
Essential Oil from Glossogyne tenuifolia Inhibits Lipopolysaccharide-Induced Inflammation-Associated Genes in Macro-Phage Cells via Suppression of NF-κB Signaling Pathway
 , ,
, ,
Abstract
:1. Introduction
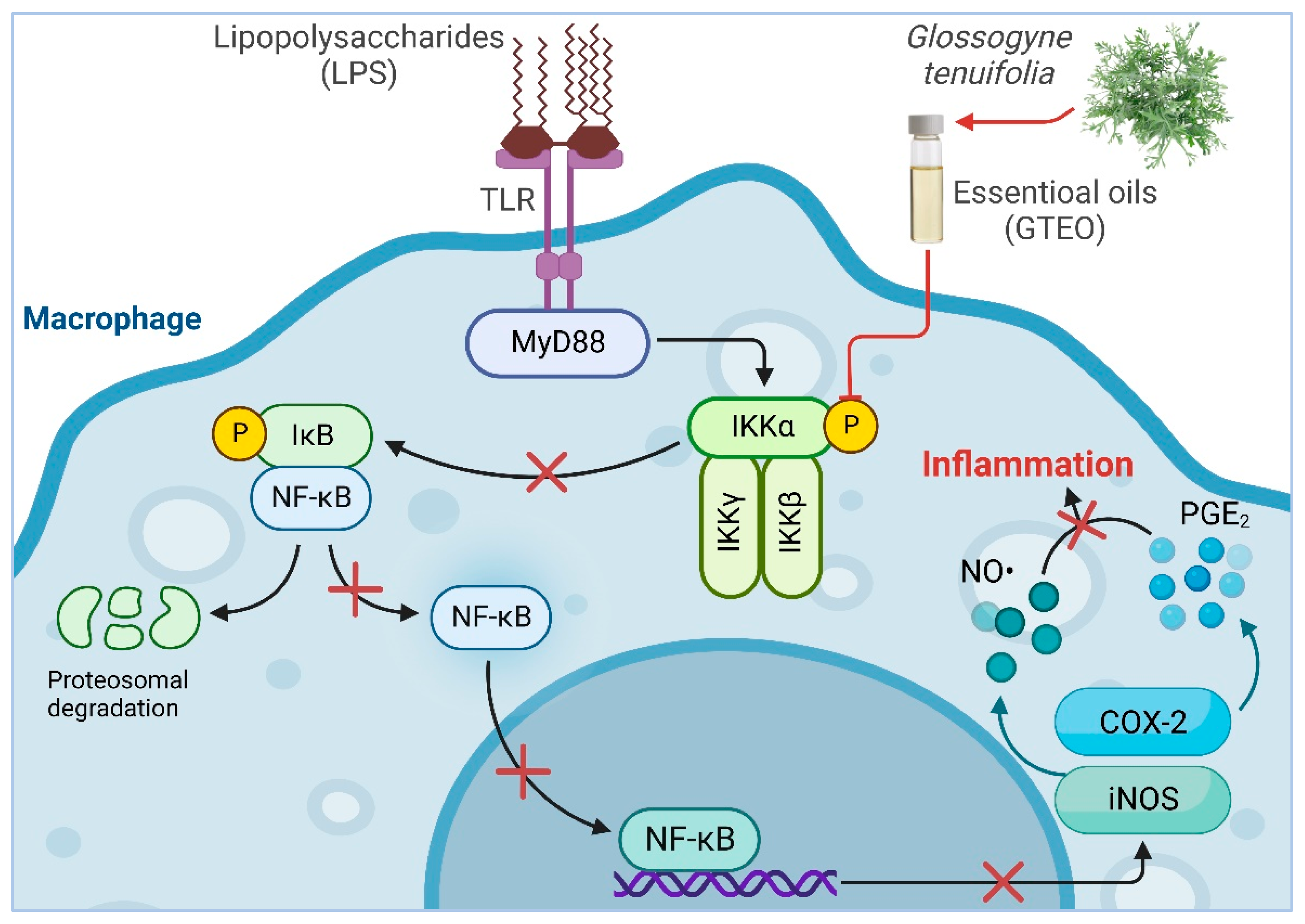
2. Results
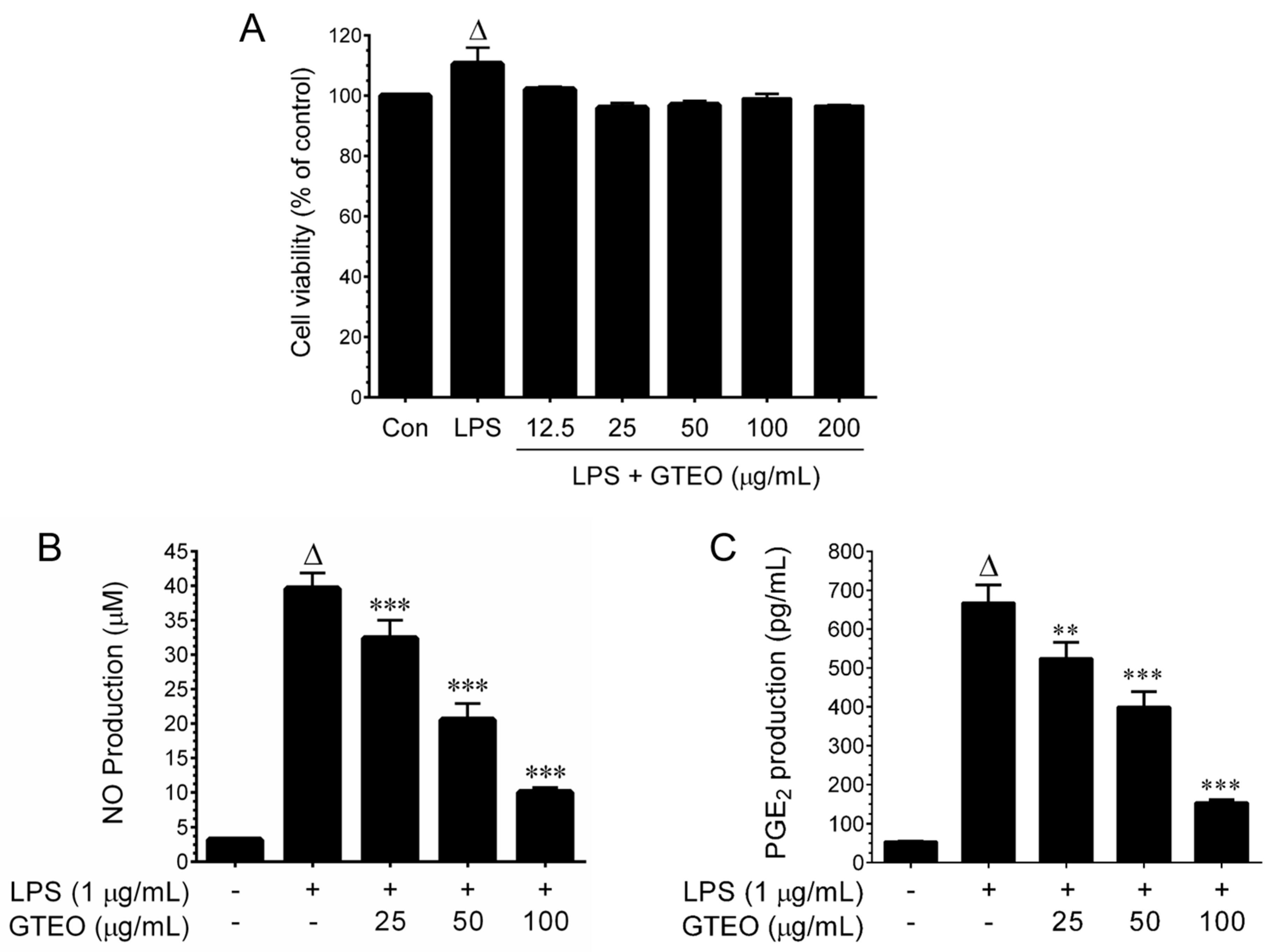
2.1. Effect of GTEO on Cell Viability in LPS-Activated RAW 264.7 Cells
2.2. GTEO Inhibits LPS-Induced NO and PGE2 Production in RAW 264.7 Cells
2.3. GTEO Inhibits LPS-Induced iNOS and COX-2 Expression in RAW 264.7 Cells
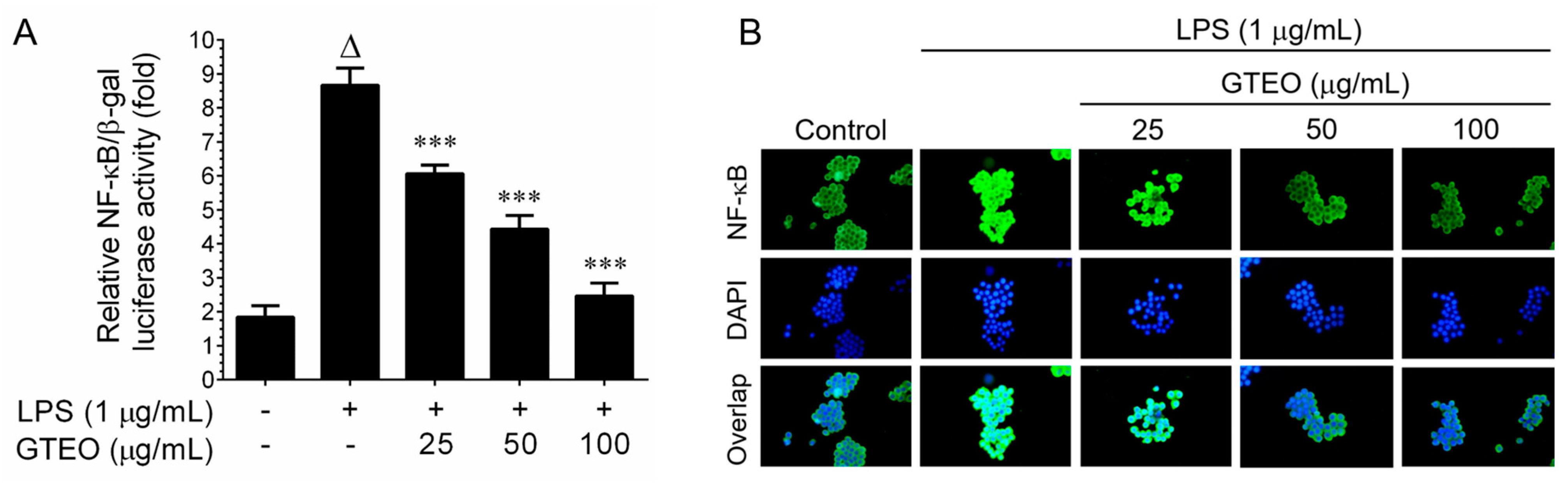
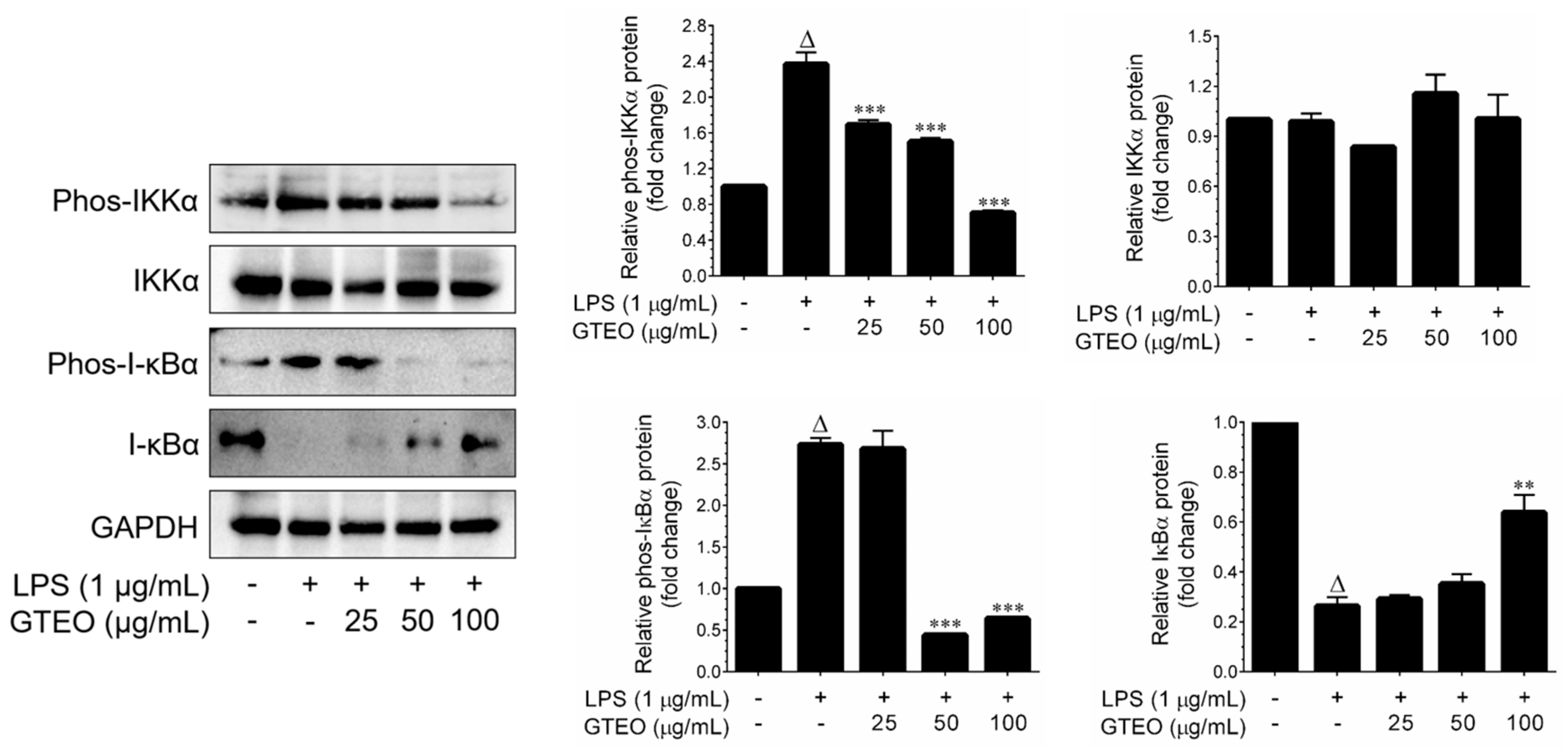
2.4. GTEO Suppressed LPS-Induced NF-κB Transcriptional Activity in RAW 264.7 Cells
2.5. Chemical Compositions of G. tenuifolia Essential Oil
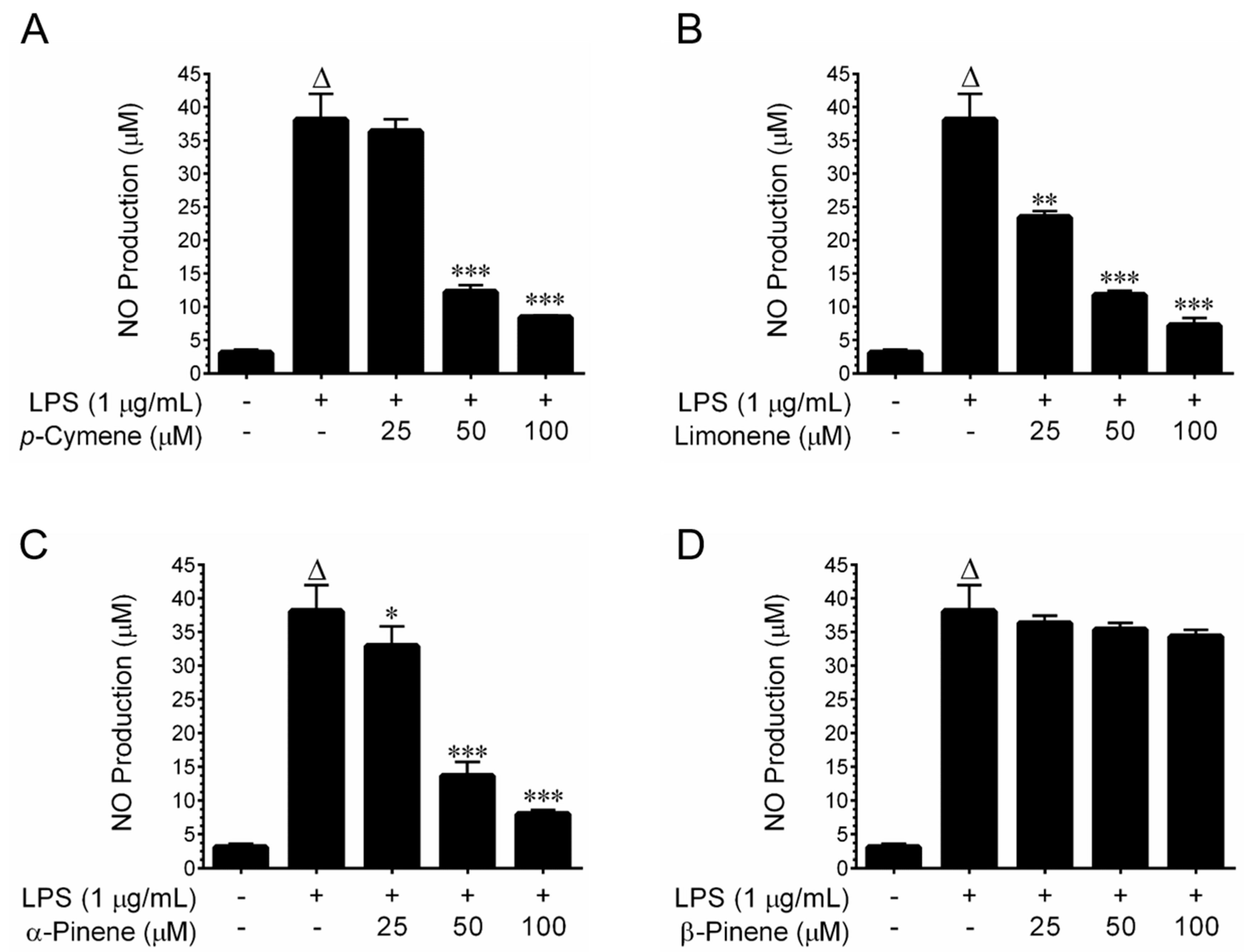
2.6. Nitric Oxide Inhibitory Effects of Major Constituents of GTEO
3. Discussion
4. Materials and Methods
4.1. Preparation of G. tenuifolia Essential Oil
4.2. Chemicals and Reagents
4.3. Cell Culture and Cell Viability Assay
4.4. Determination of NO and PGE2
4.5. RNA Extraction and Q-PCR Analyses
4.6. Protein Extraction and Immunoblotting
4.7. Immunofluorescence Assay
4.8. Luciferase Reporter Assay
4.9. GC-MS Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, M.J.; Kell, D.B.; Pretorius, E. The role of lipopolysaccharide-induced cell signalling in chronic inflammation. Chronic Stress. 2022, 6, 24705470221076390. [Google Scholar] [CrossRef] [PubMed]
