
Effects of Geographical and Climatic Factors on the Intrinsic Water Use Efficiency of Tropical Plants: Evidence from Leaf 13C
Abstract
:1. Introduction
2. Results
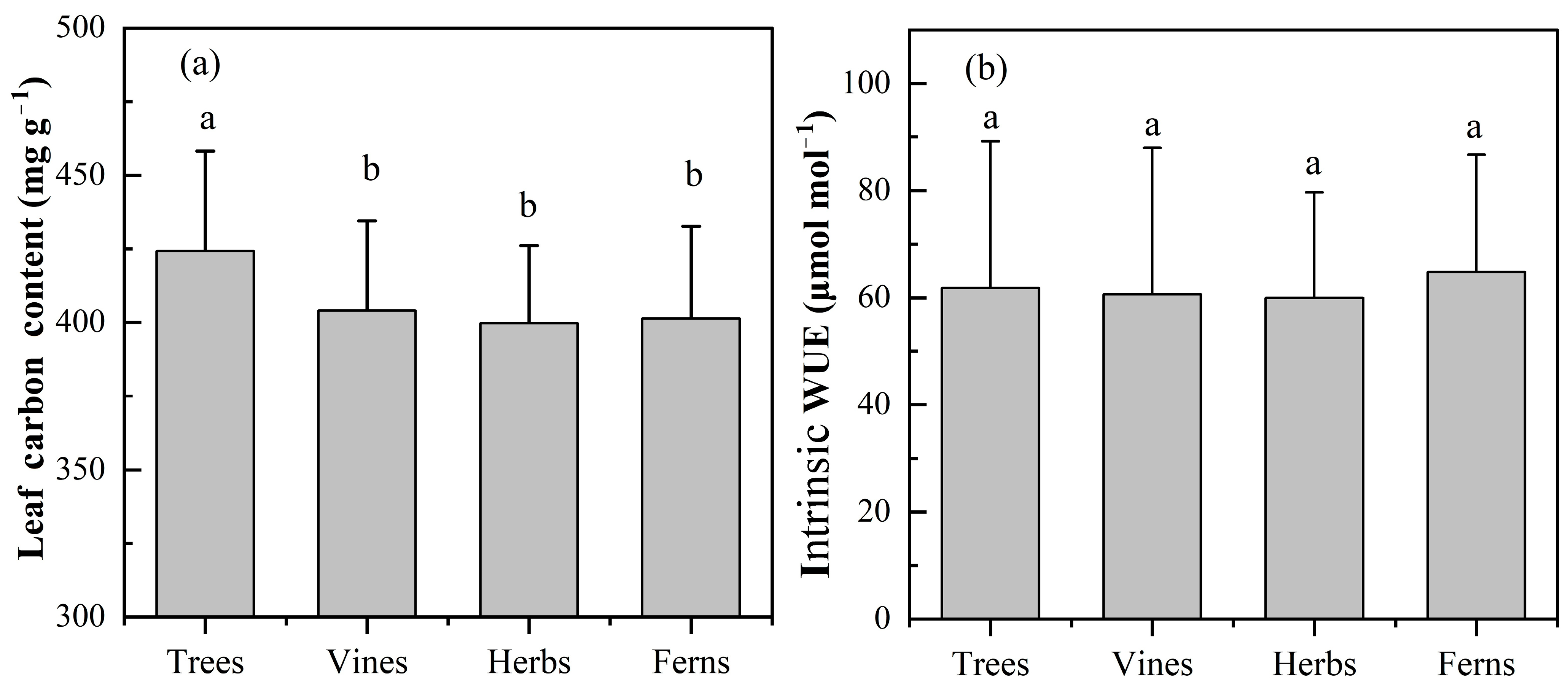
2.1. Characteristics of LCC and Intrinsic WUE
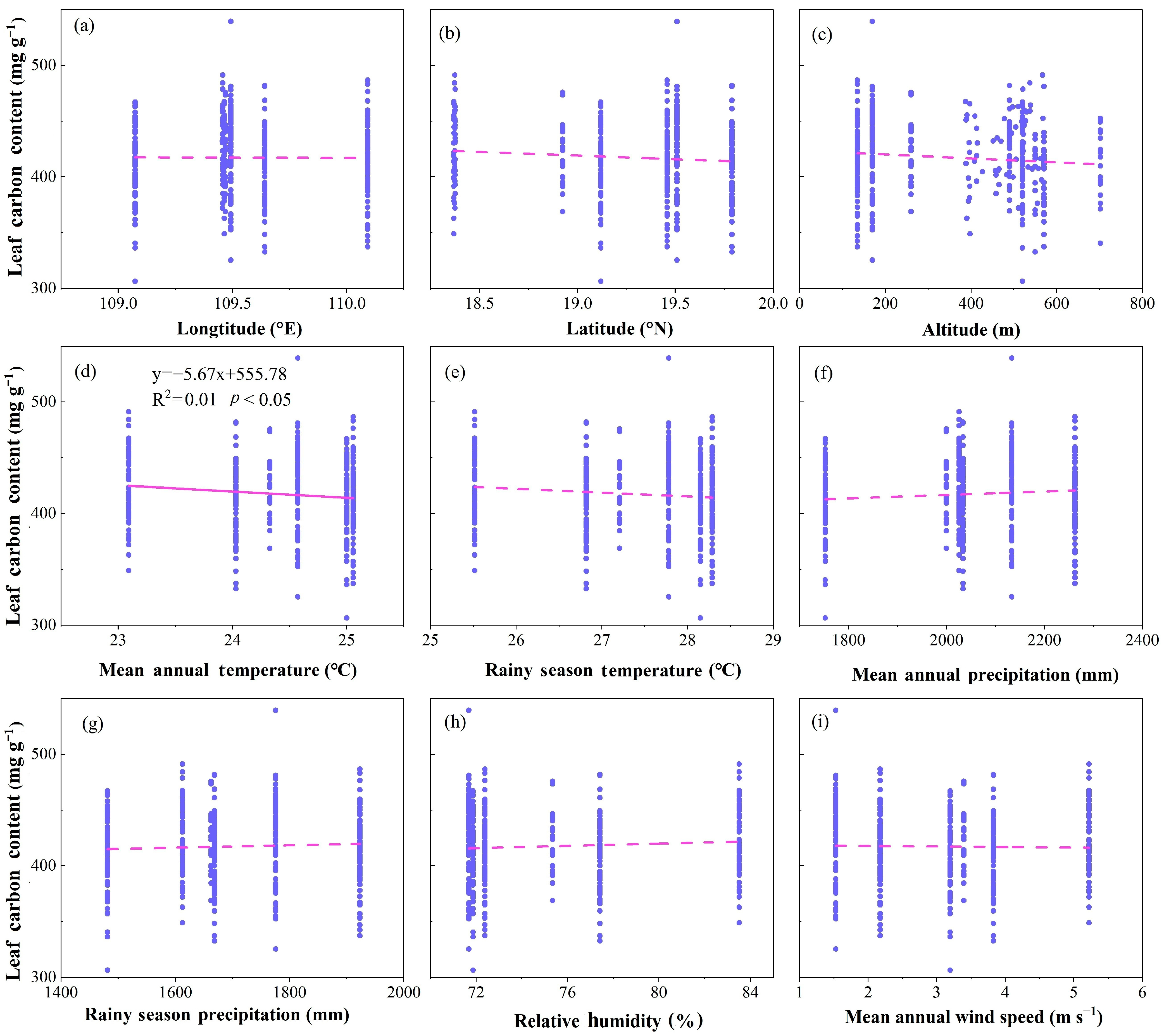
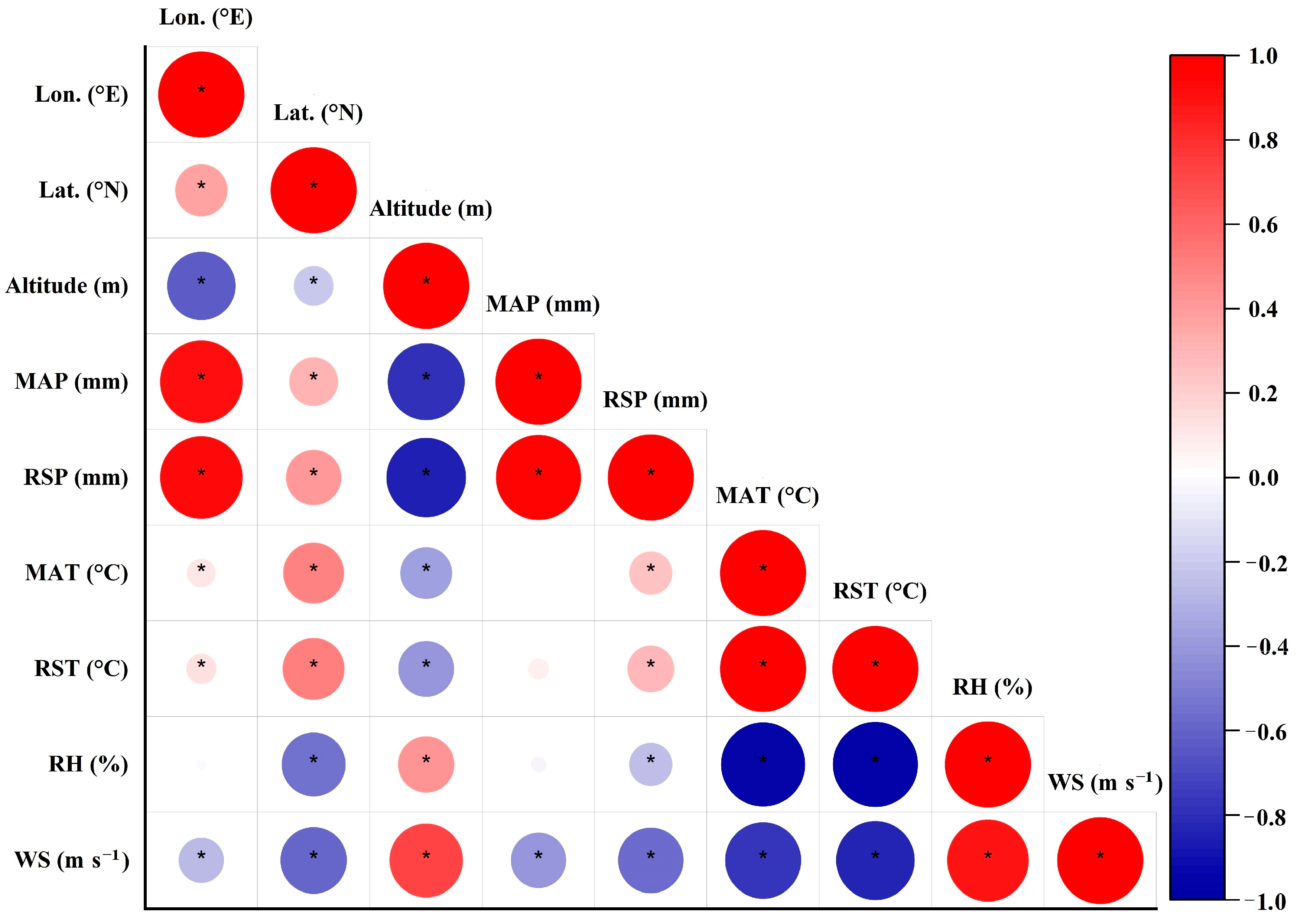
2.2. Relationships among LCC, δ13C and Geographical and Climatic Factors
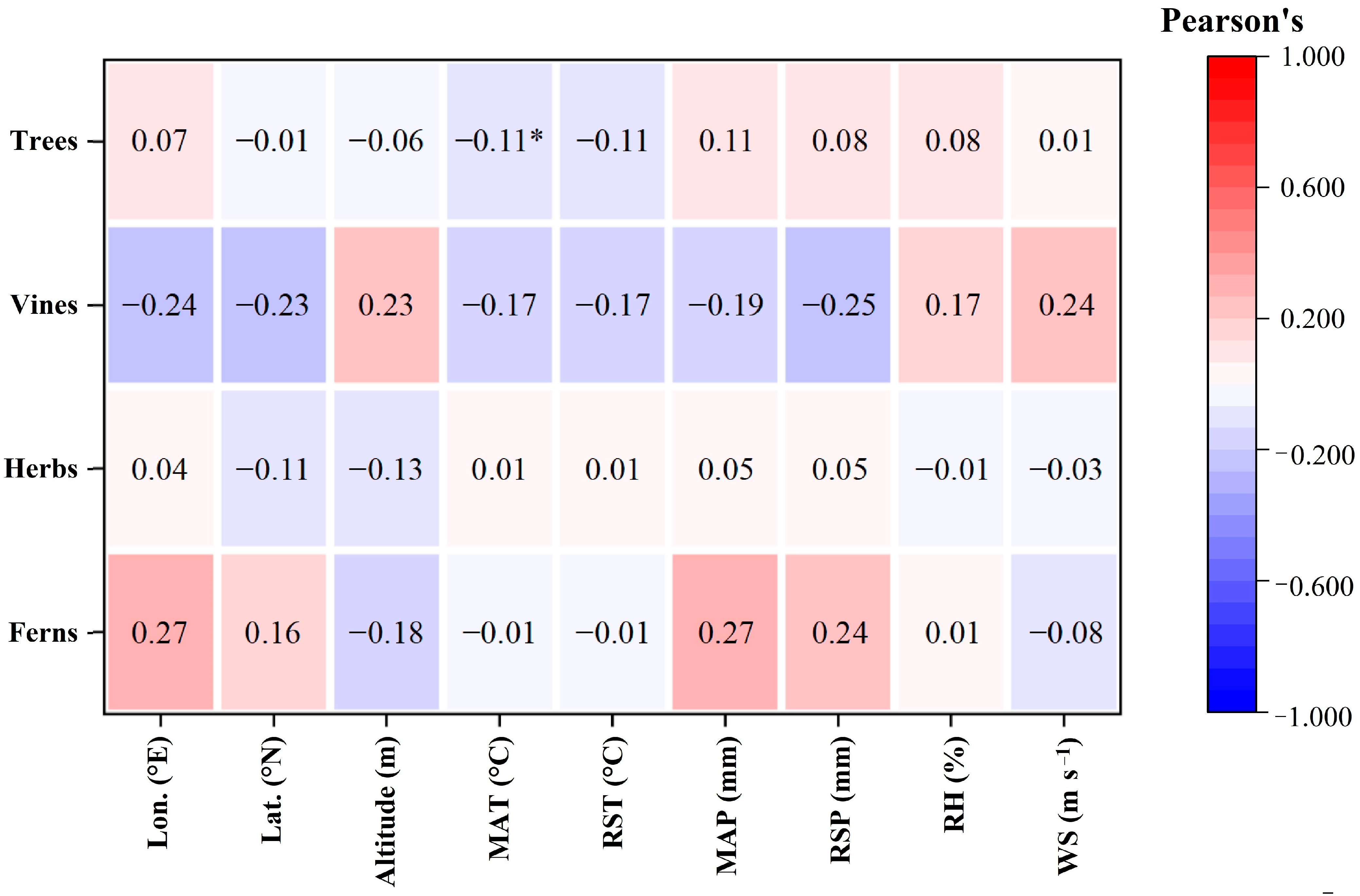
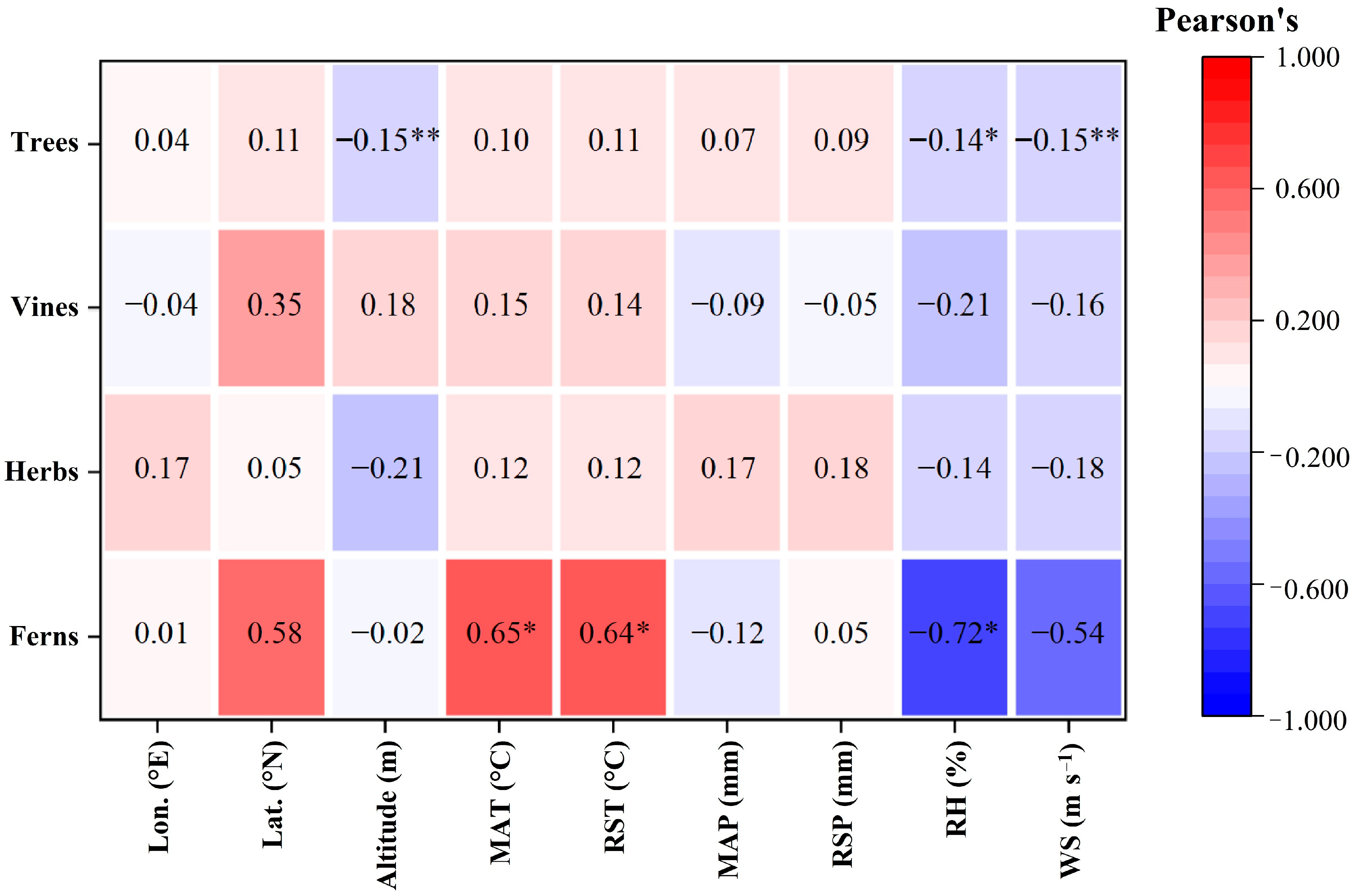
