

Aggressiveness and Patulin Production in Penicillium expansum Multidrug Resistant Strains with Different Expression Levels of MFS and ABC Transporters, in the Presence or Absence of Fludioxonil
 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Aggressiveness Measurements
2.2. Validation of Patulin Determination Analytical Method
2.3. Patulin Production
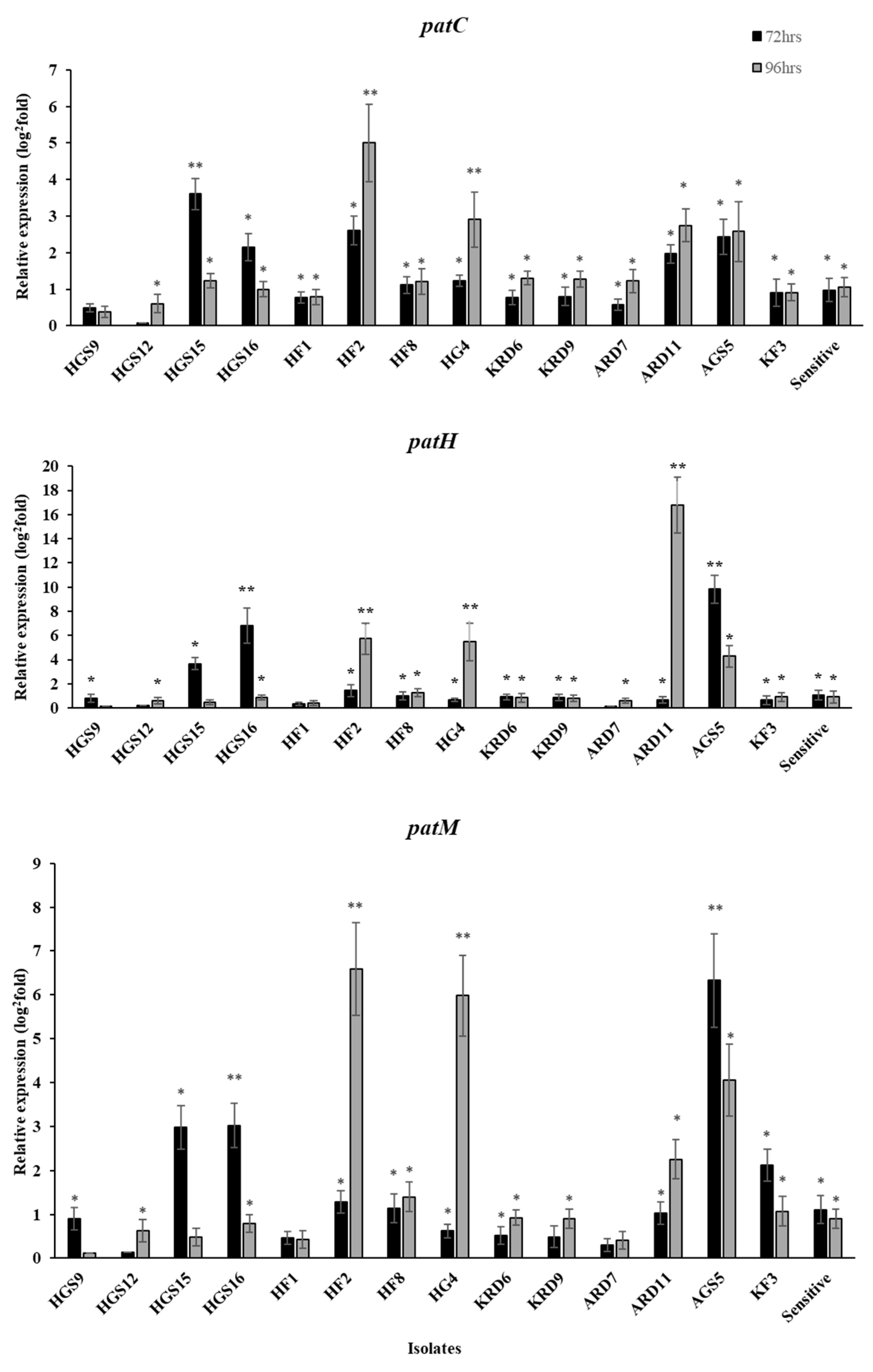
2.4. Expression Levels of Patulin Biosynthesis Genes on Fludioxonil-Treated Apple Fruit
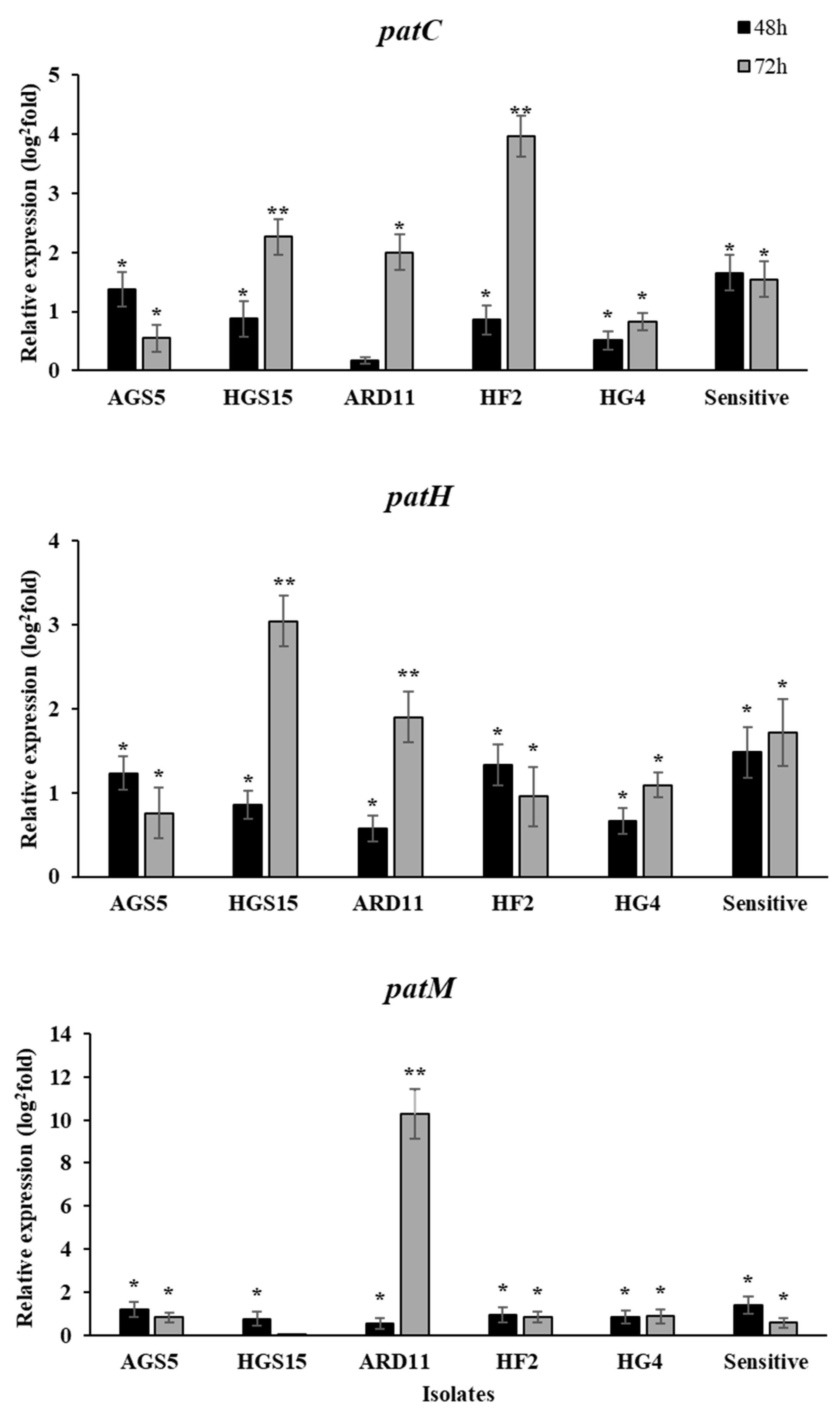
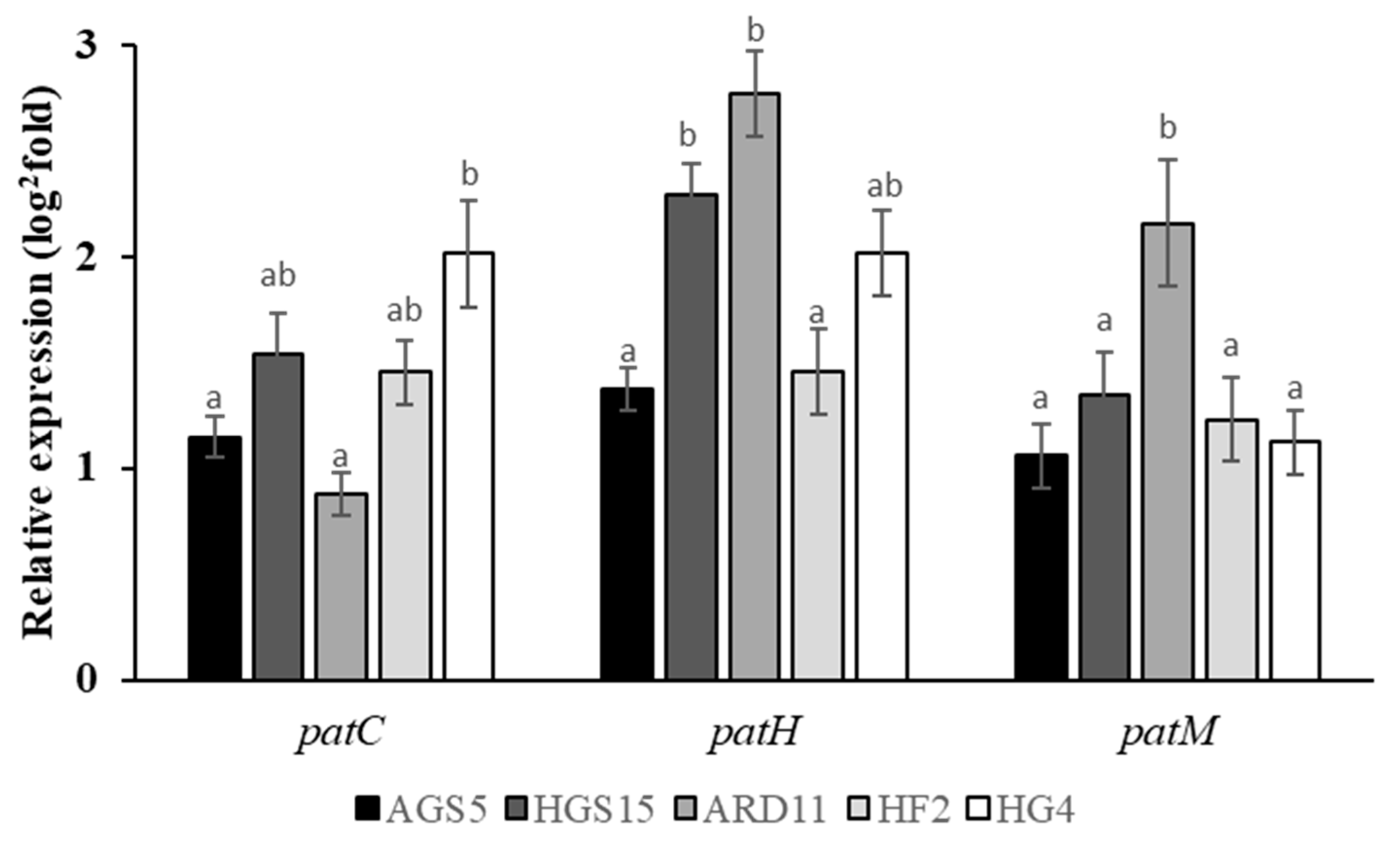
2.5. Gene Regulation of Patulin Biosynthesis Genes in MDR Isolates Grown In Vitro in the Presence of Fludioxonil
3. Discussion
4. Materials and Methods
4.1. Pathogen Isolates
4.2. Aggressiveness Measurements on Apple Fruit
4.3. Reagents and Chemicals for Mycotoxin Detection and Quantification
4.4. Patulin Extraction Procedure from Artificial Nutrient Media and Apple Fruit
4.5. Instrumental Analysis
4.6. Method Validation
4.7. Induction of Patulin Biosynthesis Related Genes In Vitro and In Vivo Samples with qRT-PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [Green Version]
- Kovac, M.; Subaric, D.; Bulaic, M.; Kovac, T.; Sarkanj, B. Yesterday masked, today modified; what do mycotoxins bring next? Arch. Ind. Hyg. Toxicol. 2018, 69, 196–214. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.J.N.D.; Schuch, P.Z.; Bernardi, C.R.; Vainstein, M.H.; Jablonski, A.; Bender, R.J. Patulin in food: State-of-the-art and analytical trends. Rev. Bras. Frutic. 2007, 29, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Sanzani, S.M.; Miazzi, M.M.; di Rienzo, V.; Fanelli, V.; Gambacorta, G.; Taurino, M.R.; Montemurro, C. A rapid assay to detect toxigenic Penicillium spp. contamination in wine and musts. Toxins 2016, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Barad, S.; Chen, Y.; Luo, X.; Tannous, J.; Dubey, A.; Matana, N.G.; Tian, S.; Li, B.; Keller, N.; et al. LaeA regulation of secondary metabolism modulates virulence in Penicillium expansum and is mediated by sucrose. Mol. Plant Pathol. 2017, 18, 1150–1163. [Google Scholar] [CrossRef]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive review of patulin control methods in foods. Compr. Rev. Food Sci. Food Saf. 2005, 4, 8–21. [Google Scholar] [CrossRef]