- Basheer, A.S.; Abas, F.; Othman, I.; Naidu, R. Role of inflammatory mediators, macrophages, and neutrophils in glioma maintenance and progression: Mechanistic understanding and potential therapeutic applications. Cancers 2021, 13, 4226. [Google Scholar] [CrossRef] [PubMed]
- Saad, N.Y.; Muller, C.D.; Lobstein, A. Major bioactivities and mechanism of action of essential oils and their components. Flavour Frag. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Shaaban, H.A.E.; El-Ghorab, A.H.; Shibamoto, T. Bioactivity of essential oils and their volatile aroma components: Review. J. Essent. Oil Res. 2012, 24, 203–212. [Google Scholar] [CrossRef]
- Houng, J.Y.; Tai, T.S.; Hsu, S.C.; Hsu, H.F.; Hwang, T.S.; Lin, C.J.; Fang, L.W. Glossogyne tenuifolia (Hsiang-ju) extract suppresses T cell activation by inhibiting activation of c-Jun N-terminal kinase. Chin. Med. 2017, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Chang, H.C.; Kuo, C.L.; Agrawal, D.C.; Wu, C.R.; Tsay, H.S. In vitro propagation and analysis of secondary metabolites in Glossogyne tenuifolia (Hsiang-Ju)—A medicinal plant native to Taiwan. Bot. Stud. 2014, 55, 45. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.S.; Chao, L.K.; Liu, T.T. Antimicrobial activity of the essential oil of Glossogyne tenuifolia against selected pathogens. J. Sci. Food Agric. 2014, 94, 2965–2971. [Google Scholar] [CrossRef]
