
Seed Size, Seed Dispersal Traits, and Plant Dispersion Patterns for Native and Introduced Grassland Plants

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
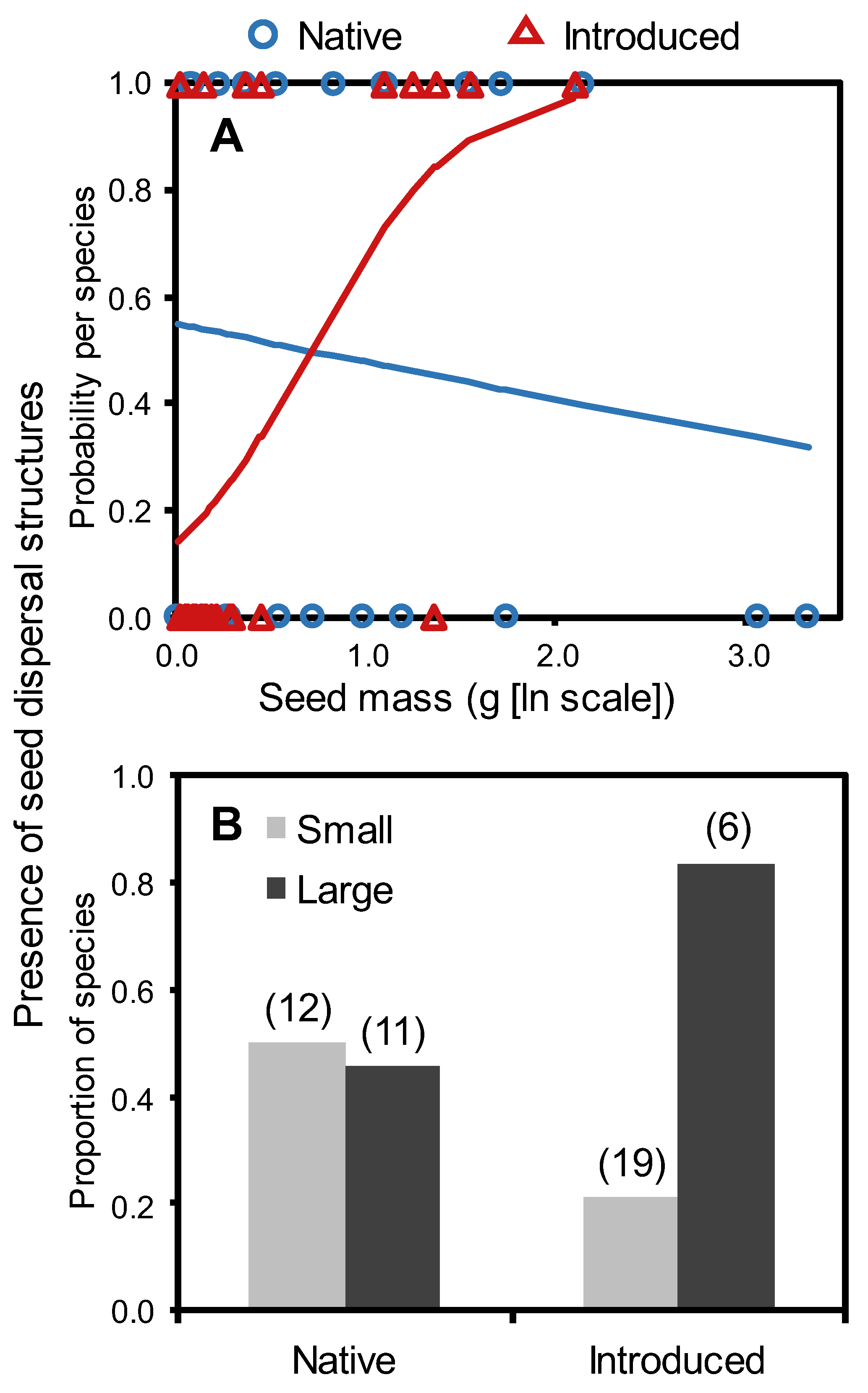
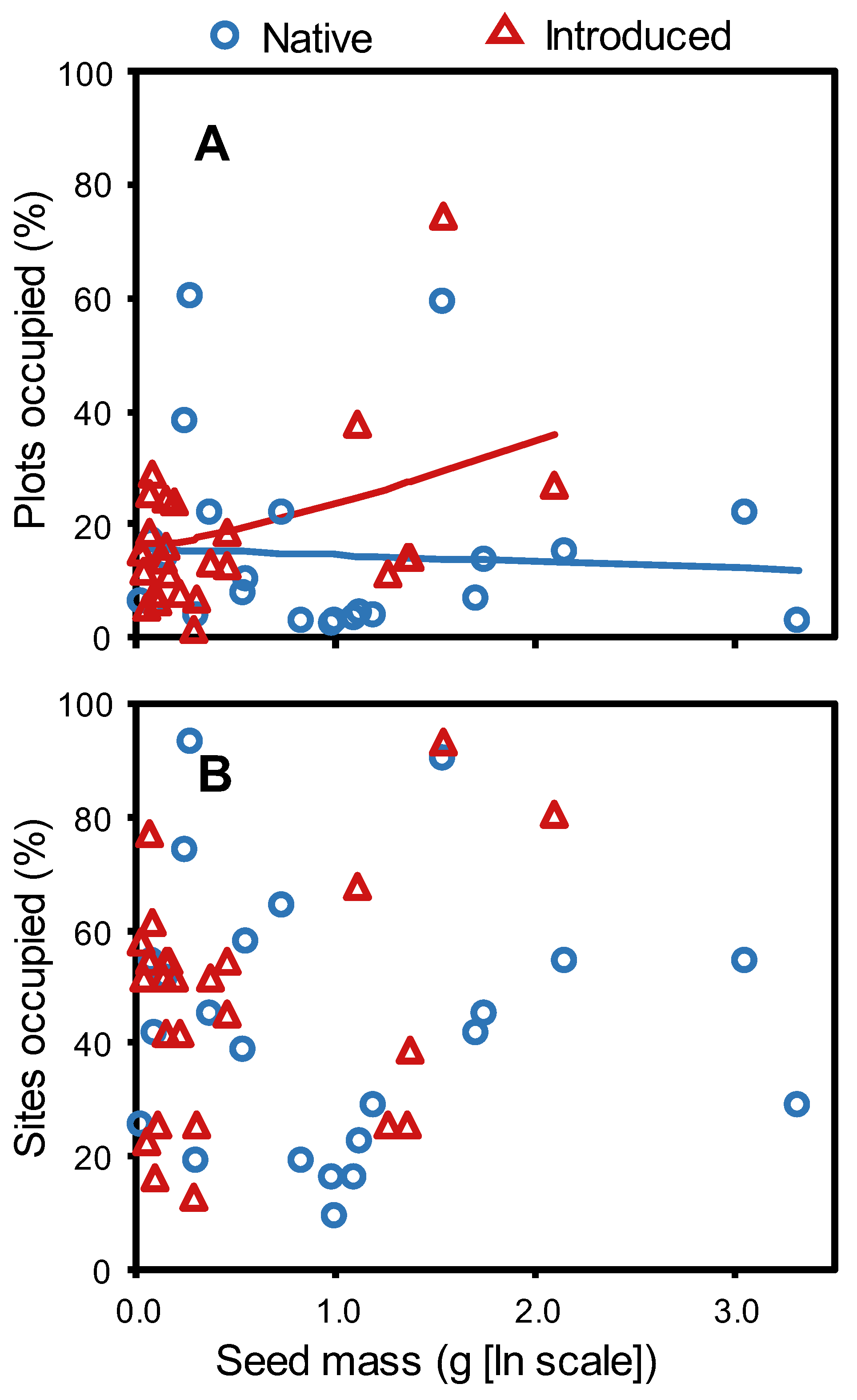
2. Results
3. Discussion
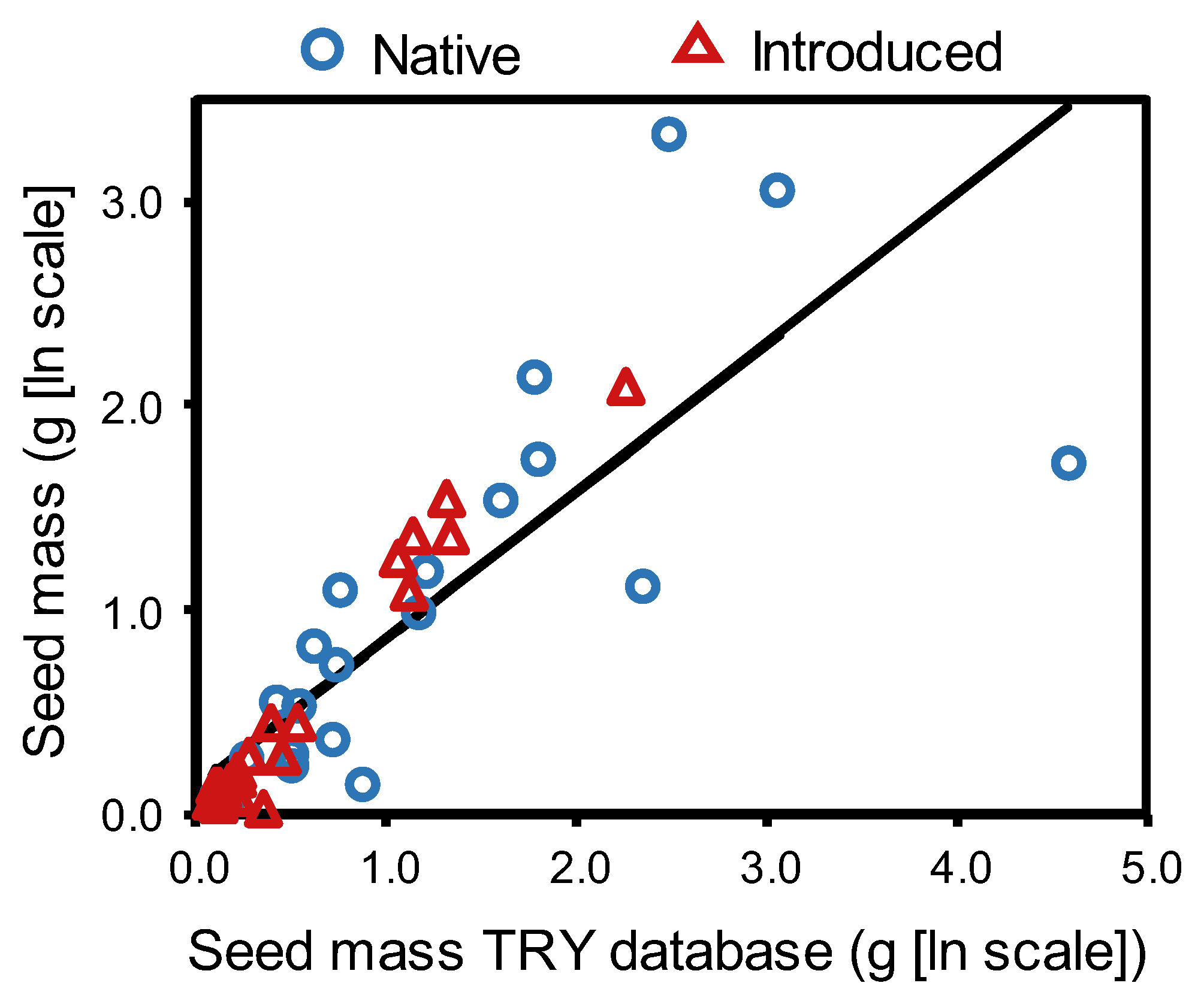
4. Materials and Methods
Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In Dynamics of Populations; Den Boer, P.J., Gradwell, G.R., Eds.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1971; pp. 298–312. [Google Scholar]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Comita, L.S.; Queenborough, S.A.; Murphy, S.J.; Eck, J.L.; Xu, K.; Krishnadas, M.; Beckman, N.; Zhu, Y.; Gómez-Aparicio, L. Testing predictions of the Janzen-Connell hypothesis: A meta-analysis of experimental evidence for distance-and density-dependent seed and seedling survival. J. Ecol. 2014, 102, 845–856. [Google Scholar] [CrossRef]
- Petermann, J.S.; Fergus, A.J.F.; Turnbull, L.A.; Schmid, B. Janzen-Connell Effects are Widespread and Strong Enough to Maintain Diversity in Grasslands. Ecology 2008, 89, 2399–2406. [Google Scholar] [CrossRef]
- Song, X.; Lim, J.Y.; Yang, J.; Luskin, M.S. When do Janzen–Connell effects matter? A phylogenetic meta-analysis of conspecific negative distance and density dependence experiments. Ecol. Lett. 2021, 24, 608–620. [Google Scholar] [CrossRef]
- Burgess, S.C.; Baskett, M.L.; Grosberg, R.K.; Morgan, S.G.; Strathmann, R.R. When is dispersal for dispersal? Unifying marine and terrestrial perspectives. Biol. Rev. 2016, 91, 867–882. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Poschlod, P.; Antonelli, A.; Liu, U.; Dickie, J. Trade-off between dispersal in space and time. Ecol. Lett. 2020, 23, 1635–1642. [Google Scholar] [CrossRef]
- Sonkoly, J.; Deák, B.; Valkó, O.; Molnár, V.A.; Tóthmérész, B.; Török, P. Do large-seeded herbs have a small range size? The seed mass–distribution range trade-off hypothesis. Ecol. Evol. 2017, 7, 11204–11212. [Google Scholar] [CrossRef]