- Li, B.Q.; Zong, Y.Y.; Du, Z.L.; Chen, Y.; Zhang, Z.Q.; Qin, G.Z.; Tian, S.P. Genomic characterization reveals insights into patulin biosynthesis and pathogenicity in Penicillium species. Mol. Plant Microbe Interact. 2015, 28, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Feng, K.; Yang, H.; Zhang, Z.; Yuan, Y.; Yue, T. Effect of cinnamaldehyde and citral combination on transcriptional profile, growth, oxidative damage and patulin biosynthesis of Penicillium expansum. Front. Microbiol. 2018, 9, 597. [Google Scholar] [CrossRef]
- Artigot, M.P.; Loiseau, N.; Laffitte, J.; Mas-Reguieg, L.; Tadrist, S.; Oswald, I.P.; Puel, O. Molecular cloning and functional characterization of two CYP619 cytochrome P450s involved in biosynthesis of patulin in Aspergillus clavatus. Microbiology 2009, 155, 1738–1747. [Google Scholar] [CrossRef] [Green Version]
- Tannous, J.; El Khoury, R.; Snini, S.P.; Lippi, Y.; El Khoury, A.; Atoui, A.; Lteif, R.; Oswald, I.P.; Puel, O. Sequencing, physical organization and kinetic expression of the patulin biosynthetic gene cluster from Penicillium expansum. Int. J. Food Microbiol. 2014, 189, 51–60. [Google Scholar] [CrossRef]
- Li, B.Q.; Chen, Y.; Zong, Y.Y.; Shang, Y.J.; Zhang, Z.Q.; Xu, X.D.; Wang, X.; Long, M.; Tian, S.P. Dissection of patulin biosynthesis, spatial control and regulation mechanism in Penicillium expansum. Environ. Microbiol. 2019, 21, 1124–1139. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Reverberi, M.; Punelli, M.; Ippolito, A.; Fanelli, C. Study on the role of patulin on pathogenicity and virulence of Penicillium expansum. Int. J. Food Microbiol. 2012, 153, 323–331. [Google Scholar] [CrossRef]
- Ballester, A.R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lazaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; Gonzalez-Candelas, L.; Gabaldon, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [Green Version]