2.3. Relationships among LCC, Intrinsic WUE and Geographical and Climatic Factors of Different Plant Functional Types
3. Discussion
3.1. Characteristics of LCC and Its Relationship with Geographical and Climatic Factors in the Study Area
3.2. Characteristics of LCC and Its Relationship with Geographical and Climatic Factors in Different Functional Types of Plants
3.3. Characteristics of Intrinsic WUE and Its Relationship with Geographical and Climatic Factors in the Study Area
3.4. Characteristics of Leaf Intrinsic WUE and Its Relationship with Geographical and Climatic Factors in Different Plant Functional Types
4. Materials and Methods
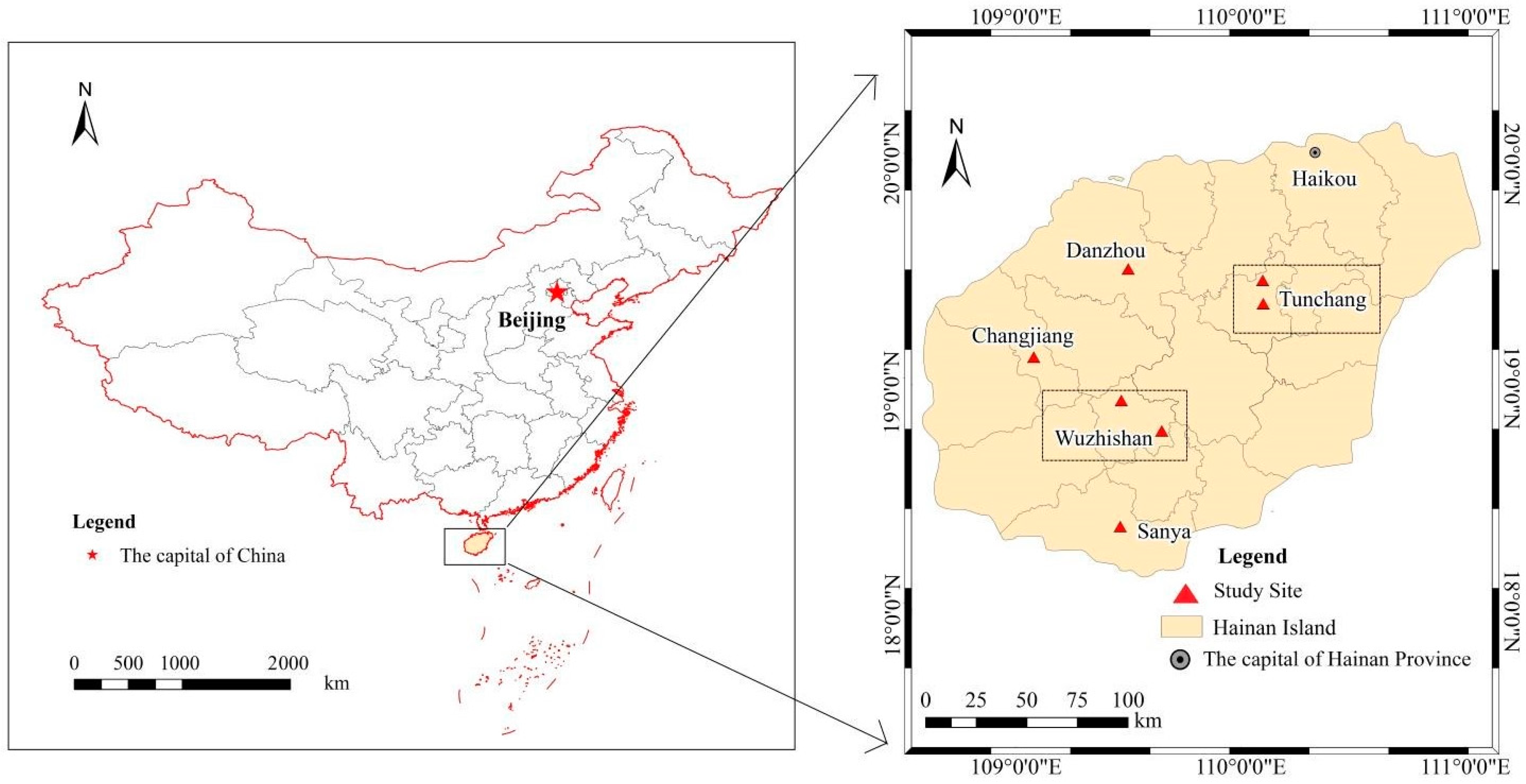
4.1. Study Site Description
4.2. Sample Collection and Analysis
4.3. Calculation of Intrinsic WUE
4.4. Acquisition of Climate Data
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Longitude (°E) | Latitude (°N) | Altitude (m) | MAT (℃) | MAP (mm) |
|---|---|---|---|---|---|
| Danzhou | 109°29′ | 19°30′ | 170.20 | 23.86 | 1934.99 |
