
Separate Effects of Sodium on Germination in Saline–Sodic and Alkaline Forms at Different Concentrations

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Solutions
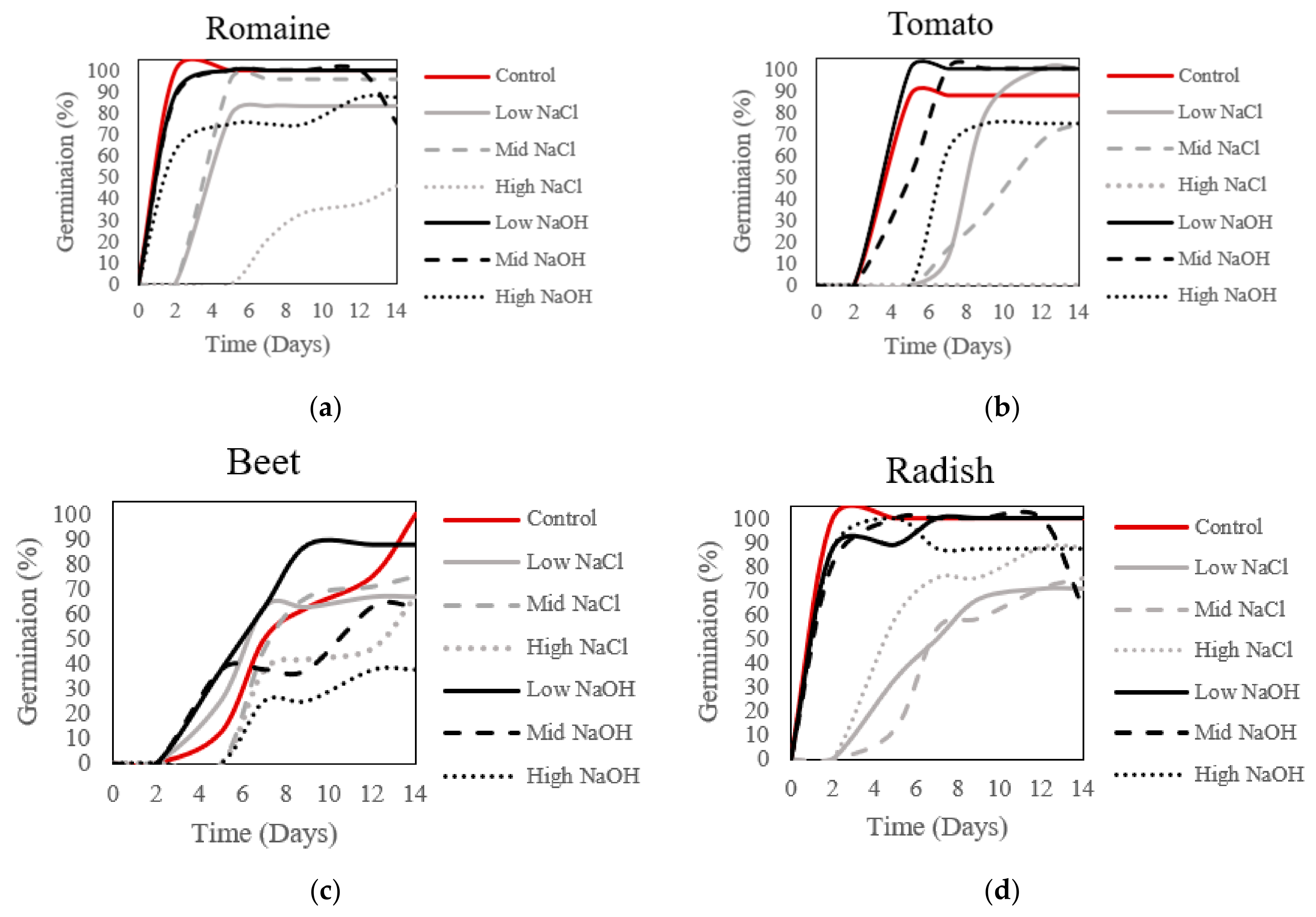
2.2. Germination and Plant Viability
2.3. pH
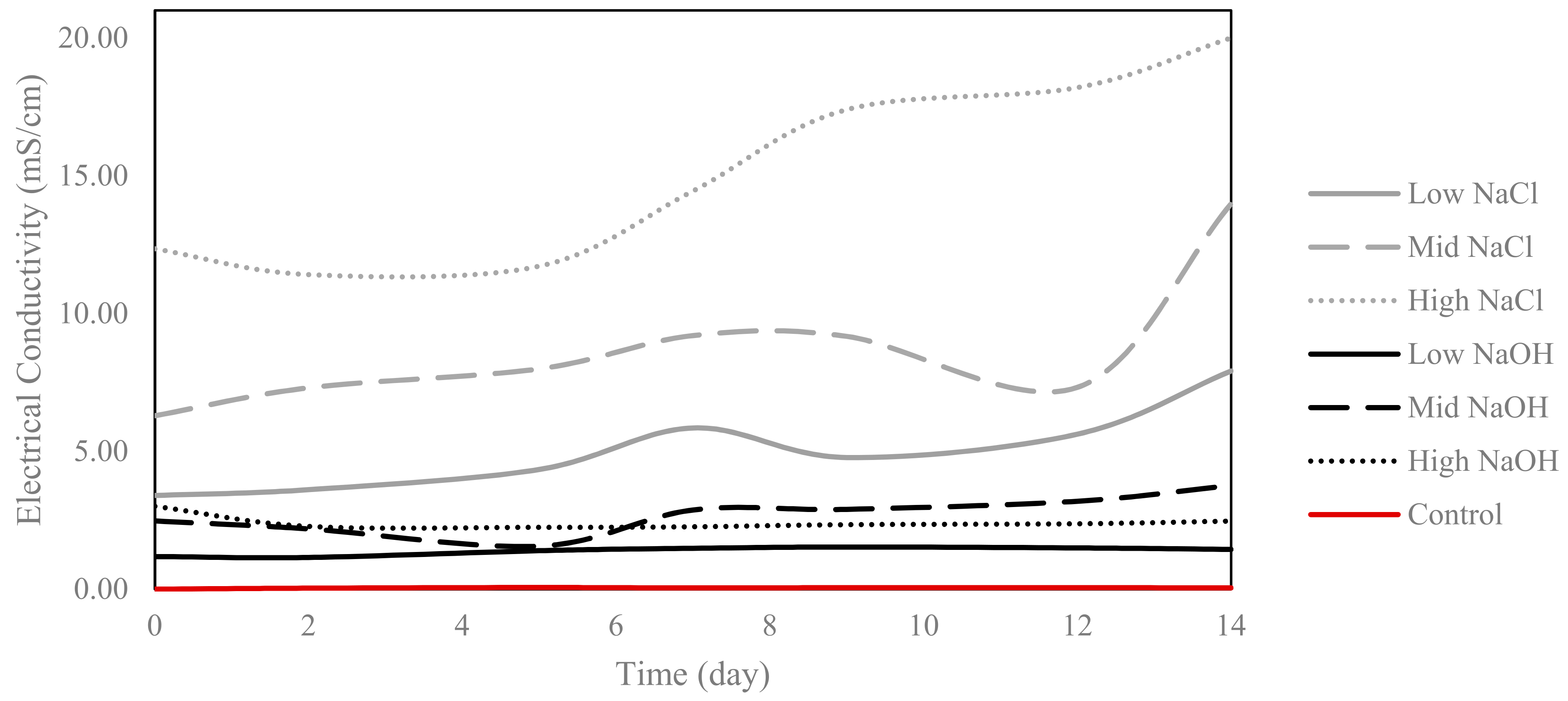
2.4. Electrical Conductivity (EC)
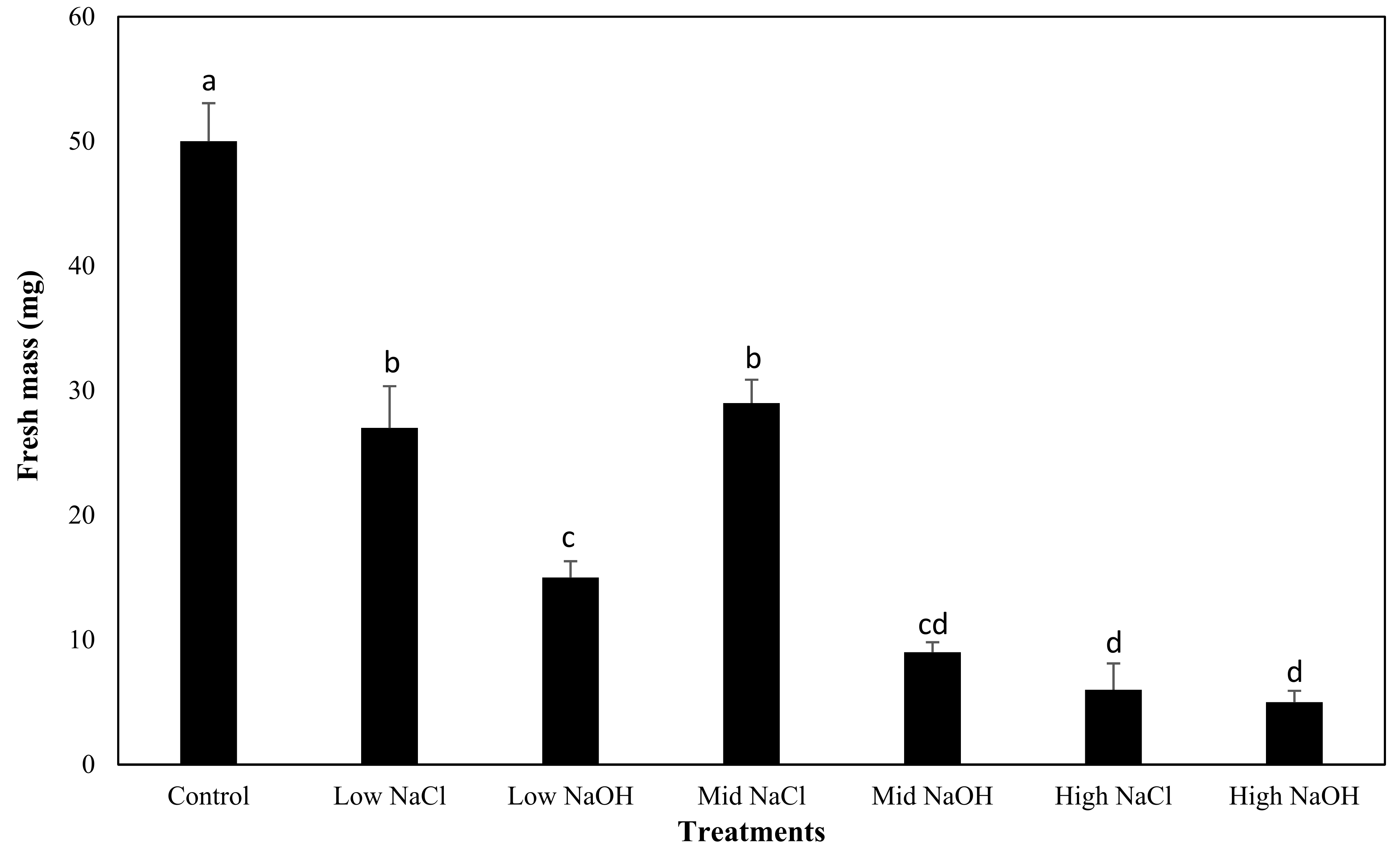
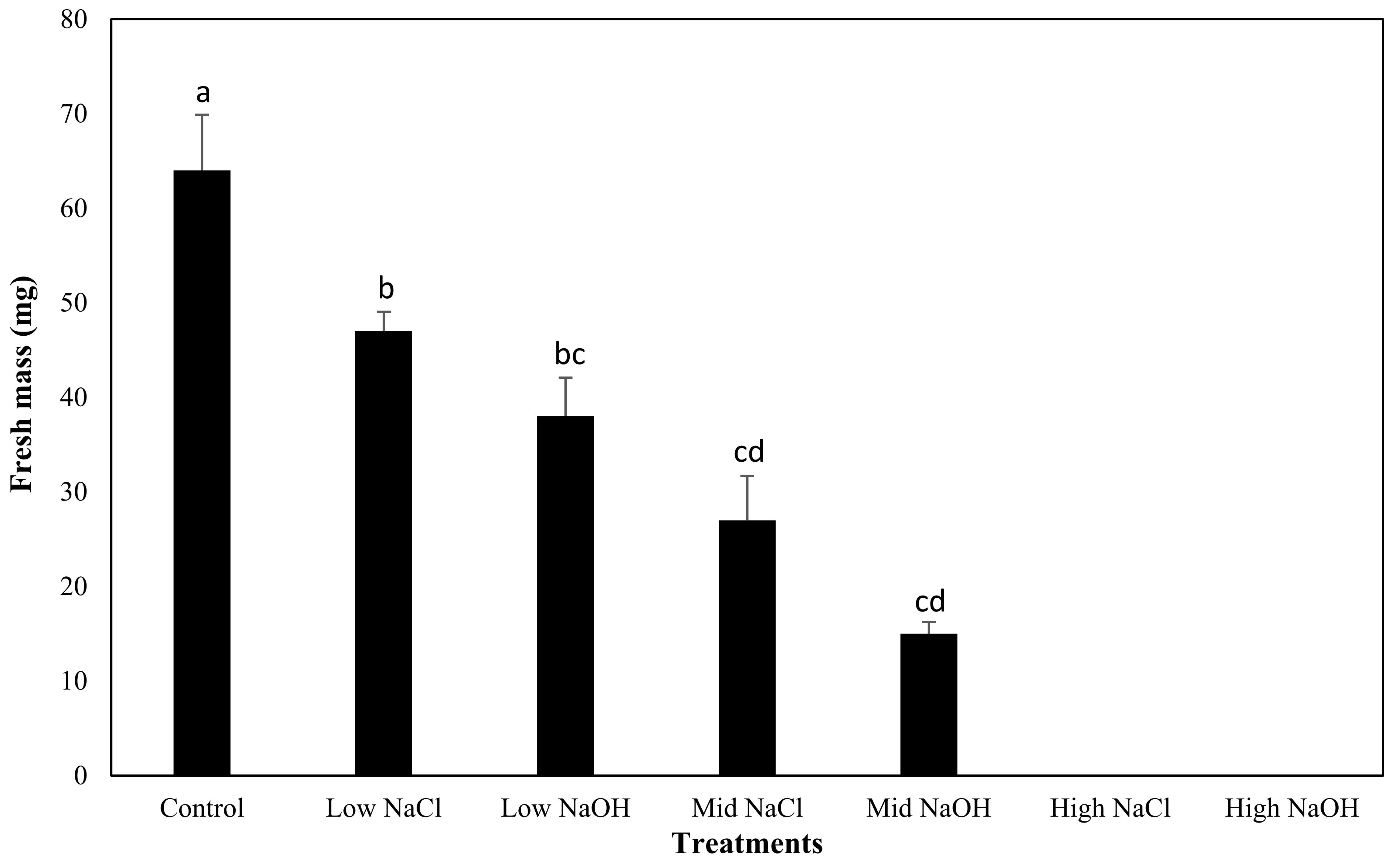
2.5. Fresh Mass Yield
2.6. Statistical Analysis
3. Results
3.1. EC
3.2. pH
3.3. Germination and Plant Viability
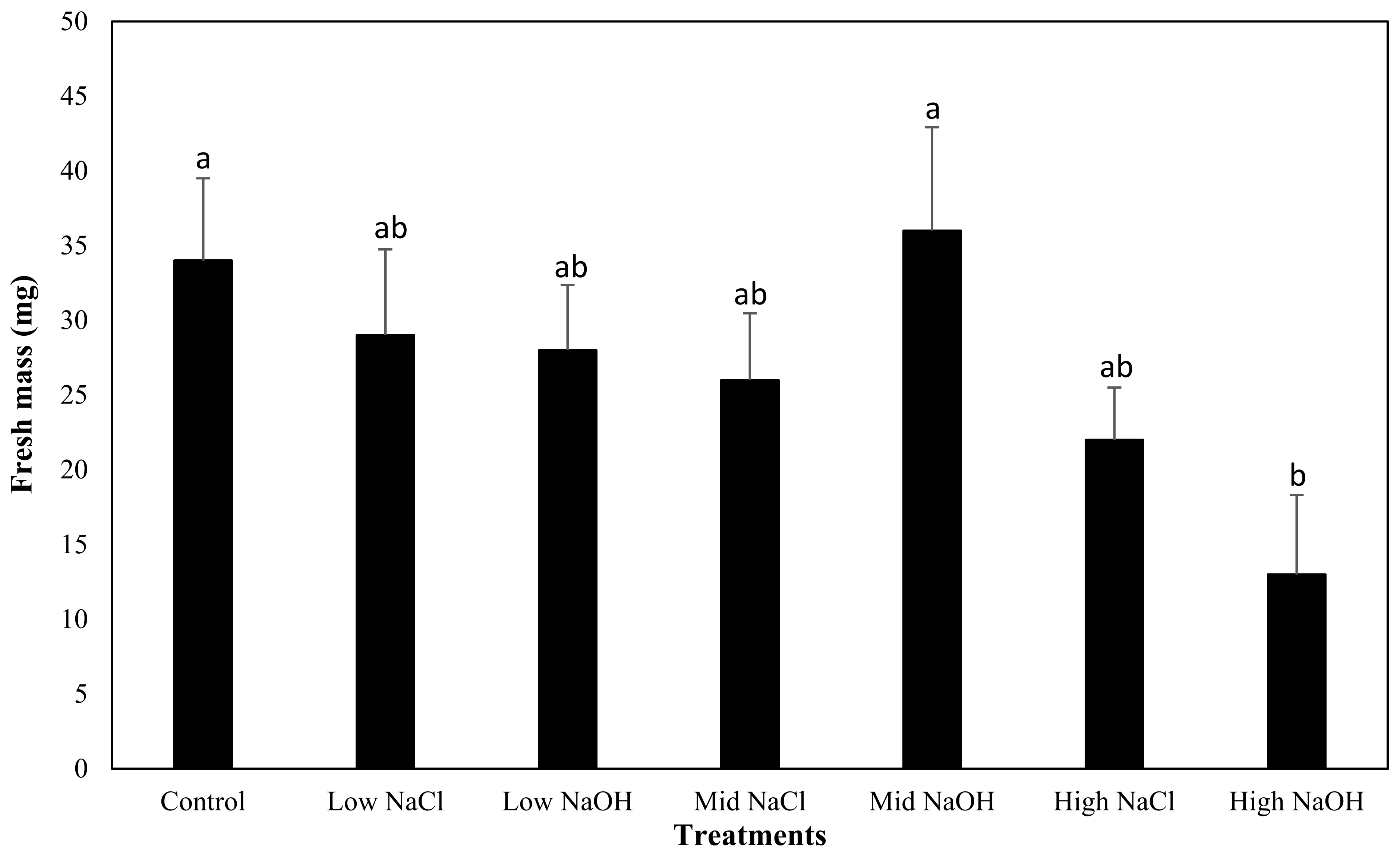
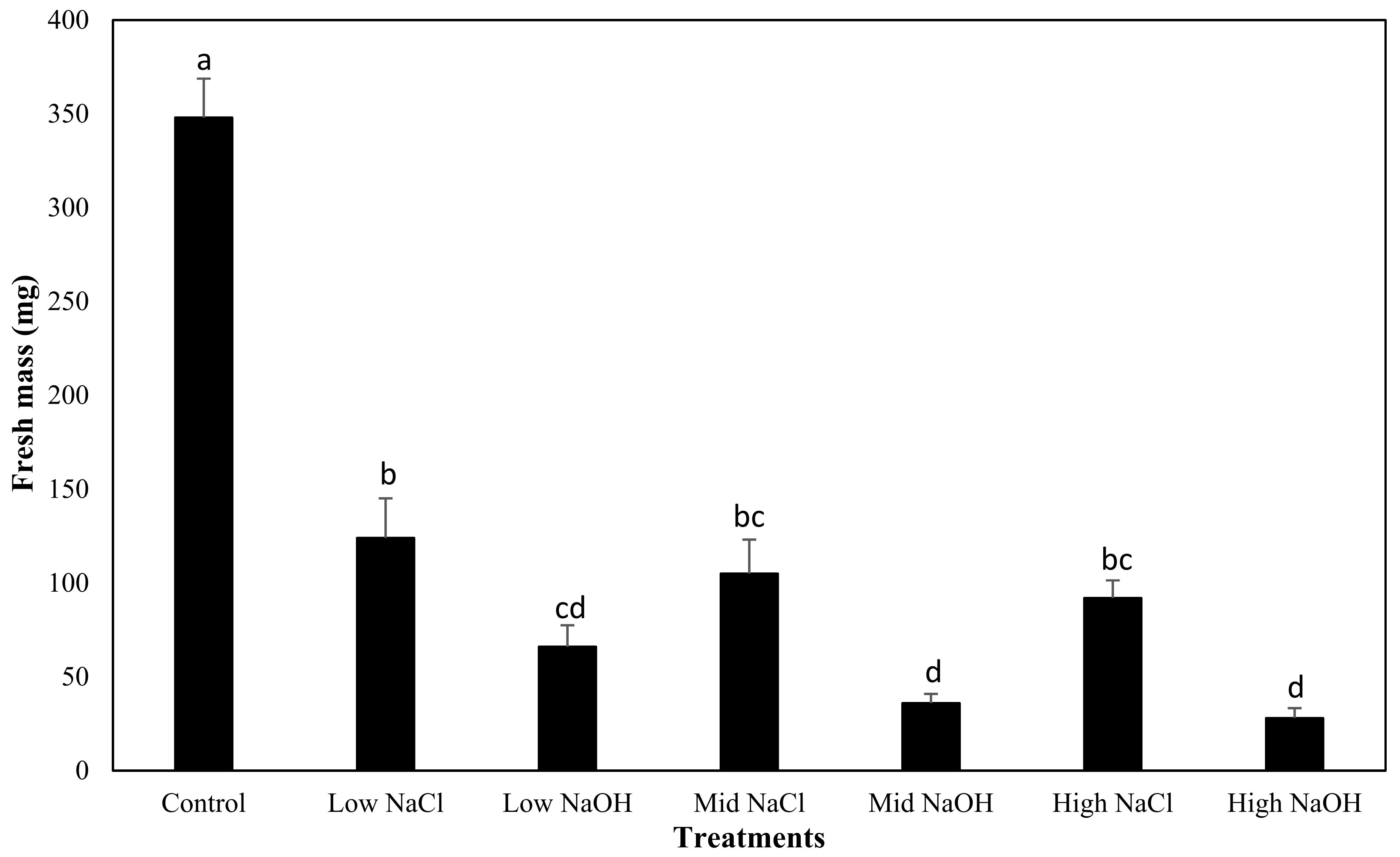
3.4. Fresh Mass Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, S. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667. [Google Scholar] [PubMed]
- Liu, J.; Guo, W.Q.; Shi, D.C. Seed germination, seedling survival, and physiological response of sunflowers under saline and alkaline conditions. Photosynthetica 2010, 48, 278–286. [Google Scholar] [CrossRef]
- Bernstein, L. Effects of salinity and sodicity on plant growth. Annu. Rev. Phytopathol. 1975, 13, 295–312. [Google Scholar] [CrossRef]
- Hopmans, J.W.; Qureshi, A.S.; Kisekka, I.; Munns, R.; Grattan, S.R.; Rengasamy, P.; Ben-Gal, A.; Assouline, S.; Javaux, M.; Minhas, P.S.; et al. Critical knowledge gaps and research priorities in global soil salinity. Adv. Agron. 2021, 169, 1–191. [Google Scholar]