- Wu, M.J.; Wang, L.; Ding, H.Y.; Weng, C.Y.; Yen, J.H. Glossogyne tenuifolia acts to inhibit inflammatory mediator production in a macrophage cell line by downregulating LPS-induced NF-kappa B. J. Biomed. Sci. 2004, 11, 186–199. [Google Scholar] [CrossRef]
- Wu, M.J.; Weng, C.Y.; Ding, H.Y.; Wu, P.J. Anti-inflammatory and antiviral effects of Glossogyne tenuifolia. Life Sci. 2005, 76, 1135–1146. [Google Scholar] [CrossRef]
- Chen, Y.J.; Baskaran, R.; Shibu, M.A.; Lin, W.T. Anti-fatigue and exercise performance improvement effect of Glossogyne tenuifolia extract in mice. Nutrients 2022, 14, 1011. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.J.; Huang, C.L.; Lian, T.W.; Kou, M.C.; Wang, L. Antioxidant activity of Glossogyne tenuifolia. J. Agric. Food Chem. 2005, 53, 6305–6312. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Tsai, S.Y.; Han, C.M.; Shih, C.C.; Mau, J.L. Antioxidant properties of Glossogyne tenuifolia. Am. J. Chin. Med. 2006, 34, 707–720. [Google Scholar] [CrossRef]
- Hsuan, C.F.; Lu, Y.C.; Tsai, I.T.; Houng, J.Y.; Wang, S.W.; Chang, T.H.; Chen, Y.L.; Chang, C.C. Glossogyne tenuifolia attenuates proliferation and migration of vascular smooth muscle cells. Molecules. 2020, 25, 5832. [Google Scholar] [CrossRef]
- Hsu, H.F.; Houng, J.Y.; Kuo, C.F.; Tsao, N.; Wu, Y.C. Glossogin, a novel phenylpropanoid from Glossogyne tenuifolia, induced apoptosis in A549 lung cancer cells. Food Chem. Toxicol. 2008, 46, 3785–3791. [Google Scholar] [CrossRef]