- Snini, S.P.; Tannous, J.; Heuillard, P.; Bailly, S.; Lippi, Y.; Zehraoui, E.; Barreau, C.; Oswald, I.P.; Puel, O. Patulin is a cultivar-dependent aggressiveness factor favouring the colonization of apples by Penicillium expansum. Mol. Plant Pathol. 2016, 17, 920–930. [Google Scholar] [CrossRef] [Green Version]
- Malandrakis, A.A.; Markoglou, A.N.; Konstantinou, S.; Doukas, E.G.; Kalampokis, J.F.; Karaoglanidis, G.S. Molecular characterization, fitness and mycotoxin production of benzimidazole-resistant isolates of Penicillium expansum. Int. J. Food Microbiol. 2013, 162, 237–244. [Google Scholar] [CrossRef]
- Ali, E.M.; Amiri, A. Selection pressure pathways and mechanism of resistance to the demethylation inhibitor-difenoconazole in Penicillium expansum. Front. Microbiol. 2018, 9, 2472. [Google Scholar] [CrossRef] [Green Version]
- Samaras, A.; Ntasiou, P.; Myresiotis, C.; Karaoglanidis, G.S. Multidrug resistance of Penicillium expansum to fungicides: Whole transcriptome analysis of MDR strains reveals overexpression of efflux transporter genes. Int. J. Food Microbiol. 2020, 335, 108896. [Google Scholar] [CrossRef]
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.S.; Fillinger, S.; Mernke, D.; Schoonbeek, H.J.; Pradier, J.M.; Leroux, P.; de Waard, M.A.; et al. Fungicide-driven evolution and molecular basis of multidrug resistance in field populations of the grey mould fungus Botrytis cinerea. PLoS Pathog. 2009, 5, e1000696. [Google Scholar] [CrossRef]
- Mernke, D.; Dahm, S.; Walker, A.-S.; Lalève, A.; Fillinger, S.; Leroch, M.; Hahn, M. Two promoter rearrangements in a drug efflux transporter gene are responsible for the appearance and spread of multi-drug resistance phenotype MDR2 in Botrytis cinerea isolates in French and German vineyards. Phytopathology 2011, 101, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Omrane, S.; Sghyer, H.; Audéon, C.; Lanen, C.; Duplaix, C.; Walker, A.-S.; Fillinger, S. Fungicide efflux and the MgMFS1 transporter contribute to the multidrug resistance phenotype in Zymoseptoria tritici field isolates. Environ. Microbiol. 2015, 17, 2805–2823. [Google Scholar] [CrossRef]