- Shi, D.; Sheng, Y. Effect of various salt–alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors. Environ. Exp. Bot. 2005, 54, 8–21. [Google Scholar] [CrossRef]
- Li, R.; Shi, F.; Fukuda, K.; Yang, Y. Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (Medicago sativa L.). Soil Sci. Plant Nutr. 2010, 56, 725–733. [Google Scholar] [CrossRef]
- Javid, M.; Ford, R.; Nicolas, M.E. Tolerance responses of Brassica juncea to salinity, alkalinity and alkaline salinity. Funct. Plant Biol. 2012, 39, 699–707. [Google Scholar] [CrossRef]
- Läuchli, A.; Epstein, E. Plant responses to saline and sodic conditions. Agric. Salin. Assess. Manag. 1990, 71, 113–137. [Google Scholar]
- Chapman, H.D. Cation-exchange capacity. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; American Society of Agronomy, Inc.: Washington, DC, USA, 1965; Volume 9, pp. 891–901. [Google Scholar]
- Zhang, J.T.; Mu, C.S. Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius. Soil Sci. Plant Nutr. 2009, 55, 685–697. [Google Scholar] [CrossRef]
- Roosta, H.R. Interaction between water alkalinity and nutrient solution pH on the vegetative growth, chlorophyll fluorescence and leaf magnesium, iron, manganese, and zinc concentrations in lettuce. J. Plant Nutr. 2011, 34, 717–731. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Islam, A.K.M.S.; Edwards, D.G.; Asher, C.J. pH optima for crop growth: Results of a flowing solution culture experiment with six species. Plant Soil 1980, 54, 339–357. [Google Scholar] [CrossRef]