- Turnbull, L.A.; Rees, M.; Crawley, M.J. Seed mass and the competition/colonization trade-off: A sowing experiment. J. Ecol. 1999, 87, 899–912. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Gambín, B.L.; Borrás, L. Resource Distribution and the Trade-off between Seed Number and Seed Weight: A Comparison across Crop Species. Ann. Appl. Biol. 2009, 156, 91–102. [Google Scholar] [CrossRef]
- Muller-Landau, H.C. The tolerance-fecundity trade-off and the maintenance of diversity in seed size. Proc. Natl. Acad. Sci. USA 2010, 107, 4242–4247. [Google Scholar] [CrossRef] [Green Version]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Tamme, R.; Götzenberger, L.; Zobel, M.; Bullock, J.M.; Hooftman, D.A.; Kaasik, A.; Pärtel, M. Predicting species’ maximum dispersal distances from simple plant traits. Ecology 2014, 95, 505–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moles, A.T.; Falster, D.S.; Leishman, M.R.; Westoby, M. Small-seeded species produce more seeds per square metre of canopy per year, but not per individual per lifetime. J. Ecol. 2004, 92, 384–396. [Google Scholar] [CrossRef]
- Sullivan, L.L.; Clark, A.T.; Tilman, D.; Shaw, A.K. Mechanistically derived dispersal kernels explain species-level patterns of recruitment and succession. Ecology 2018, 99, 2415–2420. [Google Scholar] [CrossRef] [Green Version]
- Benkman, C.W. Wind dispersal capacity of pine seeds and the evolution of different seed dispersal modes in pines. Oikos 1995, 73, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Jansen, P.A.; Bongers, F.; Hemerik, L. Seed mass and mast seeding enhance dispersal by a neotropical scatter-hoarding rodent. Ecol. Monogr. 2004, 74, 569–589. [Google Scholar] [CrossRef]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Ramp, D.; Ren, S.; Kingsford, R.T. Chasing the unknown: Predicting seed dispersal mechanisms from plant traits. J. Ecol. 2010, 98, 1310–1318. [Google Scholar] [CrossRef]
