
Four Invasive Plant Species in Southwest Saudi Arabia Have Variable Effects on Soil Dynamics
by
 , , and
, , and
Ahmed M. Abbas
1,2,*,
Wagdi S. Soliman
3,
Maryam M. Alomran
4,
Nahaa M. Alotaibi
4 and
Stephen J. Novak
5,* 1
Department of Biology, College of Science, King Khalid University, P.O. Box 9004, Abha 61413, Saudi Arabia
2
Department of Botany and Microbiology, Faculty of Science, South Valley University, Qena 83523, Egypt
3
Horticulture Department, Faculty of Agriculture and Natural Resources, Aswan University, Aswan 81528, Egypt
4
Department of Biology, College of Science, Princess Nourah bint Abdulrahman University, P.O. Box 84428, Riyadh 11671, Saudi Arabia
5
Department of Biological Sciences, Boise State University, Boise, ID 83725, USA
*
Authors to whom correspondence should be addressed.
Plants 2023, 12(6), 1231; https://doi.org/10.3390/plants12061231
Submission received: 31 January 2023
/
Revised: 4 March 2023
/
Accepted: 6 March 2023
/
Published: 8 March 2023
(This article belongs to the Topic Plant Invasion)
Abstract
:Predicting the direction and magnitude of change in soil dynamics caused by invasive plant species has proven to be difficult because these changes are often reported to be species- and habitat-specific. This study was conducted to determine changes in three soil properties, eight soil ions, and seven soil microelements under established stands of four invasive plants, Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica. Soil properties, ions, and microelements were measured in sites invaded by these four species in southwest Saudi Arabia, and these values were compared to the results for the same 18 parameters from adjacent sites with native vegetation. Because this study was conducted in an arid ecosystem, we predict that these four invasive plants will significantly alter the soil properties, ions, and microelements in the areas they invaded. While the soils of sites with the four invasive plant species generally had higher values for soil properties and ions compared to sites with native vegetation, in most instances these differences were not statistically significant. However, the soils within sites invaded by I. carnea, L. leucocephala, and P. juliflora had statistically significant differences for some soil parameters. For sites invaded by O. puntia ficus-indica, no soil properties, ions, or microelements were significantly different compared to adjacent sites with native vegetation. Sites invaded by the four plant species generally exhibited differences in the 11 soil properties, but in no instance were these differences statistically significant. All three soil properties and one soil ion (Ca) were significantly different across the four stands of native vegetation. For the seven soil microelements, significantly different values were detected for Co and Ni, but only among stands of the four invasive plant species. These results indicate that the four invasive plant species altered soil properties, ions, and microelements, but for most of the parameters we assessed, not significantly. Our results do not support our initial prediction, but are in general agreement with previous published findings, which indicate that the effects of invasive plants on soil dynamics vary idiosyncratically among invasive species and among invaded habitats.
1. Introduction
In regions where native plants and animals did not meet the needs of early colonists, these colonists began moving plants and animals around the globe, with some of these non-native species becoming invasive [1,2]. Invasive plant species are often described as having negative impacts on ecosystem processes [3]. They not only pose a threat to global biodiversity hotspots, but also to all the habitats (i.e., forest, grasslands, etc.) they invade. There are many ways that invasive species damage native communities and ecosystems, especially in remote locations and on islands, where native species may be more vulnerable to these non-natives [4,5]. The extent to which invasive plants alter ecosystem processes in their new range compared to the ecosystem processes of native species in these areas has been shown to vary among invasive species and invaded habitats [6,7,8]. Many accounts of the impacts of invasive species are available for certain habitats and regions around the world [9,10], but details on the impacts of invasive species in other habitats and regions (e.g., in Saudi Arabia) are largely unknown, but see Assaeed et al. [11].
Invasive plant species can cause rapid changes in soil properties relative to native communities, and these differences may hinder efforts to restore native plant communities [8,12]. Invasive plants modify soil conditions either directly, by depositing leaf litter of varying quality and quantity [13,14], or indirectly, by altering the native microbial communities and the biotic interactions they normally engage in [14,15]. Invasive plants may alter soil properties in ways that increase their competitive advantage. For example, the soils of sites infested with invasive plants have lower levels of accessible phosphorus, total nitrogen, and carbon than sites with native vegetation [16,17]. However, the soil carbon/nitrogen ratio of soils under invasive plants has been reported to increase [8]. Additionally, the capacity of soils in invaded wetlands to store and release carbon was influenced by plant biomass input in soils, the sediment deposition rate of the tidal salt marsh, carbon turnover, and organic carbon stability, which are generally reported to be associated with the accumulation and decomposition of organic matter [18]. Invasive species in tropical dry forests negatively alter soil characteristics [19]. In contrast, the presence of invasive species in degraded habitats can improve soil moisture conditions and can facilitate the reclamation of such degraded areas [20].
Invasive species that displace native plants create unique plant communities that alter or create new plant–soil nutrient cycles and carbon cycles and change the capacity of soil to accumulate N and P [21]. In fact, the ability of invasive plants to alter the concentration and stoichiometry of soil nutrients has been widely studied [22,23,24,25]. Some invasive plants may alter the soil properties (and other ecological processes) to such an extent that the ecosystem services associated with native communities are threatened or are no longer provided [19,26,27,28]. Such invasive plant species are referred to as “transformer species” [29]. However, recent assessments of invasive plant species highlight the need to distinguish between non-native (invasive) species that have minimal ecological consequences and non-native species that have major ecological consequences in their new habitats [30,31]. There is ample evidence that invasive species can alter ecological processes and reduce native biodiversity, yet the ecological consequences of invasive tree species and how they modify soil properties and conditions are still not well understood and not adequately documented [32].
Other studies have reported no statistically significant difference among the properties and microelements of soils from habitats infested with invasive plant species compared to the soils of equivalent habitats occupied by native plant species [6,8,12,32,33]. Moreover, several studies that have compared the capacity of multiple invasive species to alter soil properties within the same experiments design have revealed contrasting results among the species examined [6,8,32]. Other studies have reported that the same invasive species alters soil dynamics in different ways in different habitats (communities) or under different experimental conditions [6,8,12]. These results indicate that the effects of invasive plants on soil dynamics vary in an idiosyncratic manner among invasive species and among invaded habitats (see [6,8]). Thus, the alteration of soil properties by invasive plants is often species- and habitat--specific.
Invasive plant species in Saudi Arabia have negative ecological consequences and high economic costs in natural ecosystems and agriculture areas, respectively [34]. These invasive plants have displaced many native plants from species-rich areas, especially in high-elevation habitats, wadis, and meadows. Some non-native plant species are believed to have been introduced into Saudi Arabia during past Hajj seasons, while others were introduced by early settlers for use as food, medicine, ornamental uses, livestock consumption, shade, and other reasons [34]. The introduction of non-native plants continues and is even increasing due to increased international trade, commerce, travel, and accidental means [35]. In Saudi Arabia, the establishment of non-native plant species is also facilitated by human disturbance of natural habitats, including the removal of native plants for road-building activities. This process explains the presence of invasive species along roads in mountainous regions of southwest Saudi Arabia [34]. Additionally, many other regions of southwest Saudi Arabia, plant diversity hotspots such as wadis and Red Sea islands, now contain many invasive plant species [34]. Unfortunately, scientific research focused on the invasive plant species in this region of Saudi Arabia is lacking (but see Assaeed et al., [11] and Alfarhan et al., [36]). Thus, there is an urgent need to conduct research on the invasive plants in southwest Saudi Arabia and to better understand their ecological consequences.