- Leroux, P.; Gredt, M.; Remuson, F.; Micoud, A.; Walker, A.S. Fungicide resistance status in French populations of the wheat eyespot fungi Oculimacula acuformis and Oculimacula yallundae. Pest Manag. Sci. 2013, 69, 15–26. [Google Scholar] [CrossRef]
- Hulvey, J.; Popko, J.T.; Sang, H.; Berg, A.; Jung, G. Overexpression of ShCYP51B and ShatrD in Sclerotinia homoeocarpa isolates exhibiting practical field resistance to a demethylation inhibitor fungicide. Appl. Environ. Microbiol. 2012, 78, 6674–6682. [Google Scholar] [CrossRef] [Green Version]
- Nakaune, R.; Adachi, K.; Nawata, O.; Tomiyama, M.; Akutsu, K.; Hibi, T. A novel ATP-binding cassette transporter involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Appl. Environ. Microbiol. 1998, 64, 3983–3988. [Google Scholar] [CrossRef] [Green Version]
- Nakaune, R.; Hamamoto, H.; Imada, J.; Akutsu, K. A novel ABC transporter gene, PMR5, is involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Mol. Genet. Genom. 2002, 267, 179–185. [Google Scholar] [CrossRef]
- Wang, J.; Sun, X.; Lin, L.; Zhang, T.; Ma, Z.; Li, H. PdMfs1, a major facilitator superfamily transporter from Penicillium digitatum, is partially involved in the imazalil-resistance and pathogenicity. Afr. J. Microbiol. Res. 2012, 6, 95–105. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Qin, T.; Li, N.; Niu, Y.; Li, D.; Yuan, Y.; Geng, H.; Xiong, L.; Liu, D. Whole transcriptome analysis of Penicillium digitatum strains treatmented with prochloraz reveals their drug-resistant mechanisms. BMC Genom. 2015, 16, 855. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Wang, S.; Yuan, Y.; Zhang, T.; Liu, J.; Liu, D. A novel major facilitator superfamily transporter in Penicillium digitatum (PdMFS2) is required for prochloraz resistance, conidiation and full virulence. Biotechnol. Lett. 2016, 38, 1349–1357. [Google Scholar] [CrossRef]