| Sanya | 109°27′ | 18°22′ | 6.00 | 25.33 | 1918.85 |
| Haikou | 110°15′ | 20°00′ | 64.70 | 24.08 | 1807.25 |
| Dongfang | 108°37′ | 19°06′ | 8.80 | 25.15 | 1255.55 |
| Baisha | 109°26′ | 19°14′ | 219.30 | 22.29 | 1928.77 |
| Qiongzhong | 109°50′ | 19°02′ | 253.00 | 21.98 | 2134.85 |
| Qionghai | 110°28′ | 19°14′ | 25.20 | 24.64 | 2066.99 |
| Lingshui | 110°02′ | 18°33′ | 13.90 | 23.87 | 2022.10 |
References
- Rahman, M.; Islam, M.; Gebrekirstos, A.; Bräuning, A. Trends in tree growth and intrinsic water-use efficiency in the tropics under elevated CO2 and climate change. Trees 2019, 33, 623–640. [Google Scholar] [CrossRef]
- Basso, L.; Crotwell, A.; Dolman, H.; Gatti, L.; Gerbig, C.; Griffith, D.; Hall, B.; Jordan, A.; Krummel, P.; Leuenberger, M.; et al. The State of Greenhouse Gases in the Atmosphere Based on Global Observations through 2020. World Meteorol. Organ. 2021, 17, 1–6. Available online: https://library.wmo.int/doc_num.php?explnum_id=10904 (accessed on 20 December 2022).