Six non-native plant species have been identified as the most important invasive species in Saudi Arabia [34]: Prosopis juliflora, Cuscuta campestris, Cenchrus echinatus, Verbesina encelioides, Leucaena leucocephala, and Prosopis koelziana. Among these six species, P. juliflora is the most damaging invasive plant species. It was reported as a ruderal plant, mostly found in urban and suburban areas, and is rarely observed in native plant communities of the central region of Saudi Arabia [34]. Unlike other tropical and subtropical countries, the invasion of L. leucocephala in Saudi Arabia does not have severe impacts on native communities because it mostly occupies urban areas. Opuntia ficus-indica and Opuntia dillenii are widely distributed across Saudi Arabia. Ipomoea carnea ssp. fistulosa, a pantropical invasive plant, has wide ecological amplitude. According to Al-Sodany [37], the growth of I. carnea follows a seasonal pattern in which its growth is greatest from September and October. This study reported significant differences for growth parameters (e.g., height, crown diameter, crown volume, and size index), demographic characteristics (e.g., birth, mortality, and survival rates), and reproductive output (e.g., number of flowers and fruits) among populations of I. carnea in different habitats [37]. I. carnea subsp. fistulosa reproduces clonally, through the production of underground rhizomes that grow laterally through the soil [38].
This research was designed to assess the direction and magnitude of change in soil properties, soil ions, and soil microelements caused by the invasion of four invasive plant species, Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica, in southwest Saudi Arabia. This was achieved by sampling and analyzing soils in areas invaded by the four species and comparing these results to those obtained for soils from adjacent areas where native vegetation occurred. A meta-analysis by Waring et al. [39] showed that the ability of plants to alter soil dynamics is significantly stronger in arid ecosystems; therefore, we predict that all four invasive plants included in this study will significantly alter soil dynamics in the areas they have invaded. The results of this study will be used to test this prediction.
2. Results
2.1. Comparison between Invaded and Native Soil Samples
The soil properties, ion concentrations, and microelement concentrations of sites in southwest Saudi Arabia invaded by Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica were compared to the soils from sites with native vegetation. Most soil properties, ion concentrations, and soil microelements concentrations did not significantly differ between the P. juliflora stands and native stands, except for several microelements, including Cr, Co, Cu, and Ni (Table 1). The concentration of these four microelements was significantly higher for the soil under native vegetation compared to the soil of sites invaded by the tree P. juliflora. Except for pH, organic matter (OM), Ca, and SO4, no soil properties, ions, or microelements were significantly different for soils from stands invaded by the shrub I. carnea and soils from sites with native vegetation (Table 2).
The pH was significantly higher for the soil under native vegetation, while OM, CA, and SO4 were significantly higher for the soils from stands with an I. carnea overstory (Table 2). Soil properties, ion concentrations, and soil microelements did not significantly differ between stands in which the tree L. leucocephala had invaded and stands with native vegetation, except for pH, which was significantly higher in soil from areas with native vegetation (Table 3). No soil properties, ion concentrations, or soil microelements were significantly different for soils from stands invaded by the cactus O. ficus-indica and stands of native vegetation (Table 4).
2.2. Soil Properties, Ion Concentrations, and Microelement Concentrations for the Four Invasive Plant Species and Adjacent Stands of Native Vegetation
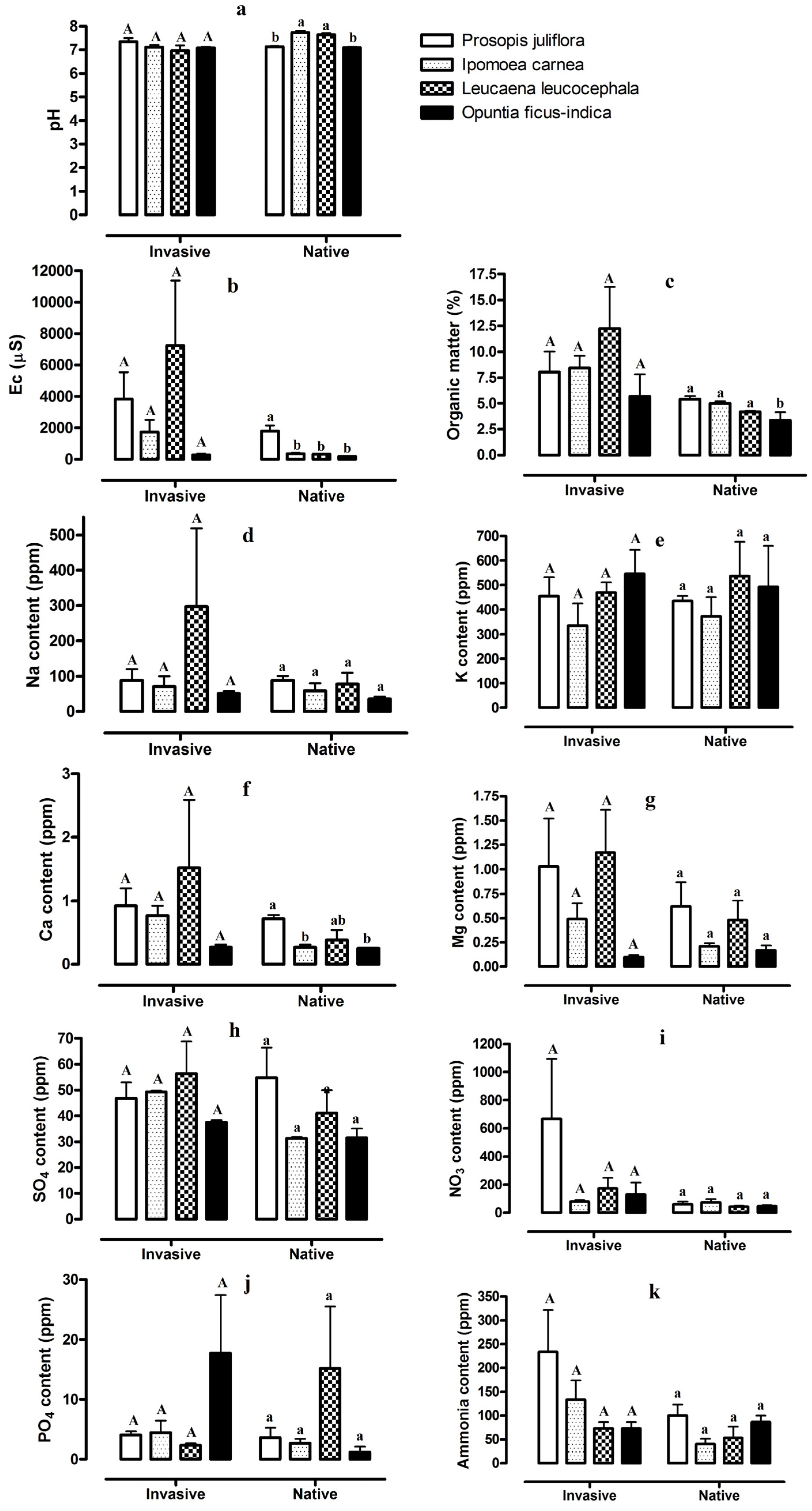
There were large differences in soil properties and ion concentrations in the soils from stands with native vegetation and the soil of stands invaded by the four invasive plant species. We detected significant differences in soil pH, EC, OM, and Ca among the four stands of native vegetation, but not for the stands of the four invasive species (Figure 1). In soils from the native sites, pH ranged from 7.1 to 7.7, and in the invaded site pH ranged from 6.95 to 7.35. pH values were higher in the areas invaded by P. juliflora and O. ficus-indica (compared to adjacent native sites) and lower in the areas invaded by I. carnea and L. leucocephala (compared to adjacent native sites). Conversely, EC and OM values were higher in the soils of invaded stands compared to the soil of native sites. The largest increase in the values of EC and OM occurred for sites invaded by L. leucocephala (Figure 1). Similarly, the values of Na, Ca, and Mg were highest in the soils of stands that were invaded by all four invasive plants, and the largest values were observed in areas invaded by L. leucocephala (Figure 1). The concentrations of K in soils of invaded stands of all four invasive species and adjacent native stands were not significantly different (Figure 1). Variation among soil SO4 values for the four invasive species in both invaded and native stands were not significantly different (Figure 1). However, the NO3 and ammonia values were generally higher in the soils of stands invaded by the four species compared to the soils of sites with native vegetation. The largest increases in NO3 values were observed for areas invaded by P. juliflora, followed by areas invaded by L. leucocephala, with the largest increases in ammonia values occurring in areas invaded with I. carnea, followed by areas infested with P. juliflora (Figure 1). The soil PO4 content exhibited variable results (Figure 1). For instance, the soil PO4 content increased in the stands invaded by O. ficus-indica compared to the soil of native stands and decreased in areas invaded by P. juliflora (Figure 1).