- Asokan, S.M.; Wang, R.Y.; Hung, T.H.; Lin, W.T. Hepato-protective effects of Glossogyne tenuifolia in Streptozotocin-nicotinamide-induced diabetic rats on high fat diet. BMC Complement. Altern. Med. 2019, 19, 117. [Google Scholar] [CrossRef] [Green Version]
- Tien, Y.H.; Chen, B.H.; Wang Hsu, G.S.; Lin, W.T.; Huang, J.H.; Lu, Y.F. Hepatoprotective and anti-oxidant activities of Glossogyne tenuifolia against acetaminophen-induced hepatotoxicity in mice. Am. J. Chin. Med. 2014, 42, 1385–1398. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.L.; Weng, C.Y.; Wang, L.; Lian, T.W.; Wu, M.J. Immunomodulatory effect of Glossogyne tenuifolia in murine peritoneal macrophages and splenocytes. J. Ethnopharmacol. 2006, 107, 116–125. [Google Scholar] [CrossRef]
- Chyau, C.C.; Tsai, S.Y.; Yang, J.H.; Weng, C.C.; Han, C.M.; Shih, C.C.; Mau, J.L. The essential oil of Glossogyne tenuifolia. Food Chem. 2007, 100, 808–812. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Targeted. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.C. The noncanonical NF-κB pathway. Immunol. Rev. 2012, 246, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Hsuan, C.F.; Hsu, H.F.; Tseng, W.K.; Lee, T.L.; Wei, Y.F.; Hsu, K.L.; Wu, C.C.; Houng, J.Y. Glossogyne tenuifolia Extract inhibits TNF-α-induced expression of adhesion molecules in human umbilical vein endothelial cells via blocking the NF-κB signaling pathway. Molecules 2015, 20, 16908–16923. [Google Scholar] [CrossRef] [Green Version]