- de Ramon-Carbonell, M.; Sanchez-Torres, P. Involvement of Penicillium digitatum PdSUT1 in fungicide sensitivity and virulence during citrus fruit infection. Microbiol. Res. 2017, 203, 57–67. [Google Scholar] [CrossRef]
- Bissinger, P.H.; Kuchler, K. Molecular cloning and expression of the S. cerevisiae STS1 gene product. J. Biol. Chem. 1994, 269, 4180–4186. [Google Scholar] [CrossRef]
- Callahan, T.M.; Rose, M.S.; Meade, M.J.; Ehrenshaft, M.; Upchurch, R.G. CFP, the putative cercosporin transporter of Cercospora kikuchii, is required for wild type cercosporin production, resistance, and virulence on soybean. Mol. Plant Microbe Interact. 1999, 12, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Choquer, M.; Lee, M.H.; Bau, H.J.; Chung, K.R. Deletion of a MFS transporter-like gene in Cercospora nicotianae reduces cercosporin toxin accumulation and fungal virulence. FEBS Lett. 2007, 581, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Amnuaykanjanasin, A.; Daub, M.E. The ABC transporter ATR1 is necessary for efflux of the toxin cercosporin in the fungus Cercospora nicotianae. Fungal Genet. Biol. 2009, 46, 146–158. [Google Scholar] [CrossRef]
- Menke, J.; Dong, Y.; Kistler, H.C. Fusarium graminearum Tri12p influences virulence to wheat and trichothecene accumulation. Mol. Plant Microbe Interact. 2012, 25, 1408–1418. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Son, H.; Lee, J.; Lee, Y.R.; Lee, Y.W. A putative ABC transporter gene, ZRA1, is required for zearalenone production in Gibberella zeae. Curr. Genet. 2011, 57, 343–351. [Google Scholar] [CrossRef]
- Chang, P.K.; Yu, J.; Yu, J.H. aflT, a MFS transporter-encoding gene located in the aflatoxin gene cluster, does not have a significant role in aflatoxin secretion. Fungal Genet. Biol. 2004, 41, 911–920. [Google Scholar] [CrossRef]
- FRAC (Fungicide Resistance Action Committee). Pathogen Risk List. Available online: https://www.frac.info/docs/default-source/publications/pathogen-risk/frac-pathogen-list-2019.pdf (accessed on 20 March 2022).