- Radi, A.A.; Abdel-Wahab, D.A.; Hamada, A.M. Evaluation of some bean lines tolerance to alkaline soil. J. Biol. Earth Sci. 2012, 2, 18–27. [Google Scholar]
- Farghaly, F.A.; Radi, A.A.; Abdel-Wahab, D.A.; Hamada, A.M. Effect of salinity and sodicity stresses on physiological response and productivity in Helianthus annuus. Acta Biol. Hung. 2016, 67, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Savvas, D.; Adamidis, K. Automated management of nutrient solutions based on target electrical conductivity, pH, and nutrient concentration ratios. J. Plant Nutr. 1999, 22, 1415–1432. [Google Scholar] [CrossRef]
- Walton, N.R.G. Electrical conductivity and total dissolved solids—What is their precise relationship? Desalination 1989, 72, 275–292. [Google Scholar] [CrossRef]
- Wilkins, C.L.; Trimpin, S. (Eds.) Ion Mobility Spectrometry-Mass Spectrometry: Theory and Applications; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Chrétien, S.; Gosselin, A.; Dorais, M. High electrical conductivity and radiation-based water management improve fruit quality of greenhouse tomatoes grown in rockwool. HortScience 2000, 35, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Light, T.S.; Licht, S.; Bevilacqua, A.C.; Morash, K.R. The fundamental conductivity and resistivity of water. Electrochem. Solid State Lett. 2004, 8, E16. [Google Scholar] [CrossRef]