There were also differences in the concentration of microelements in the soils from stands invaded by the four invasive plant species and adjacent stands of native vegetation (Figure 2). The concentration of Mn, Zn, and Fe showed higher values in areas invaded by O. ficus-indica and L. leucocephala, while these microelements had lower concentrations in areas invaded by P. juliflora and I. carnea. These differences, however, were not statistically significant. In addition, variation in Cu and Cr in the four stands of native vegetation and the four sites with invasive plants were not significantly different, but values for both microelements were lowest for the soil of sites invaded by P. juliflora (Figure 2). The concentrations of Co and Ni significantly differed among stands of the four invasive plant species and was significantly lower for the soils of sites invaded by P. juliflora, I. carnea, and L. leucocephala. The four stands of native vegetation exhibited differences for all seven microelements, but none of these differences were statistically significant (Figure 2).
3. Discussion
Invasive species are those species (taxa) that are introduced outside of their native range and persist, reproduce, and spread beyond their original points of introduction in their new range [40,41]. Invasive plant species can outcompete and reduce the abundance of native plant species [42], alter soil properties [26], and homogenize the biodiversity of invaded communities and ecosystems [43]. Invasive plant species may alter soil properties in their new ranges, but the magnitude and direction of these changes are sometimes unique to each invasive species and often vary among habitats for the same non-native species [6,8,12,32,33,44,45]. To the best of our knowledge, this is the first study to investigate the biogeochemical consequences of the invasion of Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica in Saudi Arabia. At the outset of this study we predicted that all four invasive plant species would significantly alter soil properties and microelements in the areas they invaded. Our results do not support this prediction.
Soil carbon, nitrogen, and phosphorus have a large influence on many ecosystem processes, and the direction of change for these three elements was widespread across the species and sites studied. Our study revealed higher levels for soil carbon, nitrogen, and phosphorus levels in soils of invaded habitats compared to the soils of sites with native vegetation. A recent meta-analysis discovered that invasive plant species were associated with significantly higher litter deposition and decomposition rates as well as increases in soil nitrogen mineralization and nitrification [46,47]. Ehrenfeld [48] and Ehrenfeld et al. [13] proposed that the characteristics of many invasive plant species, in comparison to native plants, may influence the specific way in which invasive plants alter ecosystem properties. These characteristics include larger size, faster growth, higher photosynthetic rates, higher nutrient concentrations in living tissue and litter, and extremely efficient nitrogen-fixing symbioses. Rout and Callaway [49] state that if invasive species increase net primary production and nitrogen cycling in their invasive ranges but not in their native ranges, their inherent traits are unlikely to drive these processes because these same ecosystem effects should also occur in their native ranges. This statement also highlights the context-specific nature of traits on invasion outcomes.
Another possibility is that invasive plants, once established, will undergo rapid evolution through natural selection [50,51], especially for key traits related to nutrient cycling. For example, Feng et al. [47] provided evidence that the invasive plant Ageratina adenophora, which is native to Mexico but has spread throughout subtropical regions around the globe, appears to have developed leaves that decompose more easily in its invaded range. Determining whether and how invasive species have more negative ecosystem consequences than native species can answer critical questions in ecology, evolution, and biogeography [52].
We detected higher NO3 levels in the soil from sites infested with all four invasive species compared to soils with native vegetation, even though these differences were not statistically significant. This trend is consistent with previous research indicating a positive relationship between the levels of soil NO3 and the abundance of invasive plant species [53]. Previous studies have also reported that plant invasions occur due to several mechanisms that help them uptake and use nutrients more efficiently [23,54]. The mechanisms associated with invasiveness appear to be different in nutrient-poor soils compared to nutrient-rich soils. Many studies indicate that the success of invasive plants in nutrient-poor soils is dependent on strategies such as increased nutrient-use efficiency [24,55], especially at shorter time scales [22]; long nutrient residence times [56]; high resistance to low levels of nutrients [57]; and high plasticity of stoichiometric ratios [23].
We also found that the soil concentrations of Na, Ca2+, and Mg2+ increase in the soils of invaded areas compared to soils from areas with native plants. According to Duda et al. [58], as Halogeton glomeratus invades native Krascheninnikovia lanata-dominated vegetation, soil concentrations of nitrate, phosphorus, potassium, and sodium increases with invasion. Rodgers et al. [59] reported that the availability of nitrogen, phosphorus, calcium, and magnesium in the soil, as well as the soil pH, were consistently and considerably higher in soils in North American temperate deciduous forests invaded by the European forb Alliaria petiolata. Similarly, our results reveal higher values for most soil properties and soil ions in sites invaded by the four invasive plant species included in this study, even if many of these values were not statistically significant (Table 1, Table 2, Table 3 and Table 4, Figure 1).
Invasive plants frequently play a role in modifying microelement (heavy metal) biogeochemical cycles in ecosystems [60]. Invading plants can increase soil microelement bioavailability in different ways: microelement secretion from leaves and release via humus breakdown and desorption. For instance, most rooted macrophytes can operate as a nutrient pump to absorb the nutrients contained in sediment pools, such as N and P, and subsequently remove them by tissue leaching and secretion and litter decomposition above the soil surface [61]. Invasive plants that perform a similar function can increase the bioavailability of soil microelements [62]. Additionally, microelements contained in plant tissues can be released into the environment by degradation of litter, other organic matter, and diagenesis [62,63]. During the decomposition of invasive plant litter, an increase in soil pH might indirectly result in the mobilization of microelements in the soil. Microelements can be released as sulfides from insoluble sulfide compounds in sediments. The microelements are then oxidized by sediment oxidation and enzymes in the roots of invasive plants [62]. In the same way, plant growth can cause microelements to be released from sediment particles into the pore water of wetlands and concentrate microelements in the rhizosphere of plants [64,65]. This can also be carried out by some native plants.