- Parnell, S.; Gilligan, C.A.; Van den Bosch, F. Small-scale fungicide spray heterogeneity and the coexistence of resistant and sensitive pathogen strains. Phytopathology 2005, 95, 632–639. [Google Scholar] [CrossRef] [Green Version]
- Karaoglanidis, G.S.; Markoglou, A.N.; Bardas, G.A.; Doukas, E.G.; Konstantinou, S.; Kalampokis, J.F. Sensitivity of Penicillium expansum field isolates to tebuconazole, iprodione, fludioxonil and cyprodinil and characterization of fitness parameters and patulin production. Int. J. Food Microbiol. 2011, 145, 195–204. [Google Scholar] [CrossRef]
- de Waard, M.A.; Andrade, A.C.; Hayashi, K.; Schoonbeek, H.J.; Stergiopoulos, I.; Zwiers, L.H. Impact of fungal drug transporters on fungicide sensitivity, multidrug resistance and virulence. Pest Manag. Sci. 2006, 62, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Rahman, T.; Yarnall, B.; Doyle, D.A. Efflux drug transporters at the forefront of antimicrobial resistance. Eur. Biophys. J. 2017, 46, 647–653. [Google Scholar] [CrossRef]
- Hardy, A.; Silva-Fernandes, A.; Speijers, G.; Hans, R.; Delcour, M.P.; Kuiper, H.; Führ, F.; Carere, A.; Richard-Molard, D.; Thomas, M. Opinion on the Relationship between the Use of Plant Protection Products on Food Plants and the Occurrence of Mycotoxins in Foods; SCP/RESI/063-Final; Scientific Committee on Plants; European Commission Health and Consumer Protection Directorate-General: Brussels, Belgium, 1999; Available online: https://ec.europa.eu/food/system/files/2020-12/sci-com_scp_out56_en.pdf (accessed on 15 May 2022).
- Commission Regulation (EC). No. 1881/2006, of 19 December 2006. Setting Maximum Levels for Certain Contaminants in Foodstuffs. Off. J. Eur. Union 2006, L364, 5–24. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:02006R1881-20140701&from=EN (accessed on 8 January 2023).
- D’Mello, J.P.F.; MacDonald, A.M.C.; Postel, D.; Dijksma, W.T.P.; Dujardin, A.; Placinta, C.M. Pesticide use and mycotoxin production in Fusarium and Aspergillus phytopathogens. Eur. J. Plant Pathol. 1998, 104, 741–751. [Google Scholar] [CrossRef]
- D’Mello, J.P.F.; Macdonald, A.M.C.; Briere, L. Mycotoxin production in a carbendazim-resistant strain of Fusarium sporotrichioides. Mycotoxin Res. 2000, 16, 101–111. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Yu, J.J.; Zhang, Y.N.; Zhang, X.; Cheng, C.J.; Wang, J.X.; Hollomon, D.W.; Fan, P.S.; Zhou, M.G. Effect of carbendazim resistance on trichothecene production and aggressiveness of Fusarium graminearum. Mol. Plant-Microbe Interact. 2009, 22, 1143–1150. [Google Scholar] [CrossRef] [Green Version]
- Doukas, E.G.; Markoglou, A.N.; Ziogas, B.N. Fungicide resistance and aflatoxin production: The effect of resistance mutations to triazoles, phenylpyrroles and anilinopyrimidine fungicides on aflatoxigenic ability of Aspergillus parasiticus. In Proceedings of the DPG-BCPC Symposium-Plant Protection and Plant Health in Europe, Berlin, Germany, 10–12 May 2007. [Google Scholar]
- Doukas, E.G.; Markoglou, A.N.; Ziogas, B.N. Biochemical and molecular study of triazole-resistance and its effect on ochratoxin production by Aspergillus ochraceus Wihl. In Proceedings of the 9th International Congress of Plant Pathology, Torino, Italy, 24–29 August 2008. [Google Scholar]
- Markoglou, A.N.; Doukas, E.G.; Ziogas, B.N. Phenylpyrrole-resistance and aflatoxin production in Aspergillus parasiticus Speare. Int. J. Food Microbiol. 2008, 127, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Markoglou, A.N.; Vattis, K.; Dimitriadis, K.; Doukas, E.G.; Ziogas, B.N. Effect of phenylpyrrole resistance mutations on mycotoxin production by Aspergillus carbonarius and Penicillium expansum. In Proceedings of the 9th International Congress of Plant Pathology, Torino, Italy, 24–29 August 2008. [Google Scholar]