- Lea-Cox, J.D.; Stutte, G.W.; Berry, W.L.; Wheeler, R.M. Nutrient dynamics and pH/charge-balance relationships in hydroponic solutions. In Proceedings of the International Symposium on Growing Media and Hydroponics, Windsor, ON, Canada, 19–26 May 1997; Volume 481, pp. 241–250. [Google Scholar]
- Patil, S.T.; Kadam, U.S.; Mane, M.S.; Mahale, D.M.; Dekale, J.S. Hydroponic nutrient solution: A review. J. Pharmacogn. Phytochem. 2020, 9, 2095–2099. [Google Scholar]
- Bugbee, B. Nutrient management in recirculating hydroponic culture. In Proceedings of the South Pacific Soilless Culture Conference-SPSCC, Palmerston North, New Zealand, 1 February 2003; Volume 648, pp. 99–112. [Google Scholar]
- Qu, X.X.; Huang, Z.Y.; Baskin, J.M.; Baskin, C.C. Effect of temperature, light and salinity on seed germination and radicle growth of the geographically widespread halophyte shrub Halocnemum strobilaceum. Ann. Bot. 2008, 101, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.X.; Li, Z.L.; Shao, S.; Wang, Y.N.; Mu, C.S. Effects of various mixed salt-alkaline stress conditions on seed germination and early seedling growth of leymus chinensis from songnen grassland of China. Not. Bot. Horti Agrobot. Cluj-Naoca 2014, 42, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Atiyeh, R.M.; Subler, S.; Edwards, C.A.; Bachman, G.; Metzger, J.D.; Shuster, W. Effects of vermicomposts and composts on plant growth in horticultural container media and