intensive crop production scheme. Agronomy 2022, 12, 1896. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Tawaha, A.; Al-Tawaha, A.R.; Gammoh, S.; Kubow, S. Herbal yield, nutritive composition, phenolic contents and antioxidant activity of purslane (Portulaca oleracea L.) grown in different soilless media in a closed system. Ind. Crops Prod. 2019, 141, 111746. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Zain, N.A.M. Nutritional composition and antioxidant activity of leaves and rhizomes of bemban (Donax grandis) grown under glasshouse conditions. Ann. Res. Rev. Biol. 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Pouris, J.; Levizou, E.; Karatassiou, M.; Meletiou-Christou, M.-S.; Rhizopoulou, S. the influence of the partitioning of sugars, starch, and free proline in various organs of Cyclamen graecum on the biology of the species and its resistance to abiotic stressors. Plants 2022, 11, 1254. [Google Scholar] [CrossRef]
- Nielsen, S.S. Standard solutions and titratable acidity. In Food Analysis Laboratory Manual; Food Science Text Series; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Ben Khadher, T.; Aydi, S.; Mars, M.; Bouajila, J. Study on the chemical composition and the biological activities of Vitis vinifera stem extracts. Molecules 2022, 27, 3109. [Google Scholar] [CrossRef]
- Sassi Aydi, S.; Aydi, S.; Kolsi, R.B.A.; Haddeji, N.; Rahmani, R.; Ktari, N.; Bouajila, J. CO2 enrichment: Enhancing antioxidant, antibacterial and anticancer activities in Arthrospira platensis. Food Biosci. 2020, 35, 100575. [Google Scholar] [CrossRef]
- Rahmani, R.; El Arbi, K.; Sassi Aydi, S.; Hzami, A.; Tlahig, S.; Najar, R.; Aydi, S.; Debouba, M. Biochemical composition and biological activities of Salicornia europaea L. from southern Tunisia. J. Food Meas. Charact. 2022, 16, 4833–4846. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Soil | DPAM | DPT | CF |
|---|---|---|---|---|
| Shoot height (m) | 1.50 ± 0.03 c | 1.86 ± 0.06 a | 1.62 ± 0.07 b | 1.71 ± 0.04 b |
| Stem diameter (cm) | 11.60 ± 0.46 b | 14.07 ± 0.44 a | 13.47 ± 0.50 a | 13.80 ± 0.40 a |
| Leaves number (plant−1) | 26.20 ± 0.62 bc | 27.80 ± 0.82 ab | 28.73 ± 0.15 a | 27.20 ± 0.86 b |
| Inflorescence number (plant−1) | 7.07 ± 0.25 b | 7.80 ± 0.40 a | 7.13 ± 0.34 ab | 7.20 ± 0.40 a |
| Substrate | Soil | DPAM | DPT | CF |
|---|---|---|---|---|
| F0 | 133.3 ± 4.33 c | 142.6 ± 4.37 ab | 155.0 ± 4.47 a | 164.3 ± 9.57 a |
| Fm | 571.0 ± 6.91 b | 657.3 ± 16.92 a | 635.8 ± 23.75 a | 626.0 ± 13.57 a |
| Y | 0.767 ± 0.01 b | 0.783 ± 0.01 a | 0.756 ± 0.01 c | 0.737 ± 0.02 d |
| ɸexc | 0.618 ± 0.02 c | 0.640 ± 0.02 b | 0.663 ± 0.02 a | 0.642 ± 0.01 b |
| qP | 0.488 ± 0.02 d | 0.599 ± 0.01 a | 0.581 ± 0.04 b | 0.536 ± 0.07 c |
| ɸPSII | 0.302 ± 0.01 c | 0.382 ± 0.07 a | 0.387 ± 0.03 a | 0.331 ± 0.04 b |
| NPQ | 0.096 ± 0.02 a | 0.025 ± 0.04 c | 0.036 ± 0.06 b | 0.032 ± 0.01 b |
| Substrate | Soil | DPAM | DPT | CF |
|---|---|---|---|---|
| WC (mL g−1 DW) | 4.71 ± 0.08 b | 5.19 ± 0.11 a | 5.34 ± 0.09 a | 5.28 ± 0.08 a |
| Ash % | 8.05 ± 0.08 d | 8.92 ± 0.10 c | 12.99 ± 0.07 a | 9.18 ± 0.11 b |
| pH | 4.34 ± 0.05 b | 4.25 ± 0.05 bc | 4.48 ± 0.08 a | 4.31 ± 0.05 b |
| Brix ° | 7.12 ± 0.10 d | 8.73 ± 0.16 b | 7.87 ± 0.12 c | 9.87 ± 0.16 a |
| TSS | 61.17 ± 1.60 d | 71.00 ± 2.10 b | 65.33 ± 3.13 c | 81.00 ± 3.35 a |
| Titratable acidity | 1.05 ± 0.02 b | 0.90 ± 0.02 c | 1.16 ± 0.03 a | 1.13 ± 0.06 a |
| Sugar (mg g−1 DW) | 30.25 ± 1.06 b | 31.52 ± 1.20 b | 36.88 ± 1.07 a | 38.57 ± 1.23 a |
| Protein (mg g−1 DW) | 33.54 ± 1.26 d | 51.04 ± 0.59 c | 53.56 ± 1.15 b | 60 ± 1.35 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydi, S.; Sassi Aydi, S.; Marsit, A.; El Abed, N.; Rahmani, R.; Bouajila, J.; Merah, O.; Abdelly, C. Optimizing Alternative Substrate for Tomato Production in Arid Zone: Lesson from Growth, Water Relations, Chlorophyll Fluorescence, and Photosynthesis. Plants 2023, 12, 1457. https://doi.org/10.3390/plants12071457
Aydi S, Sassi Aydi S, Marsit A, El Abed N, Rahmani R, Bouajila J, Merah O, Abdelly C. Optimizing Alternative Substrate for Tomato Production in Arid Zone: Lesson from Growth, Water Relations, Chlorophyll Fluorescence, and Photosynthesis. Plants. 2023; 12(7):1457. https://doi.org/10.3390/plants12071457
Chicago/Turabian StyleAydi, Samir, Sameh Sassi Aydi, Asma Marsit, Nadia El Abed, Rami Rahmani, Jalloul Bouajila, Othmane Merah, and Chedly Abdelly. 2023. "Optimizing Alternative Substrate for Tomato Production in Arid Zone: Lesson from Growth, Water Relations, Chlorophyll Fluorescence, and Photosynthesis" Plants 12, no. 7: 1457. https://doi.org/10.3390/plants12071457