- Vander Wall, S.B.; Beck, M.J. A comparison of frugivory and scatter-hoarding seed-dispersal syndromes. Bot. Rev. 2012, 78, 10–31. [Google Scholar] [CrossRef]
- Westoby, M.; Leishman, M.; Lord, J. Comparative ecology of seed size and dispersal. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1309–1318. [Google Scholar]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Weiher, E.; Keddy, P.A. Ecological Assembly Rules: Perspectives, Advances, Retreats; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Murray, B.R.; Phillips, M.L. Investment in seed dispersal structures is linked to invasiveness in exotic plant species of south-eastern Australia. Biol. Invasions 2010, 12, 2265–2275. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Traits Associated with Invasiveness in Alien Plants: Where Do we Stand? In Biological Invasions. Ecological Studies; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 193. [Google Scholar] [CrossRef]
- Flores-Moreno, H.; Thomson, F.J.; Warton, D.I.; Moles, A.T. Are Introduced Species Better Dispersers than Native Species? A Global Comparative Study of Seed Dispersal Distance. PLoS ONE 2013, 8, e68541. [Google Scholar] [CrossRef]
- Daws, M.I.; Hall, J.; Flynn, S.; Pritchard, H.W. Do invasive species have bigger seeds? Evidence from intra- and inter-specific comparisons. S. Afr. J. Bot. 2007, 73, 138–143. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, S.; Martin, T.G.; Heard, K.M.; Kinloch, J. Plant traits predict impact of invading species: An analysis of herbaceous vegetation in the subtropics. Aust. J. Bot. 2005, 53, 757–770. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Lovett-Doust, J. Ecological and taxonomic differences between native and introduced plants of southwestern Ontario. Ecoscience 2001, 8, 230–238. [Google Scholar] [CrossRef]
- Maron, J.L.; Pearson, D.E.; Potter, T.; Ortega, Y.K. Seed size and provenance mediate the joint effects of disturbance and seed predation on community assembly. J. Ecol. 2012, 100, 1492–1500. [Google Scholar] [CrossRef]