soil. Pedobiologia 2000, 44, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Lu, Z.; He, L. Effects of saline-alkaline stress on seed germination and seedling growth of Sorghum bicolor (L.) Moench. Appl. Biochem. Biotechnol. 2014, 173, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Nieman, R.H. Some effects of sodium chloride on growth, photosynthesis, and respiration of twelve crop plants. Bot. Gaz. 1962, 123, 279–285. [Google Scholar] [CrossRef]
- Yadav, S.P.; Bharadwaj, R.; Nayak, H.; Mahto, R.; Singh, R.K.; Prasad, S.K. Impact of salt stress on growth, productivity and physicochemical properties of plants: A Review. Int. J. Chem. Stud. 2019, 7, 1793–1798. [Google Scholar]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- Geng, G.; Lv, C.; Stevanato, P.; Li, R.; Liu, H.; Yu, L.; Wang, Y. Transcriptome analysis of salt-sensitive and tolerant genotypes reveals salt-tolerance metabolic pathways in sugar beet. Int. J. Mol. Sci. 2019, 20, 5910. [Google Scholar] [CrossRef] [Green Version]
- Greenway, H. Plant response to saline substrates VII. Growth and ion uptake throughout plant development in two varieties of Hordeum vulgare. Aust. J. Biol. Sci. 1965, 18, 763–780. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Chong, J.N.; Kim, C.M.; Li, C.Y.; Shi, D.C.; Wang, D.L. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant Soil 2007, 294, 263–276. [Google Scholar] [CrossRef]