- Malandrakis, A.A.; Vattis, K.N.; Markoglou, A.N.; Karaoglanidis, G.S. Characterization of boscalid-resistance conferring mutations in the SdhB subunit of respiratory complex II and impact on fitness and mycotoxin production in Penicillium expansum laboratory strains. Pestic. Biochem. Physiol. 2017, 138, 97–103. [Google Scholar] [CrossRef]
- Qiu, J.B.; Yu, M.Z.; Yin, Q.; Xu, J.H.; Shi, J.R. Molecular characterization, fitness, and mycotoxin production of Fusarium asiaticum strains resistant to fludioxonil. Plant Dis. 2018, 102, 1759–1765. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.B.; Tao, X.; Zhao, H.H.; Xiao, X.M.; Li, M.; Wang, J.X.; Zhou, M.G. Activity of demethylation inhibitor fungicide metconazole on Chinese Fusarium graminearum species complex and its application in carbendazim resistance management of Fusarium head blight in wheat. Plant Dis. 2019, 103, 929–937. [Google Scholar] [CrossRef]
- Duan, Y.; Lu, F.; Zhou, Z.; Zhao, H.; Zhang, J.; Mao, Y.; Li, M.; Wang, J.; Zhou, M. Quinone outside inhibitors affect don biosynthesis, mitochondrial structure and toxisome formation in Fusarium graminearum. J. Hazard. Mater. 2020, 398, 122908. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Duan, Y.B.; Zhou, M.G. Carbendazim-resistance associated β2-tubulin substitutions increase deoxynivalenol biosynthesis by reducing the interaction between β2-tubulin and IDH3 in Fusarium graminearum. Environ. Microbiol. 2020, 22, 598–614. [Google Scholar] [CrossRef]
- Paterson, R.R.M. Some fungicides and growth inhibitor/biocontrolenhancer 2-deoxy-D-glucose increase patulin from Penicillium expansum strains in vitro. Crop Prot. 2007, 26, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Morales, H.; Marín, S.; Ramos, A.J.; Sanchis, V. Influence of post-harvest technologies applied during cold storage of apples in Penicillium expansum growth and patulin accumulation: A review. Food Control 2010, 21, 953–962. [Google Scholar] [CrossRef]
- Ramirez, M.L.; Chulze, S.; Magan, N. Impact of environmental factors and fungicides on growth and deoxinivalenol production by Fusarium graminearum isolates from Argentinian wheat. Crop Prot. 2004, 23, 117–125. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; Zwiers, L.H.; De Waard, M.A. Secretion of Natural and Synthetic Toxic Compounds from Filamentous Fungi by Membrane Transporters of the ATP-binding Cassette and Major Facilitator Superfamily. Eur. J. Plant Pathol. 2002, 108, 719–734. [Google Scholar] [CrossRef]
- Zong, Y.; Li, B.; Tian, S. Effects of carbon, nitrogen and ambient pH on patulin production and related gene expression in Penicillium expansum. Int. J. Food Microbiol. 2015, 206, 102–108. [Google Scholar] [CrossRef]
- Hayashi, K.; Schoonbeek, H.J.; De Waard, M.A. Bcmfs1, a novel major facilitator superfamily transporter from Botrytis cinerea, provides tolerance towards the natural toxic compounds camptothecin and cercosporin and towards fungicides. Appl. Environ. Microbiol. 2002, 68, 4996–5004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banani, H.; Marcet-Houben, M.; Ballester, A.R.; Abbruscato, P.; González-Candelas, L.; Gabaldón, T.; Spadaro, D. Genome sequencing and secondary metabolism of the postharvest pathogen Penicillium griseofulvum. BMC Genom. 2016, 17, 19. [Google Scholar] [CrossRef] [Green Version]