- Moravcová, L.; Pyšek, P.; Jarošík, V.; Havlíčková, V.; Zákravský, P. Reproductive characteristics of neophytes in the Czech Republic: Traits of invasive and noninvasive species. Preslia 2010, 82, 365–390. [Google Scholar]
- Pearson, D.E.; Icasatti, N.; Hierro, J.L.; Bird, B. Are local filters blind to provenance? Ant seed predation suppresses exotic plants more than natives. PLoS ONE 2014, 9, e103824. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Thomas, H.J.; Bjorkman, A.D. Plant traits inform predictions of tundra responses to global change. New Phytol. 2019, 221, 1742–1748. [Google Scholar] [CrossRef] [PubMed]
- Sakschewski, B.; von Bloh, W.; Boit, A.; Rammig, A.; Kattge, J.; Poorter, L.; Peñuelas, J.; Thonicke, K. Leaf and stem economics spectra drive diversity of functional plant traits in a dynamic global vegetation model. Glob. Chang. Biol. 2015, 21, 2711–2725. [Google Scholar] [CrossRef]
- Schumacher, J.; Roscher, C. Differential effects of functional traits on aboveground biomass in semi-natural grasslands. Oikos 2009, 118, 1659–1668. [Google Scholar] [CrossRef]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating species diversity and distribution in the era of Big Data: To what extent can we trust public databases? Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Pearson, D.E.; Ortega, Y.K.; Eren, Ö.; Hierro, J.L. Community assembly theory as a conceptual framework for invasions. Trends Ecol. Evol. 2018, 33, 313–325. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Martin, C.E.; Falasco, M.R.; James, J.J. Variation in resource acquisition and utilization traits between native and invasive perennial forbs. Am. J. Bot. 2008, 95, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Pearson, D.E.; Ortega, Y.K.; Sears, S.J. Darwin’s naturalization hypothesis up-close: Intermountain grassland invaders differ morphologically and phenologically from native community dominants. Biol. Invasions 2012, 14, 901–913. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cleland, E.E. The phenology of plant invasions: A community ecology perspective. Front. Ecol. Environ. 2011, 9, 287–294. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cleland, E.E. Phenological niches and the future of invaded ecosystems with climate change. AoB Plants 2014, 6, plu013. [Google Scholar] [CrossRef] [Green Version]
- Rosbakh, S.; Chalmandrier, L.; Phartyal, S.; Poschlod, P. Inferring community assembly processes from functional seed trait variation along elevation gradient. J. Ecol. 2022, 110, 2374–2387. [Google Scholar] [CrossRef]
- Schamp, B.; Hettenbergerova, E.; Hajek, M. Testing community assembly predictions for nominal and continuous plant traits in species-rich grasslands. Preslia 2011, 83, 329–346. [Google Scholar]
- Rice, K.J.; Gerlach, J.D.; Dyer, A.R.; McKay, J.K. Evolutionary ecology along invasion fronts of the annual grass Aegilops triuncialis. Biol. Invasions 2013, 15, 2531–2545. [Google Scholar] [CrossRef] [Green Version]
- Morin, X.; Chuine, I. Niche breadth, competitive strength and range size of tree species: A trade-off based framework to understand species distributions. Ecol. Lett. 2006, 9, 185–195. [Google Scholar] [CrossRef]
- Murray, B.R.; Thrall, P.H.; Gill, A.G.; Nicotra, A.B. How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales. Austral Ecol. 2002, 27, 291–310. [Google Scholar] [CrossRef]
- Van der Veken, S.; Bellemare, J.; Verheyen, K.; Hermy, M. Life-history traits are correlated to geographical distribution patterns of western European forest herb species. J. Biogeogr. 2007, 34, 1723–1735. [Google Scholar] [CrossRef]
- Lockwood, J.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef]
- Cordlandwehr, V.; Meredith, R.L.; Ozinga, W.A.; Bekker, R.M.; van Groenendael, J.M.; Bakker, J.P. Do plant traits retrieved from a database accurately predict on-site measurements? J. Ecol. 2013, 101, 662–670. [Google Scholar] [CrossRef]