- Lin, J.; Li, X.; Zhang, Z.; Yu, X.; Gao, Z.; Wang, Y.; Wang, J.; Li, Z.; Mu, C. Salinity-alkalinity tolerance in wheat: Seed germination, early seedling growth, ion relations and solute accumulation. Afr. J. Agric. Res. 2012, 7, 467–474. [Google Scholar]
- Arnon, D.I.; Johnson, C.M. Influence of hydrogen ion concentration on the growth of higher plants under controlled conditions. Plant Physiol. 1942, 17, 525. [Google Scholar] [CrossRef] [Green Version]
- Malekzadeh Shamsabad, M.R.; Roosta, H.R.; Esmaeilizadeh, M. Responses of seven strawberry cultivars to alkalinity stress under soilless culture system. J. Plant Nutr. 2021, 44, 166–180. [Google Scholar] [CrossRef]
- Resh, H.M. Hydroponic Food Production: A Definitive Guidebook for the Advanced Home Gardener and the Commercial Hydroponic Grower; CRC Press: Boca Raton, FL, USA, 2012; pp. 593–595. [Google Scholar]
- Merle, G.; Wessling, M.; Nijmeijer, K. Anion exchange membranes for alkaline fuel cells: A review. J. Membr. Sci. 2011, 377, 1–35. [Google Scholar] [CrossRef]
- Zhang, Q.; Shao, X.; Yi, J.; Liu, Y.; Zhang, J. An experimental study of electroreduction of CO2 to HCOOH on SnO2/C in presence of alkali metal cations (Li+, Na+, K+, Rb+ and Cs+) and anions (HCO3−, Cl−, Br− and I−). Chin. J. Chem. Eng. 2020, 28, 2549–2554. [Google Scholar] [CrossRef]
- Shi, D.; Yin, L. Difference between salt (NaCl) and alkaline (Na2CO3) stresses on Puccinellia tenuiflora (Griseb.) Scribn. et Merr. plants. J. Integr. Plant Biol. 1993, 35, 144–149. [Google Scholar]