Increased mobilization can lead to higher rates of leaching and faster uptake of these microelements by plants, which can lower their total concentration in soil by a large amount [66], even though this would be expected to increase bioavailability. In addition to their role in the biogeochemical cycling of microelements, some invasive plants can reduce microelement mobility by adsorption, absorption, immobilization, and “returning elements to belowground pools”. Spartina alterniflora, the most pervasive invasive plant in China’s coastal wetland, has caused a considerable decrease in Cs137 and Pb210 adsorption from beach deposition of silt–fine sand and mud–sand [60]. The roots of hydrophytes, especially invasive hydrophytes, frequently show signs of iron plaque. As a major mechanism for tolerating hazardous metals, it adsorbs and immobilizes heavy metals efficiently, hence reducing their mobility [67]. Furthermore, invasive plants with interactions with symbiotic microorganisms may be able to sequester microelement toxicity through microbial biosynthesis of antioxidants and enzymes [68]. Therefore, the appearance of invasive species in areas contaminated with toxic microelements may not increase the potential enrichment of these toxic elements in the food chain and would in fact contribute to phytostabilization [69].
The results of this study reveal that the microelement values of soils sampled from three of the invasive species analyzed are generally lower than that of the soils from adjacent stands of native vegetation (Table 1, Table 2 and Table 3). The only exception to this pattern occurred for soils from sites invaded by O. ficus-indica (Table 4). Among the four invasive plant species, there were no significant differences in the levels of Mn, Zn, Fe, Cr, and Cu in the soils where they invaded; however, the levels of Co and Ni were significantly lower for soils invaded by P. jliflora, I. carnea, and L. leucocephala compared to those invaded by O. ficus-indica (Figure 2).
4. Materials and Methods
Saudi Arabia contains a variety of ecosystems, ranging from high-altitude mountains (Jabals) (up to 3050 m asl), valleys or river drainages that are seasonally dry (wadis), meadows (Raudhas), salt pans (Sabkhas), lava flows (Harrats), deep sands (Nafud), and irrigation canals. Winters are generally cool with occasional frost in the mountains of the northwestern region. The summer months are hot, with temperatures sometimes reaching above 50 °C. Rainfall is typically unpredictable and erratic in most parts of the country and mainly falls in winter and spring (100–150 mm), except for the southwestern region of the country, which can receive much more precipitation (>600 mm from September to November). Humidity levels are low in the interior of the country (15–20%) and high along the coast (55–75%).
4.1. Species Selection
Four species were selected for study: Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala and Opuntia ficus-indica. All these species form mono-specific stands in invaded localities and are recognized as being among the most invasive plant species in Saudi Arabia [34].
4.2. Field Sampling
Within each habitat, large stands of the four invasive plant species were selected to serve as “invaded” sampling locations. The size of the mono-specific stands varied and was usually 10–12 ha for P. juliflora, 2–6 ha for I. carnea, 0.6–1 ha for L. leucocephala, and 0.5–2 ha for O. ficus-indica. The invaded study sites were heavily infested by one of the four invasive species and were located on one side of a distinct ecotone. On the other side of the ecotone were areas consisting of native vegetation; these were the “native” sampling locations. Our experimental design therefore consisted of paired invaded and native sampling locations for each of the four invasive species. Fieldwork was conducted on 3 March 2022 for P. juliflora, 15 March 2022 for I. carnea, 3 April 2022 for L. leucocephala, and 23 April 2022 for O. ficus-indica.
Soil samples from the paired invaded and native sampling locations were generally located 20 m from each other. Soil samples were collected at a depth of 0–15 cm at each sampling location. At each invasive and native sampling location, a 20 m transect line was positioned perpendicular to the axis of the ecotone. Ten subsamples were collected at 2 m intervals along the transects established in invaded and native stands at each sampling location and these 10 subsamples were combined into one homogenized soil sample. Six sampling locations were selected for collecting paired soil samples for each of the four invasive plant species, yielding a total of six replicates per species. This approach means that we collected 120 soil subsamples for each species, and a total of 480 soil subsamples for the entire study.
4.3. Soil Analyses
Each soil sample replicate was separated into two subsamples. One subsample was dried at 105 °C for standard chemical assays. The soil pH and electrical conductivity (EC) were measured electrometrically in two different soil–H2O solutions (1:1, vol/vol and 1:2, vol/vol, respectively) [70]. The total carbon and nitrogen content were determined using dry combustion (FLASH EA1112 CHN analyzer; Thermo Fisher, Waltham, MA, USA). The available P was determined with the Olsen method [71]. Extractable microelements were quantified following digestion with diethylene-triamine-penta-acetic acid [72]. We determined the concentration of seven microelements: manganese (Mn), zinc (Zn), iron (Fe), chromium (Cr), cobalt (Co), copper (Cu), and nickel (Ni). The total microelement content was quantified following digestion with nitric acid [73]. The analysis was conducted using inductively coupled plasma optical emission spectrometry (Spectro Genesis; SPECTRO Analytical Instruments, Kleve, Germany). The second soil subsample was used to determine nitrate and ammonium colorimetrically, following 2 M potassium chloride extraction [74]. Nitrate and ammonium were quantified at the end of the incubation period.
4.4. Statistical Analysis
The data generated in this study were tested for normality using the Kolmogorov–Smirnov test and for homogeneity of variance using the Brown–Forsythe test. For each of the four invasive plant species, mean values were calculated based on the results of the six replicates for each species and variation in the data was calculated as the standard error (SE) of the mean. Paired t-tests were used to determine differences between invaded and native soil samples for each of the four invasive species. Statistically significant differences in the soil properties, ion concentrations, and microelement concentrations for the stands of the four invasive plants and the native stands were tested using one-way analysis of variance (ANOVA, F test) [75]. The Tukey’s honestly significant difference (HSD) test was used to determine the significance among means for treatments with a significant F-test. A significance level of p < 0.05 was applied for all analyses. All statistical analyses were performed using JMP (version 4.0 SAS Institute, Cary, NC, USA).
5. Conclusions
The results reported here do not support our prediction that we would detect significant changes in soil properties and microelements in areas infested by the four invasive plant species compared to the soils from stands of native vegetation. However, our results do indicate that the alteration of soil parameters varies among the four invasive plant species and varies for the different soil parameters we examined. For instance, the soils of sites with the four invasive plant species generally had higher values for soil properties compared to sites with native vegetation, but in most cases these differences were not statistically significant. The soils of stands invaded by P. juliflora and I. carnea had the largest number of statistically significant differences: four microelements and four soil properties, respectively. While the soils of sites invaded by O. ficus-indica and sites with native vegetation exhibited no significant differences for the 18 soil parameters. The soil of areas invaded by the four invasive species showed no significant differences for the three soil properties and the eight soil ions we measured, and no significant differences for five of the seven soil microelements. The levels of Co and Ni were significantly higher for the sites invaded by O. ficus-indica compared to the levels of the other three invasive species. Our results are in general agreement with previously published findings that indicate that the effects of invasive plants on soil dynamics vary idiosyncratically among invasive species and among invaded habitats (i.e., these effects are species- and habitat specific). These four invasive plants may simply be incapable of significantly altering soil properties and soil microelements. Conversely, our results do suggest a trend that might signal significant alteration of soil dynamics in the future, especially if the abundance of these invasive plant species increases. Thus, the invasion of these four plant species in southwest Saudi Arabia should be closely monitored and studied to determine their ecological consequences and their impacts on the native vegetation in this region.
Author Contributions
Conceptualization, A.M.A. and S.J.N.; methodology, A.M.A.; software, W.S.S.; validation, A.M.A., W.S.S., M.M.A., N.M.A. and S.J.N.; formal analysis, W.S.S.; investigation, A.M.A.; resources, A.M.A.; data curation, S.J.N.; writing—original draft preparation, W.S.S. and S.J.N.; writing—review and editing, A.M.A. and S.J.N.; visualization, W.S.S.; supervision, A.M.A., W.S.S. and S.J.N.; project administration, A.M.A., M.M.A. and N.M.A.; funding acquisition, A.M.A. and N.M.A. All authors have read and agreed to the published version of the manuscript.