- Hsuan, C.F.; Lee, T.L.; Tseng, W.K.; Wu, C.C.; Chang, C.C.; Ko, T.L.; Chen, Y.L.; Houng, J.Y. Glossogyne tenuifolia extract increases nitric oxide production in human umbilical vein endothelial cells. Pharmaceuticals 2021, 14, 577. [Google Scholar] [CrossRef] [PubMed]
- Teng, O.; Ang, C.K.E.; Guan, X.L. Macrophage–Bacteria Interactions—A Lipid-Centric Relationship. Front. Immunol. 2017, 8, 1836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Joo, T.; Sowndhararajan, K.; Hong, S.; Lee, J.; Park, S.Y.; Kim, S.; Jhoo, J.W. Inhibition of nitric oxide production in LPS-stimulated RAW 264.7 cells by stem bark of Ulmus pumila L. Saudi. J. Biol. Sci. 2014, 21, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, M.; Rosenberg, D.W. Multifaceted roles of PGE2 in inflammation and cancer. Semin. Immunopathol. 2013, 35, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ohigashi, H. Targeting NOX, iNOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Lee, H.J.; Jeon, Y.D.; Han, Y.H.; Kee, J.Y.; Kim, H.J.; Shin, H.J.; Kang, J.; Lee, B.S.; Kim, S.H.; et al. Alpha-pinene exhibits anti-inflammatory activity through the suppression of MAPKs and the NF-κB pathway in mouse peritoneal macrophages. Am. J. Chin. Med. 2015, 43, 731–742. [Google Scholar] [CrossRef]