- Mokria, M.; Gebrekirstos, A.; Abiyu, A.; Noordwijk, M.V.; Bräuning, A. Multi-century tree-ring precipitation record reveals increasing frequency of extreme dry events in the upper Blue Nile River catchment. Glob. Chang. Biol. 2017, 23, 5436–5454. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, X.; Cui, X.; Han, Y.; Wang, G.; Li, J. Evaluating the response of δ13C in Haloxylon ammodendron, a dominant C4 species in Asian desert ecosystems, to water and nitrogen addition as well as the availability of its δ13C as an indicator of water use efficiency. Biogeosciences 2021, 18, 2859–2870. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 1998; pp. 51–57. [Google Scholar] [CrossRef]
- Camarero, J.J.; Colangelo, M.; Gazol, A.; Azorín-Molina, C. Drought and cold spells trigger dieback of temperate oak and beech forests in northern Spain. Dendrochronologia 2021, 66, 125812. [Google Scholar] [CrossRef]
- Hussain, M.I.; El-Keblawy, A.; Tsombou, F.M. Leaf age, canopy position, and habitat affect the carbon isotope discrimination and water-use efficiency in three C3 leguminous prosopis species from a hyper-Arid climate. Plants 2019, 8, 402. [Google Scholar] [CrossRef] [Green Version]
- Pennington, R.E.; Tischler, C.R.; Johnson, H.B.; Polley, H.W. Genetic variation for carbon isotope composition in honey mesquite (Prosopis glandulosa). Tree Physiol. 1999, 19, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Rangel, A.; Ávila-Lovera, E.; Guzman, M.E.D.; Torres, L.; Haro, R. Evaluation of leaf carbon isotopes and functional traits in avocado reveals water-use efficient cultivars. Agric. Ecosyst. Environ. 2018, 263, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Maier, C.A.; Burley, J.; Cook, R.; Ghezehei, S.B.; Hazel, D.W.; Nichols, E.G. Tree water use, water use efficiency, and carbon isotope discrimination in relation to growth potential in populus deltoides and hybrids under field conditions. Forests 2019, 10, 993. [Google Scholar] [CrossRef] [Green Version]
- Vantyghem, M.; Merckx, R.; Stevens, B.; Hood-Nowotny, R.; Swennen, R.; Dercon, G. The potential of stable carbon isotope ratios and leaf temperature as proxies for drought stress in banana under field conditions. Agric. Water Manag. 2022, 260, 107247. [Google Scholar] [CrossRef]
- Cano, F.J.; López, R.; Warren, C.R. Implications of the mesophyll conductance to CO2 for photosynthesis and water use efficiency during long-term water stress and recovery in two contrasting Eucalyptus species. Plant Cell Environ. 2014, 37, 2470–2490. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D. On the nature of carbon isotope discrimination in C4 species. Aust. J. Plant Physiol. 1983, 10, 205–226. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Hubick, K.T.; Condon, A.G.; Richards, R.A. Carbon isotope discrimination and plant water-use efficiency. Stable Isot. Ecol. Res. 1989, 68, 21–40. [Google Scholar] [CrossRef]
- Marshall, J.D.; Zhang, J. Carbon isotope discrimination and water-use efficiency in native plants of the north-central Rockies. Ecology 1994, 75, 1887–1895. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Funct. Plant Biol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- McCarth, H.R.; Patak, D.E.; Jeneret, G.D. Plant water-use efficiency as a metric of urban ecosystem services. Ecol. Appl. 2011, 21, 3115–3127. [Google Scholar] [CrossRef]
- Chaves, M.M.; Osório, J.; Pereira, J.S. Water use efficiency and photosynthesis. In Water Use Efficiency in Plant Biology; Blackwell: Oxford, UK, 2004; pp. 51–62. [Google Scholar]
- Driscoll, A.W.; Bitter, N.Q.; Sandquist, D.R.; Ehleringer, J.R. Multidecadal records of intrinsic water-use efficiency in the desert shrub Encelia farinose reveal strong responses to climate change. Proc. Natl. Acad. Sci. USA 2021, 117, 18161–18168. [Google Scholar] [CrossRef]
- Kørup, K.; Lǣrke, P.E.; Baadsgaard, H.; Andersen, M.N.; Kristensen, K.; Münnich, C.; Didion, T.; Jensen, E.S.; Mårtensson, L.; Jørgensen, U. Biomass production and water use efficiency in perennial grasses during and after drought stress. Glob. Chang. Biol. Bioenergy 2017, 10, 12–27. [Google Scholar] [CrossRef]
- Mathias, J.M.; Thomas, R.B. Global tree intrinsic water use efficiency is enhanced by increased atmospheric CO2 and modulated by climate and plant functional types. Proc. Natl. Acad. Sci. USA 2021, 118, e2014286118. [Google Scholar] [CrossRef]