Funding
The authors received funding for this study from the Deanship of Scientific Research at the King Khalid University through the Research Group Project under grant number (RGP. 2/92/43) and Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2023R356), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Data Availability Statement
Not applicable.
Acknowledgments
The authors extend their appreciation to the Deanship of Scientific Research at the King Khalid University for funding through the Research Group Project (RGP. 2/92/43) and Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2023R356), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia. SJN acknowledges financial support from the Department of Biological Sciences at Boise State University to work on this project and manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mack, R.N. The motivation for importing potentially invasive plant species: A primal urge. In Proceedings of the VI International Rangeland Congress, International Rangeland Congress, Aitkenvale, QLD, Australia, 19–23 July 1999; Volume 2, pp. 557–562. [Google Scholar]
- Lockwood, J.L.; Hoopes, M.F.; Marchetti, M.P. Invasion Ecology; John Wiley & Sons: New York, NY, USA, 2013; p. 444. [Google Scholar]
- Czortek, P.; Królak, E.; Borkowska, L.; Bielecka, A. Impacts of soil properties and functional diversity on the performance of invasive plant species Solidago canadensis L. on post-agricultural wastelands. Sci. Total Environ. 2020, 729, 139077. [Google Scholar] [CrossRef]
- Mack, R.N. Assessing the Extent, Status, and Dynamism of Plant Invasions: Current and Emerging Approaches. Invasive Species in a Changing World; Island Press: Washington, WA, USA, 2000; pp. 141–168. [Google Scholar]
- Davis, M.A. Biotic globalization: Does competition from introduced species threaten biodiversity? Bioscience 2003, 53, 481–489. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Vanparys, V.; Hayez, M.; Gruber, W.; Meerts, P. Impacts of alien plants on soil nutrients are correlated with initial site conditions in NW Europe. Oecologia 2008, 157, 131–140. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Gibbons, S.M.; Lekberg, Y.; Mummey, D.L.; Sangwan, N.; Ramsey, P.W.; Gilbert, J.A. Invasive plants rapidly reshape soil properties in a grassland ecosystem. MSystems 2017, 2, e00178-16. [Google Scholar] [CrossRef]
- Jamil, M.D.; Waheed, M.; Akhtar, S.; Bangash, N.; Chaudhari, S.K.; Majeed, M.; Hussain, M.; Ali, K.; Jones, D.A. Invasive plants diversity, ecological status, and distribution pattern in relation to edaphic factors in different habitat types of district Mandi Bahauddin, Punjab, Pakistan. Sustainability 2022, 14, 13312. [Google Scholar] [CrossRef]
- Clements, D.R.; Upadhyaya, M.K.; Joshi, S.; Shrestha, A. Global Plant Invasions; Springer: Cham, Switzerland, 2022; p. 381. [Google Scholar]
- Assaeed, A.M.; Alharthi, A.S.; Abd-ElGawad, A.M. Impacts of Nicotiana glauca Graham invasion on the vegetation composition and soil: A case study of Taif, western Saudi Arabia. Plants 2021, 10, 2587. [Google Scholar] [CrossRef] [PubMed]
- Unger, I.M.; Kremer, R.J.; Veum, K.S.; Goyne, K.W. Immediate and long-term effects of invasive plant species on soil characteristics. Soil Ecol. Lett. 2022, 4, 276–288. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Kourtev, P.; Huang, W. Changes in soil functions following invasions of exotic understory plants in deciduous forests. Ecol. Appl. 2001, 11, 1287–1300. [Google Scholar] [CrossRef]
- Weidenhammer, J.D.; Callaway, R.M. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J. Chem. Ecol. 2010, 36, 59–69. [Google Scholar] [CrossRef]
- Qu, T.; Du, X.; Peng, Y.; Guo, W.; Zhao, C.; Losapio, G. Invasive species allelopathy decreases plant growth and soil microbial activity. PLoS ONE 2021, 16, e0246685. [Google Scholar] [CrossRef] [PubMed]
- Coykendall, K.E.; Houseman, G.R. Lespedeza cuneata invasion alters soils facilitating its own growth. Biol. Invasions 2014, 16, 1735–1742. [Google Scholar] [CrossRef]
- Anning, A.K.; Gyamfi, B.; Effah, A.T. Broussonetia papyrifera controls nutrient return to soil to facilitate its invasion in a tropical forest of Ghana. J. Plant Ecol. 2018, 11, 909–918. [Google Scholar] [CrossRef]
- Wang, W.; Sardans, J.; Wang, C.; Asensio, D.; Bartrons, M.; Peñuelas, J. Species-specific impacts of invasive plant success on vertical profiles of soil carbon accumulation and nutrient retention in the Minjiang River tidal estuarine wetlands of China. Soil Syst. 2018, 2, 5. [Google Scholar] [CrossRef]
- Maass, J.M.; Balvanera, P.; Castillo, A.; Daily, G.C.; Mooney, H.A.; Ehrlich, P.; Quesada, M.; Miranda, A.; Jaramillo, V.J.; García-Oliva, F.; et al. Ecosystem services of tropical dry forests: Insights from long-term ecological and social research on the Pacific Coast of Mexico. Ecol. Soc. 2005, 10. [Google Scholar] [CrossRef]
- Vasquez-Valderrama, M.; González-M, R.; López-Camacho, R.; Baptiste, M.P.; Salgado-Negret, B. Impact of invasive species on soil hydraulic properties: Importance of functional traits. Biol. Invasions 2020, 22, 1849–1863. [Google Scholar] [CrossRef]
- Liao, C.; Peng, R.; Luo, Y.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L.; Vitousek, P.M. Resource-use efficiency and plant invasion in low-resource systems. Nature 2007, 446, 1079–1081. [Google Scholar] [CrossRef]
- González, A.L.; Kominoski, J.S.; Danger, M.; Ishida, S.; Iwai, N.; Rubach, A. Can ecological stoichiometry help explain patterns of biological invasions? Oikos 2010, 119, 779–790. [Google Scholar] [CrossRef]
- Matzek, V. Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment. Biol. Invasions 2011, 13, 3005–3014. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [PubMed]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, J.G.; Stander, E.K. Habitat function in urban riparian zones. Urban Ecosyst. Ecol. 2010, 55, 103–118. [Google Scholar]
- Elgersma, K.J.; Ehrenfeld, J.G.; Yu, S.; Vor, T. Legacy effects overwhelm the short-term effects of exotic plant invasion and restoration on soil microbial community structure, enzyme activities, and nitrogen cycling. Oecologia 2011, 167, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Davis, M.A.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; Ewel, J.J.; Vermeij, G.J.; Brown, J.H.; Rosenzweig, M.L.; Gardener, M.R.; Carroll, S.P.; et al. Don’t judge species on their origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef]
- Reaser, J.K.; Simpson, A.; Guala, G.F.; Morisette, J.T.; Fuller, P. Envisioning a national invasive species information framework. Biol. Invasions 2020, 22, 21–36. [Google Scholar] [CrossRef]
- Vujanović, D.; Losapio, G.; Milić, S.; Milić, D. The Impact of multiple species invasion on soil and plant communities increases with invasive species co-occurrence. Front. Plant Sci. 2022, 13, 875824. [Google Scholar] [CrossRef]
- Jamin, J.; Diehl, D.; Meyer, M.; Davis, J.; Schaumann, G.E. Physio-chemical soil properties affected by invasive plants in southwest Germany (Rhineland-Palatinate)—A case study. Soil Syst. 2022, 6, 93. [Google Scholar] [CrossRef]
- Thomas, J.; El-Sheikh, M.A.; Alfarhan, A.H.; Alatar, A.A.; Sivadasan, M.; Basahi, M.; Al-Obaid, S.; Rajakrishnan, R. Impact of alien invasive species on habitats and species richness in Saudi Arabia. J. Arid Environ. 2016, 127, 53–65. [Google Scholar] [CrossRef]
- Lehan, N.E.; Murphy, J.R.; Thorburn, L.P.; Bradley, B.A. Accidental introductions are an important source of invasive plants in the continental United States. Am. J. Bot. 2013, 100, 1287–1293. [Google Scholar] [CrossRef]