- Santos, E.S.; Abrantes Coelho, G.L.; Saraiva Fontes Loula, Y.K.; Saraiva Landim, B.L.; Fernandes Lima, C.N.; Tavares de Sousa Machado, S.; Pereira Lopes, M.J.; Soares Gomes, A.D.; Martins da Costa, J.G.; Alencar de Menezes, I.R.; et al. Hypoglycemic, Hypolipidemic, and Anti-Inflammatory effects of beta-pinene in diabetic rats. Evid.-Based Complement Alternat. Med. 2022, 2022, 8173307. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.J.; Lee, N.H.; Hyun, C.G. Limonene suppresses lipopolysaccharide-induced production of nitric oxide, prostaglandin E2, and pro-inflammatory cytokines in RAW 264.7 macrophages. J. Oleo Sci. 2010, 59, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Santana, M.F.; Guimarães, A.G.; Chaves, D.O.; Silva, J.C.; Bonjardim, L.R.; Lucca Júnior, W.d.; Ferro, J.N.d.S.; Barreto, E.d.O.; Santos, F.E.d.; Soares, M.B.P.; et al. The anti-hyperalgesic and anti-inflammatory profiles of p-cymene: Evidence for the involvement of opioid system and cytokines. Pharm. Biol. 2015, 53, 1583–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, P.H.; Brand, C.; Carson, C.F.; Riley, T.V.; Prager, R.H.; Finlay-Jones, J.J. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res. 2000, 49, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.J.; Li, J.; Vani, M.G.; Hsieh, Y.H.; Kuo, Y.H.; Wang, S.Y. Bornyl cinnamate inhibits inflammation-associated gene expression in macrophage cells through suppression of nuclear factor-κB signaling pathway. Planta Med. 2015, 81, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Senthil Kumar, K.J.; Gokila Vani, M.; Wang, C.S.; Chen, C.C.; Chen, Y.C.; Lu, L.P.; Huang, C.H.; Lai, C.S.; Wang, S.Y. Geranium and lemon essential oils and their active compounds downregulate angiotensin-converting enzyme 2 (ACE2), a SARS-CoV-2 spike receptor-binding domain, in epithelial cells. Plants 2020, 9, 770. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2017; ISBN 978-1-932633-21-4. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Compounds | Concentration (%) | KI a | Identification b |
|---|---|---|---|---|
| 10.29 | α-Pinene | 6.69 | 935 | KI/MS |
| 12 | β-Pinene | 3.16 | 975 | KI/MS |
| 12.19 | β-Myrcene | 14.68 | 979 | KI/MS |
| 14.36 | p-Cymene | 35.3 | 1026 | KI/MS |
| 14.56 | D-Limonene | 5.17 | 1031 | KI/MS |
| 14.63 | cis-β-Ocimene | 8.49 | 1032 | KI/MS |
| 15.93 | trans-Sabinene hydrate | 0.31 | 1060 | KI/MS |
| 17.95 | Linalool | 0.82 | 1100 | KI/MS |
| 21.74 | Terpinen-4-ol | 1.55 | 1182 | KI/MS |
| 22.03 | 2-Cyclohexen-1-one, 4-(1-methylethyl) | 1.14 | 1187 | KI/MS |
| 22.44 | α-Terpineol | 0.89 | 1195 | KI/MS |
| 24.61 | Benzaldehyde, 4-(1-methylethyl)- | 1.05 | 1244 | KI/MS |
| 26.21 | Phellandral | 2.9 | 1278 | KI/MS |
| 27.01 | Sabinyl acetate | 0.2 | 1295 | KI/MS |