- Olson, E.J.; Dodd, J.P.; River, M.A. Prosopis sp. tree-ring oxygen and carbon isotope record of regional-scale hydroclimate variability during the last 9500 years in the Atacama Desert. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 238, 109408. [Google Scholar] [CrossRef]
- Guo, G.; Fang, K.; Li, J.; Linderholm, H.W.; Li, D.; Zhou, F.; Dong, Z.; Li, Y.; Wang, L. Increasing intrinsic water-use efficiency over the past 160 years does not stimulate tree growth in southeastern China. Clim. Res. 2018, 76, 115–130. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, X.; Jiang, H.; Qiao, S.; Guan, M.; Huang, Y.; Gong, R. Widespread decline in winds promoted the growth of vegetation. Sci. Total Environ. 2020, 825, 153682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Marshall, J.D.; Jaquish, B.C. Genetic differentiation in carbon isotope discrimination and gas exchange in Pseudotsuga menziesii. Oecologia 1993, 93, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Mumbanza, F.M.; Bauters, M.; Meunier, F.; Boeckx, P.; Cernusak, L.A.; De Deurwaerder, H.P.T.; Demol, M.; Meeussen, C.; Sercu, B.; Verryckt, L.T.; et al. Lianas and trees exhibit divergent intrinsic water-use efficiency along elevational gradients in South American and African tropical forests. Glob. Ecol. Biogeogr. 2021, 30, 2259–2272. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, J.; Terradas, J. Variability of plant nitrogen and water use in a 100-m transect of a subdesertic depression of the Ebro valley (Spain) characterized by leaf δ13C and δ15N. Acta Oecon. 1999, 20, 119–123. [Google Scholar] [CrossRef]
- Marcott, S.A.; Shakun, J.D.; Clark, P.U.; Mix, A.C. A reconstruction of regional and global temperature for the past 11,300 years. Science 2013, 339, 1198–1201. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- He, J.; Fang, J.; Wang, Z.; Guo, D.; Flynn, D.F.B.; Geng, Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia 2006, 149, 115–122. [Google Scholar] [CrossRef]
- Tong, R.; Zhou, B.; Jiang, L.; Ge, X.; Cao, Y. Spatial patterns of leaf carbon, nitrogen, and phosphorus stoichiometry and nutrient resorption in Chinese fir across subtropical China. Catena 2021, 201, 105221. [Google Scholar] [CrossRef]
- Zhao, N.; He, N.; Wang, Q.; Zhang, X.; Wang, R.; Xu, Z.; Yu, G. The altitudinal patterns of leaf C:N:P stoichiometry are regulated by plant growth form, climate and soil on Changbai Mountain, China. PLoS ONE 2014, 9, e95196. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2011; pp. 97–112. [Google Scholar] [CrossRef]
- Millard, P.; Sommerkorn, M.; Grelet, G. Environmental change and carbon limitation in trees: A biochemical, ecophysiological and ecosystem appraisal. New Phytol. 2007, 17, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Hoch, G.; Körner, C. Global patterns of mobile carbon stores in trees at the high-elevation tree line. Glob. Ecol. Biogeogr. 2012, 21, 861–871. [Google Scholar] [CrossRef]
- Chaitanya, K.V.; Sundar, D.; Reddy, A.R. Mulberry leaf metabolism under high temperature stress. Biol. Plant 2001, 44, 379–384. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.V.V.; Boyle, D.L.; Schapaugh, W.T. High-temperature stress and soybean leaves: Leaf anatomy and photosynthesis. Crop Sci. 2011, 51, 2125–2131. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, B.; Wang, B.; Zhang, G.; Zhang, W.; Zhang, B.; Chen, Z.; Chen, T.; Liu, G. Leaf elemental stoichiometry of Tamarix Lour. species in relation to geographic, climatic, soil, and genetic components in China. Ecol. Eng. 2017, 106, 448–457. [Google Scholar] [CrossRef]
- Huang, C.; Chu, C.; Hsieh, C.; Palmroth, S.; Katul, G. Wind-induced leaf transpiration. Adv. Water Resour. 2015, 86, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Li, X.; Wang, Q.; Tian, J.; Tang, X.; Tang, Z.; Xie, Z.; He, N.; Yu, G. Spatial patterns and environmental factors influencing leaf carbon content in the forests and shrublands of China. J. Geogr. Sci. 2018, 28, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Chen, H. Global trends in senesced-leaf nitrogen and phosphorus. Glob. Ecol. Biogeorgr. 2009, 18, 532–542. [Google Scholar] [CrossRef]
- Chapin, F.; Shaver, G. Difference in carbon and nutrient fractions among arctic growth forms. Oecologia 1988, 77, 506–514. [Google Scholar] [CrossRef]
- Umair, M.; Sun, N.; Du, H.; Chen, K.; Tao, H.; Yuan, J.; Abbasi, A.M.; Liu, C. Differential stoichiometric responses of shrubs and grasses to increased precipitation in a degraded karst ecosystem in Southwestern China. Sci. Total Environ. 2020, 700, 134421. [Google Scholar] [CrossRef]