- DeMalach, N.; Ron, R.; Kadmon, R. Mechanisms of seed mass variation along resource gradients. Ecol. Lett. 2019, 22, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Soper Gorden, N.L.; Winkler, K.J.; Jahnke, M.R.; Marshall, E.; Horky, J.; Hudelson, C.; Etterson, J.R. Geographic patterns of seed mass are associated with climate factors, but relationships vary between species. Am. J. Bot. 2016, 103, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueggler, W.F.; Stewart, W.L. Grassland and Shrubland Habitat Types of Western Montana; USDA FS General Technical Report, INT-66; USDA: Ogden, UT, USA, 1980.
- Pearson, D.E.; Ortega, Y.K.; Eren, O.; Hierro, J.L. Quantifying “apparent” impact and distinguishing impact from invasiveness in multispecies plant invasions. Ecol. Appl. 2016, 26, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W.S. ImageJ; National Institutes of Health: Bethesda, MD, USA, 2018. Available online: http://imagej.nih.gov/ij/ (accessed on 15 January 2021).
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database–enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royal Botanical Gardens KEW. Seed Information Database (SID). Version 7.1; Royal Botanic Gardens Kew: Richmond, UK, 2008. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT User’s Guide, Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuthill, J.E.; Ortega, Y.K.; Pearson, D.E. Seed Size, Seed Dispersal Traits, and Plant Dispersion Patterns for Native and Introduced Grassland Plants. Plants 2023, 12, 1032. https://doi.org/10.3390/plants12051032
Tuthill JE, Ortega YK, Pearson DE. Seed Size, Seed Dispersal Traits, and Plant Dispersion Patterns for Native and Introduced Grassland Plants. Plants. 2023; 12(5):1032. https://doi.org/10.3390/plants12051032
Chicago/Turabian StyleTuthill, Jane E., Yvette K. Ortega, and Dean E. Pearson. 2023. "Seed Size, Seed Dispersal Traits, and Plant Dispersion Patterns for Native and Introduced Grassland Plants" Plants 12, no. 5: 1032. https://doi.org/10.3390/plants12051032