| 27.19 | Carvacrol | 0.38 | 1298 | KI/MS |
| 28.24 | Myrtenyl acetate | 0.55 | 1323 | KI/MS |
| 30.66 | (E)-β-Damascenone | 0.26 | 1378 | KI/MS |
| 32.3 | β-Cedrene | 9.8 | 1416 | KI/MS |
| 35.05 | Germacrene D | 1.02 | 1483 | KI/MS |
| 38.16 | Nerolidol | 0.36 | 1561 | KI/MS |
| 38.93 | Caryophyllene oxide | 1.23 | 1580 | KI/MS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-T.; He, Y.-H.; Lo, Y.-H.; Chiang, Y.-T.; Wang, S.-Y.; Bezirganoglu, I.; Kumar, K.J.S. Essential Oil from Glossogyne tenuifolia Inhibits Lipopolysaccharide-Induced Inflammation-Associated Genes in Macro-Phage Cells via Suppression of NF-κB Signaling Pathway. Plants 2023, 12, 1241. https://doi.org/10.3390/plants12061241
Lin W-T, He Y-H, Lo Y-H, Chiang Y-T, Wang S-Y, Bezirganoglu I, Kumar KJS. Essential Oil from Glossogyne tenuifolia Inhibits Lipopolysaccharide-Induced Inflammation-Associated Genes in Macro-Phage Cells via Suppression of NF-κB Signaling Pathway. Plants. 2023; 12(6):1241. https://doi.org/10.3390/plants12061241
Chicago/Turabian StyleLin, Wan-Teng, Yen-Hua He, Yun-Hsin Lo, Yu-Ting Chiang, Sheng-Yang Wang, Ismail Bezirganoglu, and K. J. Senthil Kumar. 2023. "Essential Oil from Glossogyne tenuifolia Inhibits Lipopolysaccharide-Induced Inflammation-Associated Genes in Macro-Phage Cells via Suppression of NF-κB Signaling Pathway" Plants 12, no. 6: 1241. https://doi.org/10.3390/plants12061241