- Alfaran, A.H.; Thomas, J.; Rajakrishnan, R. Impact assessment and management of invasive species in plant diversity centers and agricultural fields in Saudi Arabia. In Invasive Alien Species: Observations and Issues from Around the World. Volume 2: Issues and Invasions in Asia and the Pacific Region; Pullaiah, T., Ielmini, M.R., Eds.; John Wiley & Sons: New York, NY, USA, 2021; pp. 207–225. [Google Scholar]
- Al-Sodany, Y.M. A new record to the flora of Saudi Arabia: Ipomoea carnea Jacq., Convolvulaceae. World J. Res. Rev. 2016, 3, 25–30. [Google Scholar]
- Shaltout, K.H.; Al-Sodany, Y.M.; Eid, E.M. The biology of Egyptian woody perennials-2. Ipomoea Carnea Jacq. Ass. Univ. Bull. Environ. Res. 2006, 9, 75–91. [Google Scholar]
- Waring, B.G.; Alvarez-Cansino, L.; Barry, K.E.; Becklund, K.K.; Dale, S.; Gei, M.G.; Keller, A.B.; Lopez, O.R.; Markesteijn, L.; Mangan, S.; et al. Pervasive and strong effects of plants on soil chemistry: A meta-analysis of individual plant ‘Zinke’ effects. Proc. R. Soc. B. 2015, 282, 20151001. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Skočajić, D.; Nešić, M.L. Invasive Species: Routes of Introduction, Establishment, and Expansion; Springer: Berlin/Heidelberg, Germany, 2021; pp. 571–582. [Google Scholar]
- Gaertner, M.; Den Breeyen, A.; Hui, C.; Richardson, D.M. Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: A meta-analysis. Prog. Phys. Geogr. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Joshi, C.; De Leeuw, J.; van Duren, I.C. Remote sensing and GIS applications for mapping and spatial modelling of invasive species. Proc. ISPRS 2004, 35, B7. [Google Scholar]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Godoy, O.; Alonso, A.; Gallardo, A.; Saldaña, A. What explains variation in the impacts of exotic plant invasions on the nitrogen cycle? A meta-analysis. Ecol. Lett. 2014, 17, 1–12. [Google Scholar] [CrossRef]
- Ashton, I.W.; Hyatt, L.A.; Howe, K.M.; Gurevitch, J.; Lerdau, M.T. Invasive species accelerate decomposition and litter nitrogen loss in a mixed deciduous forest. Ecol. Appl. 2005, 15, 1263–1272. [Google Scholar] [CrossRef]
- Feng, Y.L.; Lei, Y.B.; Wang, R.F.; Callaway, R.M.; Valiente-Banuet, A.; Inderjit; Li, Y.P.; Zheng, Y.L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proc. Natl. Acad. Sci. USA 2009, 106, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, J.G. Implications of invasive species for belowground community and nutrient processes. Weed Technol. 2004, 18, 1232–1235. [Google Scholar] [CrossRef]
- Rout, M.E.; Callaway, R.M. An invasive plant paradox. Science 2009, 324, 734. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.J. The role of evolution in the invasion process. Proc. Natl. Acad. Sci. USA 2007, 104, 3671–3672. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.-Y.; Julien, M.H.; Fatemi, M.; Girod, C.; van Klinken, R.; Gross, C.L.; Novak, S.J. Phenotypic divergence during the invasion of Pyla canescens in Australia and France: Evidence for selection-driven evolution. Ecol. Lett. 2010, 13, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Sax, D.F.; Stachowicz, J.J.; Gaines, S.D. (Eds.) Species Invasions: Insights into Ecology, Evolution and Biogeography; Sinauer: Sunderland, MA, USA, 2005. [Google Scholar]
- Gilliam, F.S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef]
- Daehler, C.C. Performance comparisons of co-occurring native and alien invasive plants: Implications for conservation and restoration. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 183–211. [Google Scholar] [CrossRef]
- Ostertag, R.; Verville, J.H. Fertilization with nitrogen and phosphorus increases abundance of non-native species in Hawaiian montane forests. Plant Ecol. 2002, 162, 77–90. [Google Scholar] [CrossRef]
- Laungani, R.; Knops, J.M. Species-driven changes in nitrogen cycling can provide a mechanism for plant invasions. Proc. Natl. Acad. Sci. USA 2009, 106, 12400–12405. [Google Scholar] [CrossRef]
- Kueffer, C. Reduced risk for positive soil-feedback on seedling regeneration by invasive trees on a very nutrient-poor soil in Seychelles. Biol. Invasions 2010, 12, 97–102. [Google Scholar] [CrossRef]
- Duda, J.J.; Freeman, D.C.; Emlen, J.M.; Belnap, J.; Kitchen, S.G.; Zak, J.C.; Sobek, E.; Tracy, M.; Montante, J. Differences in native soil ecology associated with invasion of the exotic annual chenopod, Halogeton glomeratus. Biol. Fertil. Soils 2003, 38, 72–77. [Google Scholar] [CrossRef]
- Rodgers, V.L.; Wolfe, B.E.; Werden, L.K.; Finzi, A.C. The invasive species Alliaria petiolata (garlic mustard) increases soil nutrient availability in northern hardwood-conifer forests. Oecologia 2008, 157, 459–471. [Google Scholar] [CrossRef]
- Li, H.; Meng, Z.; Dang, X.; Yang, P. Soil Properties under artificial mixed forests in the desert-yellow river coastal transition zone, China. Forests 2022, 13, 1174. [Google Scholar] [CrossRef]
- Odum, W.E. Comparative ecology of tidal freshwater and salt marshes. Annu. Rev. Ecol. Syst. 1988, 19, 147–176. [Google Scholar] [CrossRef]
- Windham, L.; Weis, J.S.; Weis, P. Patterns and processes of mercury release from leaves of two dominant salt marsh macrophytes, Phragmites australis and Spartina alterniflora. Estuaries 2001, 24, 787–795. [Google Scholar] [CrossRef]
- Ermakov, V.V.; Korobova, E.M.; Degtyarev, A.P.; Tyutikov, S.F.; Karpova, E.A.; Petrunina, N.S. Impact of natural and man-made factors on migration of heavy metals in the Ardon River basin (North Ossetia). J. Soils Sediments 2016, 16, 1253–1266. [Google Scholar] [CrossRef]
- Chai, M.; Shi, F.; Li, R.; Shen, X. Heavy metal contamination and ecological risk in Spartina alterniflora marsh in intertidal sediments of Bohai Bay, China. Mar. Pollut. Bull. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Li, J.; Lu, H.; Liu, J.; Hong, H.; Yan, C. The influence of flavonoid amendment on the absorption of cadmium in Avicennia marina roots. Ecotoxicol. Environ. Saf. 2015, 120, 1–6. [Google Scholar] [CrossRef]
- Selvi, F.; Carrari, E.; Colzi, I.; Coppi, A.; Gonnelli, C. Responses of serpentine plants to pine invasion: Vegetation diversity and nickel accumulation in species with contrasting adaptive strategies. Sci. Total Environ. 2017, 595, 72–80. [Google Scholar] [CrossRef]
- Li, J.; Du, Z.; Zou, C.B.; Dai, Z.; Du, D.; Yan, C. The mutual restraint effect between the expansion of Alternanthera philoxeroides (Mart.) Griseb and cadmium mobility in aquatic environment. Ecotoxicol. Environ. Saf. 2018, 148, 237–243. [Google Scholar] [CrossRef]
- El-Bakatoushi, R.; Alframawy, A.M.; Tammam, A.; Youssef, D.; El-Sadek, L. Molecular and physiological mechanisms of heavy metal tolerance in Atriplex halimus. Int. J. Phytoremed. 2015, 17, 789–800. [Google Scholar] [CrossRef]
- Ellili, A.; Rabier, J.; Prudent, P.; Salducci, M.D.; Heckenroth, A.; Lachaâl, M.; Laffont-Schwob, I. Decision-making criteria for plant-species selection for phytostabilization: Issues of biodiversity and functionality. J. Environ. Manag. 2017, 201, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.A.; Sommers, L. Total carbon, organic carbon, and organic matter. Methods Soil Anal. Part 2 Chem. Microbiol. Prop. 1983, 9, 539–579. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate (No. 939); US Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Berghage, R.D.; Krauskopf, D.M.; Warncke, D.D.; Widders, I. Micronutrient testing of plant growth media: Extractant identification and evaluation. Commun. Soil Sci. Plant Anal. 1987, 18, 1089–1109. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Warman, P.R. Comparison of three digestion methods for the recovery of 17 plant essential nutrients and trace elements from six composts. Compos. Sci. Util. 2002, 10, 197–203. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989; p. 1191. [Google Scholar]
Figure 1.