- El-Samad, H.M.; Shaddad, M.A.K. Comparative effect of sodium carbonate, sodium sulphate, and sodium chloride on the growth and related metabolic activities of pea plants. J. Plant Nutr. 1996, 19, 717–728. [Google Scholar] [CrossRef]
- Campbell, S.A.; Nishio, J.N. Iron deficiency studies of sugar beet using an improved sodium bicarbonate-buffered hydroponic growth system. J. Plant Nutr. 2000, 23, 741–757. [Google Scholar] [CrossRef]
- Yang, C.; Shi, D.; Wang, D. Comparative effects of salt and alkali stresses on growth, osmotic adjustment and ionic balance of an alkali-resistant halophyte Suaeda glauca (Bge.). Plant Growth Regul. 2008, 56, 179–190. [Google Scholar] [CrossRef]
- Valdez-Aguilar, L.A.; Reed, D.W. Growth and nutrition of young bean plants under high alkalinity as affected by mixtures of ammonium, potassium, and sodium. J. Plant Nutr. 2010, 33, 1472–1488. [Google Scholar] [CrossRef]
- Sonneveld, C.; Voogt, W. Nutrient solutions for soilless cultures. In Plant Nutrition of Greenhouse Crops; Springer: Dordrecht, The Netherlands, 2009; pp. 257–275. [Google Scholar]
- Pulford, I.D.; Watson, C. Phytoremediation of heavy metal-contaminated land by trees—A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef]