- Cai, Z.; Schnitzer, S.; Bongers, F. Seasonal differences in leaf-level physiology give lianas a competitive advantage over trees in a tropical seasonal forest. Oecologia 2009, 161, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asner, G.; Martin, R. Contrasting leaf chemical traits in tropical lianas and trees: Implications for future forest composition. Ecol. Lett. 2012, 15, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.; Wright, S.; Wurzburger, N. Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees. Oecologia 2016, 180, 1037–1047. [Google Scholar] [CrossRef]
- Pasquini, S.; Wright, S.; Santiago, L. Lianas always outperform tree seedlings regardless of soil nutrients: Results from a long-term fertilization experiment. Ecology 2015, 96, 1866–1876. [Google Scholar] [CrossRef] [Green Version]
- Doi, M.; Wada, M.; Shimazaki, K. The fern Adiantum capillus-veneris lacks stomatal responses to blue light. Plant Cell Physiol. 2006, 47, 748–755. [Google Scholar] [CrossRef]
- Li, H.; Wei, M.; Dong, L.; Hu, W.; Xiong, J.; Sun, Y.; Sun, Y.; Yao, S.; Gong, H.; Zhang, Y.; et al. Leaf and ecosystem water use efficiencies differ in their global-scale patterns and drivers. Agric. For. Meteorol. 2022, 319, 108919. [Google Scholar] [CrossRef]
- Ren, M.; Liu, Y.; Li, Q.; Song, H.; Cai, Q.; Sun, C. Responses of tree growth and intrinsic water use efficiency to environmental factors in Central and Northern China in the context of global warming. Forests 2022, 13, 1209. [Google Scholar] [CrossRef]
- Ma, W.; Tcherkez, G.; Wang, X.; Schaufele, R.; Schnyder, H.; Yang, Y.; Gong, X. Accounting for mesophyll conductance substantially improves 13C-based estimates of intrinsic water-use efficiency. New Phytol. 2021, 229, 1326–1338. [Google Scholar] [CrossRef]
- Doughty, C.E.; Goulden, M.L. Are tropical forests near a high temperature threshold? J. Geophys. Res. 2008, 113, G00B07. [Google Scholar] [CrossRef] [Green Version]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef] [Green Version]
- Onoda, Y.; Anten, P.R.N. Challenges to understand plant responses to wind. Plant Signal Behav. 2011, 6, 1057–1059. [Google Scholar] [CrossRef] [Green Version]
- López, R.; Cano, F.J.; Rodríguez-Calcerrada, J.; Sangüesa-Barreda, G.; Gazol, A.; Camarero, J.J.; Rozenberg, P.; Gil, L. Tree-ring density and carbon isotope composition are early-warning signals of drought-induced mortality in the drought tolerant Canary Island pine. Agric. For. Meteorol. 2021, 310, 108634. [Google Scholar] [CrossRef]






| F | L | A | F × L | F × A | L × A | |
|---|---|---|---|---|---|---|
| LCC | 3.78 * | 0.37 | 1.08 | 0.49 | 0.49 | 7.91 ** |
| Intrinsic WUE | 3.07 * | 6.74 *** | 4.42 ** | 1.73 | 0.91 | 2.56 |
| Methods | Factors Included in the Models | R2 | p | |
|---|---|---|---|---|
| All sample | Input | All factors | 0.083 | <0.001 |
| Stepwise | WS | 0.024 | 0.001 | |
| Woody plants | Input | All factors | 0.081 | <0.001 |
| Stepwise | Altitude | 0.024 | 0.006 | |
| Ferns | Input | All factors | 0.810 | 0.048 |
| Stepwise | Altitude, RH | 0.766 | 0.006 |
| Study Site | Longitude (°E) | Latitude (°N) | Altitude (m) | MAT (℃) | MAP (mm) | Species |
|---|---|---|---|---|---|---|
| Changjiang | 109°04′ | 19°07′ | 520–800 | 17.35 | 1563.12 | 79 |
| Danzhou | 109°29′ | 19°30′ | 137 | 23.86 | 1934.99 | 86 |
| Sanya | 109°27′ | 18°22′ | 387–568 | 23.47 | 1918.85 | 59 |
| Tunchang | 110°06′ | 19°21′ | 110–160 | 23.13 | 2105.15 | 84 |
| 110°05′ | 19°27′ | |||||
| Wuzhishan | 109°38′ | 18°47′ | 490–520 | 22.80 | 2080.95 | 106 |
| 109°28′ | 18°55′ | 260 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, X.; Wu, B.; Wang, J.; Wang, G.; Chen, Z.; Liang, Y.; Liu, J.; Wang, H. Effects of Geographical and Climatic Factors on the Intrinsic Water Use Efficiency of Tropical Plants: Evidence from Leaf 13C. Plants 2023, 12, 951. https://doi.org/10.3390/plants12040951
Lin X, Wu B, Wang J, Wang G, Chen Z, Liang Y, Liu J, Wang H. Effects of Geographical and Climatic Factors on the Intrinsic Water Use Efficiency of Tropical Plants: Evidence from Leaf 13C. Plants. 2023; 12(4):951. https://doi.org/10.3390/plants12040951
Chicago/Turabian StyleLin, Xiaoyan, Bingsun Wu, Jingjing Wang, Guoan Wang, Zixun Chen, Yongyi Liang, Jiexi Liu, and Hao Wang. 2023. "Effects of Geographical and Climatic Factors on the Intrinsic Water Use Efficiency of Tropical Plants: Evidence from Leaf 13C" Plants 12, no. 4: 951. https://doi.org/10.3390/plants12040951