Values of soil properties and ion concentrations, pH (a), EC (b), organic matter [OM] (c), Na (d), K (e), Ca (f), Mg (g), SO4 (h), NO3 (i), PO4 (j), and ammonia (k), from stands invaded by Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica and adjacent stands of native vegetation in southwest Saudi Arabia. Different capital letters indicate significant differences among the soils from the stands invaded by four invasive plant species and different lower-case letters indicate significant differences among the soils from stands of native vegetation (n = 6, p < 0.05).
Figure 1.
Values of soil properties and ion concentrations, pH (a), EC (b), organic matter [OM] (c), Na (d), K (e), Ca (f), Mg (g), SO4 (h), NO3 (i), PO4 (j), and ammonia (k), from stands invaded by Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica and adjacent stands of native vegetation in southwest Saudi Arabia. Different capital letters indicate significant differences among the soils from the stands invaded by four invasive plant species and different lower-case letters indicate significant differences among the soils from stands of native vegetation (n = 6, p < 0.05).

Figure 2.
Values of soil microelements, Mn (a), Zn (b), Fe (c), Cr (d), Co (e), Cu (f), and Ni (g), from stands invaded by Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica and adjacent stands of native vegetation in southwest Saudi Arabia. Different capital letters indicate significant differences among the soils from stands invaded by four invasive plant species and different lower-case letters indicate significant differences among the soils from stands of native vegetation (n = 6, p < 0.05).
Figure 2.
Values of soil microelements, Mn (a), Zn (b), Fe (c), Cr (d), Co (e), Cu (f), and Ni (g), from stands invaded by Prosopis juliflora, Ipomoea carnea, Leucaena leucocephala, and Opuntia ficus-indica and adjacent stands of native vegetation in southwest Saudi Arabia. Different capital letters indicate significant differences among the soils from stands invaded by four invasive plant species and different lower-case letters indicate significant differences among the soils from stands of native vegetation (n = 6, p < 0.05).


{kind=link}
{kind=link}
Table 1.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Prosopis juliflora and adjacent stands with native vegetation in southwest Saudi Arabia.
Table 1.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Prosopis juliflora and adjacent stands with native vegetation in southwest Saudi Arabia.
| Invasive | Native | t-Test | p-Values | |
|---|---|---|---|---|
| Soil properties | ||||
| pH | 7.35 ± 0.15 | 7.13 ± 0.03 | 1.960 | 0.234 |
| EC μS | 3837.67 ± 1712.79 | 1797.67 ± 357.34 | 1.359 | 0.308 |
| OM % | 8.03 ± 1.99 | 5.39 ± 0.31 | 1.726 | 0.259 |
| Ions (cations and anions) | ||||
| Na⁺ ppm | 88.09 ± 33.55 | 87.95 ± 12.63 | 0.001 | 0.982 |
| K⁺ ppm | 454.52 ± 77.13 | 435.58 ± 20.53 | 0.056 | 0.824 |
| Ca2+ ppm | 0.92 ± 0.28 | 0.72 ± 0.06 | 0.477 | 0.528 |
| Mg2+ ppm | 1.03 ± 0.49 | 0.62 ± 0.25 | 0.551 | 0.499 |
| SO42− ppm | 46.68 ± 6.28 | 54.89 ± 11.58 | 0.389 | 0.567 |
| NO3− ppm | 666.67 ± 425.57 | 60.00 ± 20.00 | 2.028 | 0.228 |
| PO43− ppm | 4.08 ± 0.60 | 3.59 ± 1.66 | 0.078 | 0.794 |
| Ammonia (NH4+) ppm | 233.33 ± 88.19 | 100.00 ± 23.09 | 2.139 | 0.217 |
| Microelements | ||||
| Mn ppm | 172.92 ± 122.92 | 270.83 ± 90.60 | 0.411 | 0.556 |
| Zn mg/L | 43.54 ± 3.15 | 67.92 ± 8.90 | 6.665 | 0.061 |
| Fe ppm | 10,445.83 ± 3732.17 | 22,385.42 ± 7700.57 | 1.947 | 0.235 |
| Cr ppm | 4.99 ± 0.47 | 12.05 ± 0.90 | 48.472 | 0.002 |
| Co ppm | 4.25 ± 0.48 | 8.91 ± 0.42 | 53.902 | 0.002 |
| Cu ppm | 5.42 ± 1.63 | 12.50 ± 0.63 | 16.514 | 0.015 |
| Ni ppm | 9.69 ± 1.31 | 79.58 ± 9.42 | 54.015 | 0.002 |
Data represented as means ± SE, n = 6.
Table 2.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Ipomoea carnea and adjacent stands with native vegetation in southwest Saudi Arabia.