- Velazquez-Gonzalez, R.S.; Garcia-Garcia, A.L.; Ventura-Zapata, E.; Barceinas-Sanchez, J.D.O.; Sosa-Savedra, J.C. A review on hydroponics and the technologies associated for medium-and small-scale operations. Agriculture 2022, 12, 646. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Buffer | Components | Manufacturer |
|---|---|---|
| pH 8 | KH2PO4, NaOH | Thermo Fisher Scientific (Portsmouth, NH, US) |
| pH 9 | H2O, KCl, H2BO3, NaOH, and preservative | Ricca Chemical Company (Arlington, TX, US) |
| Saline-Sodic | Alkaline | ||||||
|---|---|---|---|---|---|---|---|
| Solution | Control | Low-NaCl | Mid-NaCl | High-NaCl | Low-NaOH | Mid-NaOH | High-NaOH |
| NaCl [mM] | - | 12.32 | 24.47 | 49.11 | 12.32 | 24.47 | 49.11 |
| Buffer volume | - | - | - | - | ¼ pH 8 buffer | ½ pH 8 buffer | ½ pH 9 buffer |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hitti, Y.; MacPherson, S.; Lefsrud, M. Separate Effects of Sodium on Germination in Saline–Sodic and Alkaline Forms at Different Concentrations. Plants 2023, 12, 1234. https://doi.org/10.3390/plants12061234
Hitti Y, MacPherson S, Lefsrud M. Separate Effects of Sodium on Germination in Saline–Sodic and Alkaline Forms at Different Concentrations. Plants. 2023; 12(6):1234. https://doi.org/10.3390/plants12061234
Chicago/Turabian StyleHitti, Yasmeen, Sarah MacPherson, and Mark Lefsrud. 2023. "Separate Effects of Sodium on Germination in Saline–Sodic and Alkaline Forms at Different Concentrations" Plants 12, no. 6: 1234. https://doi.org/10.3390/plants12061234