- Andersen, B.; Smedsgaard, J.; Frisvad, J.C. Penicillium expansum: Consistent production of patulin, chaetoglobosins, and other secondary metabolites in culture and their natural occurrence in fruit products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef]
- Anastassiades, M.; Lehotay, S.J.; Štajnbaher, D.; Schenck, F.J. Fast and easy multiresidue method employing acetonitrile extraction/partitioning and “dispersive solid-phase extraction” for the determination of pesticide residues in produce. J. AOAC Int. 2003, 86, 412–431. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Phenotype | Lesion Diameter (cm) | Control Efficacy % | |
|---|---|---|---|---|
| Untreated Fruit | Fludioxonil-Treated Fruit | |||
| HGS9 | MDR | 2 | 1.6 | 20 |
| HF2 | 2 | 1.6 | 20 | |
| HF8 | 3 | 2 | 33.3 | |
| HF1 | 2.8 | 2 | 28.5 | |
| KRD6 | 3 | 2 | 33.3 | |
| KRD9 | 2 | 1.5 | 25 | |
| ARD7 | 2.8 | 2 | 28.5 | |
| ARD11 | 3 | 2.5 | 16.6 | |
| HG4 | 2.7 | 2 | 25.9 | |
| HGS12 | 3 | 2.4 | 20 | |
| HGS15 | 3 | 2.5 | 16.6 | |
| HGS16 | 3 | 2.5 | 16.6 | |
| KF3 | 3 | 2.3 | 23.3 | |
| AGS5 | 3 | 2.3 | 23.3 | |
| Group mean | 2.7a ** | 2.1a | 23.67a | |
| ZRD13 | Sensitive | 3.4 | 1.5 | 55.8 |
| KF14 | 3.4 | 1.5 | 55.8 | |
| HF5 | 3.5 | 1.7 | 51.4 | |
| HRD7 | 3.5 | 1.5 | 57.1 | |
| Group mean | 3.5b | 1.55a | 55.08b | |
| Isolate | Phenotype | In Vitro (μg/g) a | In Vivo (μg/g) b | ||
|---|---|---|---|---|---|
| Control Medium | Fludioxonil-Amended Medium | Untreated Fruit | Fludioxonil-Treated Fruit | ||
| HGS9 | MDR | 587.8 | 415.5 | 3.38 | 25.62 |
| HF2 | 826.1 | 711.4 | 2.45 | 13.56 | |
| HF8 | 790.1 | 622.3 | 3.38 | 13.12 | |
| HF1 | 518.7 | 425.6 | 3.5 | 4.26 | |
| KRD6 | 616.4 | 504.6 | 9.5 | 8.6 | |
| KRD9 | nd c | nd | 3.73 | 52.4 | |
| ARD7 | 576.8 | 489.5 | 11.18 | 4.6 | |
| ARD11 | 324.5 | 277.2 | 0.2 | 0.8 | |
| HG4 | 364.5 | 231.5 | 31.3 | 58.58 | |
| HGS12 | 351.7 | 347.5 | 32.5 | 19.86 | |
| HGS15 | 441.2 | 441 | 27.25 | 100.5 | |
| HGS16 | 252.7 | 205.7 | 18.49 | 130 | |
| KF3 | 396.2 | 212.7 | 5.54 | 40.3 | |
| AGS5 | 466.8 | 315.2 | 7.31 | 1.7 | |
| Group mean | 465.2 ± 57.3a d | 371.4 ± 49.3a | 11.4 ±3.0a * | 25.5 ± 10.2a | |
| ZRD13 | Sensitive | 409.8 | 388.2 | 1.68 | 5.66 |
| KF14 | 210.7 | 237.8 | 5.45 | 15.38 | |
| HF5 | 321.2 | 355.3 | 0.34 | 15.86 | |
| HRD7 | 372.4 | 328 | 13.35 | 30.2 | |
| Group mean | 328.5 ± 43.2b | 327.3 ± 32.2a | 5.2 ± 2.9a * | 16.7 ± 5.0a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntasiou, P.; Samaras, A.; Papadakis, E.-N.; Menkissoglu-Spiroudi, U.; Karaoglanidis, G.S. Aggressiveness and Patulin Production in Penicillium expansum Multidrug Resistant Strains with Different Expression Levels of MFS and ABC Transporters, in the Presence or Absence of Fludioxonil. Plants 2023, 12, 1398. https://doi.org/10.3390/plants12061398
Ntasiou P, Samaras A, Papadakis E-N, Menkissoglu-Spiroudi U, Karaoglanidis GS. Aggressiveness and Patulin Production in Penicillium expansum Multidrug Resistant Strains with Different Expression Levels of MFS and ABC Transporters, in the Presence or Absence of Fludioxonil. Plants. 2023; 12(6):1398. https://doi.org/10.3390/plants12061398
Chicago/Turabian StyleNtasiou, Panagiota, Anastasios Samaras, Emmanouil-Nikolaos Papadakis, Urania Menkissoglu-Spiroudi, and George S. Karaoglanidis. 2023. "Aggressiveness and Patulin Production in Penicillium expansum Multidrug Resistant Strains with Different Expression Levels of MFS and ABC Transporters, in the Presence or Absence of Fludioxonil" Plants 12, no. 6: 1398. https://doi.org/10.3390/plants12061398