Table 2.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Ipomoea carnea and adjacent stands with native vegetation in southwest Saudi Arabia.
| Invasive | Native | t-Test | p-Values | |
|---|---|---|---|---|
| Soil properties | ||||
| pH | 7.11 ± 0.09 | 7.73 ± 0.07 | 28.782 | 0.006 |
| EC μS | 1736.67 ± 777.63 | 357.33 ± 63.34 | 3.126 | 0.152 |
| OM % | 8.44 ± 1.17 | 4.99 ± 0.20 | 8.393 | 0.044 |
| Ions (cations and anions) | ||||
| Na⁺ ppm | 70.61 ± 28.87 | 58.42 ± 21.24 | 0.116 | 0.751 |
| K⁺ ppm | 334.65 ± 90.64 | 372.31 ± 79.34 | 0.098 | 0.770 |
| Ca2+ ppm | 0.77 ± 0.15 | 0.27 ± 0.04 | 10.843 | 0.030 |
| Mg2+ ppm | 0.49 ± 0.16 | 0.21 ± 0.03 | 2.770 | 0.171 |
| SO42− ppm | 49.25 ± 0.52 | 31.27 ± 0.61 | 511.249 | 0.001 |
| NO3− ppm | 80.00 ± 11.55 | 73.33 ± 24.04 | 0.063 | 0.815 |
| PO43− ppm | 4.42 ± 2.04 | 2.67 ± 0.73 | 0.651 | 0.465 |
| Ammonia (NH4+) ppm | 133.33 ± 40.55 | 40.00 ± 11.55 | 4.900 | 0.091 |
| Microelements | ||||
| Mn ppm | 229.17 ± 182.30 | 269.58 ± 126.90 | 0.033 | 0.864 |
| Zn mg/L | 55.42 ± 11.59 | 45.00 ± 17.14 | 0.253 | 0.641 |
| Fe ppm | 10,564.58 ± 2597.91 | 16,856.25 ± 9276.55 | 0.427 | 0.549 |
| Cr ppm | 9.96 ± 6.17 | 12.97 ± 7.86 | 0.091 | 0.778 |
| Co ppm | 7.52 ± 2.82 | 9.76 ± 4.41 | 0.183 | 0.691 |
| Cu ppm | 10.21 ± 6.01 | 19.38 ± 9.57 | 0.658 | 0.463 |
| Ni ppm | 35.69 ± 25.95 | 66.25 ± 32.37 | 0.543 | 0.502 |
Data represented as means ± SE, n = 6.
Table 3.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Leucaena leucocephala and adjacent stands with native vegetation in southwest Saudi Arabia.
Table 3.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Leucaena leucocephala and adjacent stands with native vegetation in southwest Saudi Arabia.
| Invasive | Native | t-Test | p-Values | |
|---|---|---|---|---|
| Soil properties | ||||
| pH | 6.97 ± 0.22 | 7.64 ± 0.07 | 8.894 | 0.041 |
| EC μS | 7250.00 ± 4145.68 | 324.00 ± 12.22 | 2.791 | 0.170 |
| OM % | 12.22 ± 4.06 | 4.19 ± 0.09 | 3.901 | 0.119 |
| Ions (cations and anions) | ||||
| Na⁺ ppm | 297.22 ± 221.53 | 77.89 ± 31.77 | 0.960 | 0.383 |
| K⁺ ppm | 468.29 ± 41.84 | 536.73 ± 139.35 | 0.221 | 0.663 |
| Ca2+ ppm | 1.52 ± 1.07 | 0.38 ± 0.16 | 1.096 | 0.354 |
| Mg2+ ppm | 1.17 ± 0.44 | 0.48 ± 0.20 | 2.029 | 0.227 |
| SO42− ppm | 56.42 ± 12.45 | 41.07 ± 8.84 | 1.010 | 0.372 |
| NO3− ppm | 173.33 ± 75.13 | 43.33 ± 8.82 | 2.953 | 0.161 |
| PO43− ppm | 2.33 ± 0.30 | 15.20 ± 10.33 | 1.551 | 0.281 |
| Ammonia (NH4+) ppm | 73.33 ± 13.33 | 53.33 ± 24.04 | 0.529 | 0.507 |
| Microelements | ||||
| Mn ppm | 73.33 ± 13.33 | 53.33 ± 24.04 | 0.529 | 0.507 |
| Zn mg/L | 57.71 ± 10.28 | 40.63 ± 15.66 | 0.831 | 0.413 |
| Fe ppm | 23,525.00 ± 9341.35 | 17,068.75 ± 10,154.34 | 0.219 | 0.664 |
| Cr ppm | 9.60 ± 1.21 | 12.24 ± 8.09 | 0.104 | 0.763 |
| Co ppm | 7.52 ± 0.34 | 8.09 ± 4.66 | 0.015 | 0.908 |
| Cu ppm | 12.29 ± 3.61 | 16.25 ± 10.23 | 0.133 | 0.734 |
| Ni ppm | 18.52 ± 2.13 | 45.50 ± 37.03 | 0.529 | 0.507 |
Data represented as means ± SE, n = 6.
Table 4.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Opuntia ficus-indica and adjacent stands with native vegetation in southwest Saudi Arabia.
Table 4.
Soil properties, ion concentrations, and microelement concentrations of soils in stands invaded by Opuntia ficus-indica and adjacent stands with native vegetation in southwest Saudi Arabia.
| Invasive | Native | t-Test | p-Values | |
|---|---|---|---|---|
| Soil properties | ||||
| pH | 7.08 ± 0.04 | 7.09 ± 0.03 | 0.084 | 0.786 |
| EC μS | 288.60 ± 68.97 | 191.23 ± 20.50 | 1.831 | 0.247 |
| OM % | 5.68 ± 2.14 | 3.36 ± 0.80 | 1.026 | 0.368 |
| Ions (cations and anions) | ||||
| Na⁺ ppm | 51.36 ± 6.46 | 35.95 ± 6.49 | 2.828 | 0.168 |
| K⁺ ppm | 545.34 ± 98.00 | 491.97 ± 167.64 | 0.076 | 0.797 |
| Ca2+ ppm | 0.27 ± 0.04 | 0.25 ± 0.00 | 0.143 | 0.725 |
| Mg2+ ppm | 0.20 ±0.02 | 0.17 ± 0.05 | 1.690 | 0.263 |
| SO42− ppm | 37.47 ± 0.96 | 31.50 ± 3.56 | 2.616 | 0.181 |
| NO3− ppm | 126.67 ± 86.67 | 46.67 ± 6.67 | 0.847 | 0.409 |
| PO43− ppm | 17.70 ± 9.75 | 1.17 ± 0.92 | 2.848 | 0.167 |
| Ammonia (NH4+) ppm | 86.67 ± 13.33 | 73.33 ± 13.33 | 0.500 | 0.519 |
| Microelements | ||||
| Mn ppm | 312.50 ± 160.20 | 181.25 ± 76.38 | 0.547 | 0.501 |
| Zn mg/L | 81.25 ± 29.05 | 32.50 ± 6.68 | 2.674 | 0.177 |
| Fe ppm | 17,970.83 ± 5340.86 | 9552.08 ± 2189.11 | 2.127 | 0.218 |
| Cr ppm | 14.43 ± 3.84 | 33.13 ± 24.06 | 0.589 | 0.486 |
| Co ppm | 12.40 ± 0.74 | 10.10 ± 2.36 | 0.869 | 0.404 |
| Cu ppm | 18.96 ± 4.71 | 17.71 ± 4.65 | 0.036 | 0.859 |
| Ni ppm | 133.13 ± 17.51 | 52.78 ± 33.61 | 4.493 | 0.101 |
Data represented as means ± SE, n = 6.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abbas, A.M.; Soliman, W.S.; Alomran, M.M.; Alotaibi, N.M.; Novak, S.J. Four Invasive Plant Species in Southwest Saudi Arabia Have Variable Effects on Soil Dynamics. Plants 2023, 12, 1231. https://doi.org/10.3390/plants12061231
AMA Style
Abbas AM, Soliman WS, Alomran MM, Alotaibi NM, Novak SJ. Four Invasive Plant Species in Southwest Saudi Arabia Have Variable Effects on Soil Dynamics. Plants. 2023; 12(6):1231. https://doi.org/10.3390/plants12061231
Chicago/Turabian StyleAbbas, Ahmed M., Wagdi S. Soliman, Maryam M. Alomran, Nahaa M. Alotaibi, and Stephen J. Novak. 2023. "Four Invasive Plant Species in Southwest Saudi Arabia Have Variable Effects on Soil Dynamics" Plants 12, no. 6: 1231. https://doi.org/10.3390/plants12061231
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.