
High Prevalence of Clonal Reproduction and Low Genetic Diversity in Scutellaria floridana, a Federally Threatened Florida-Endemic Mint

Abstract
:1. Introduction
2. Results
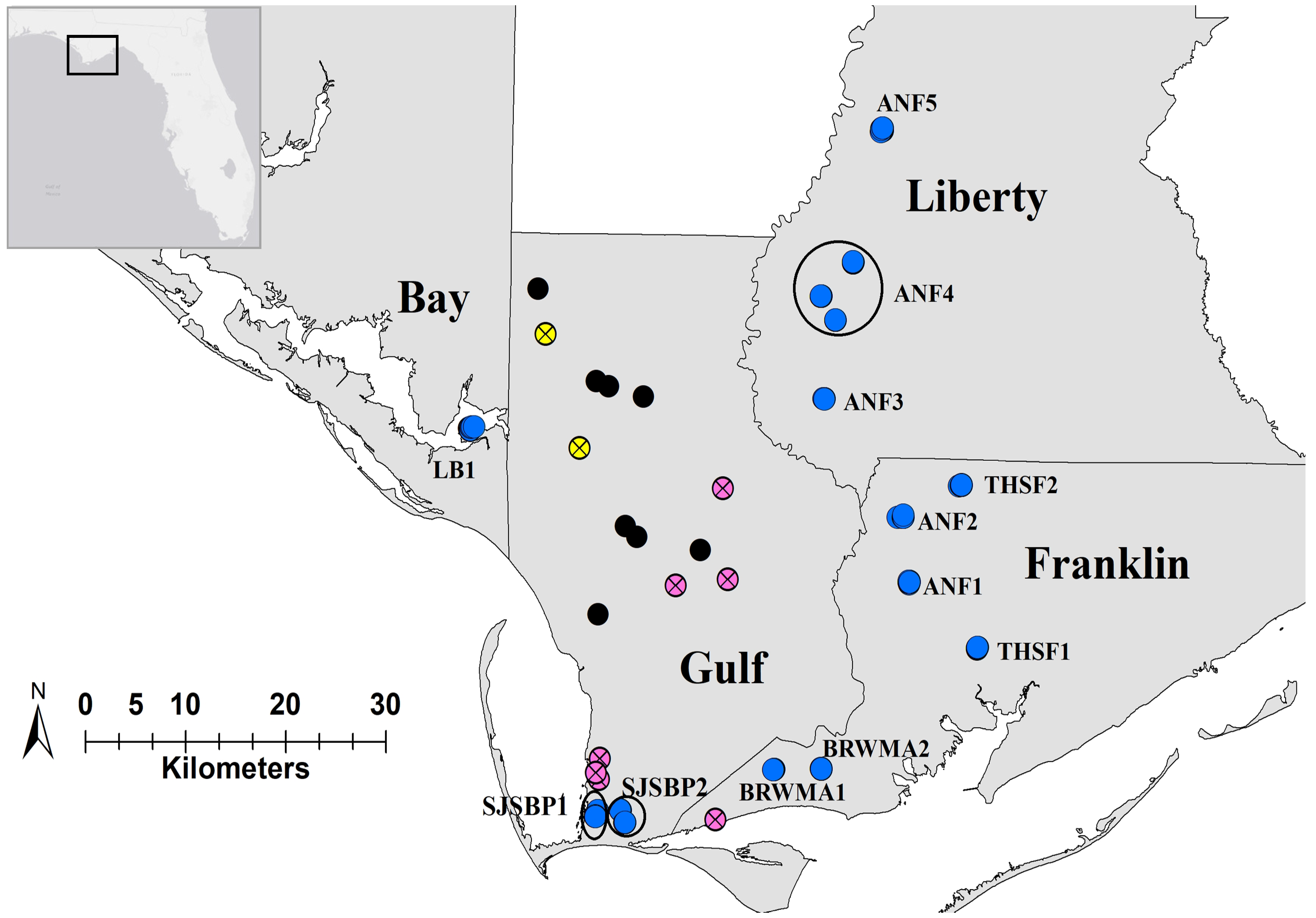
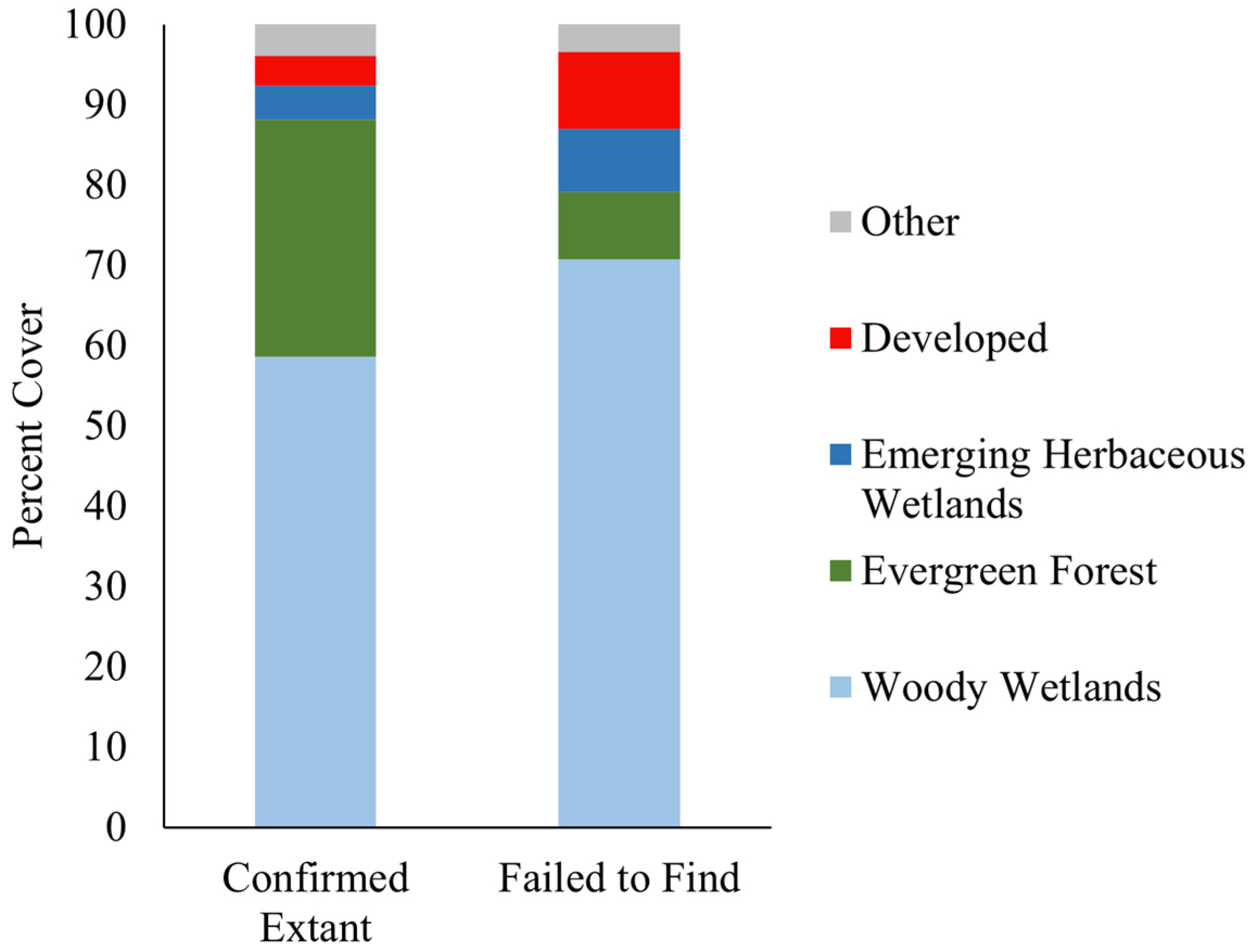
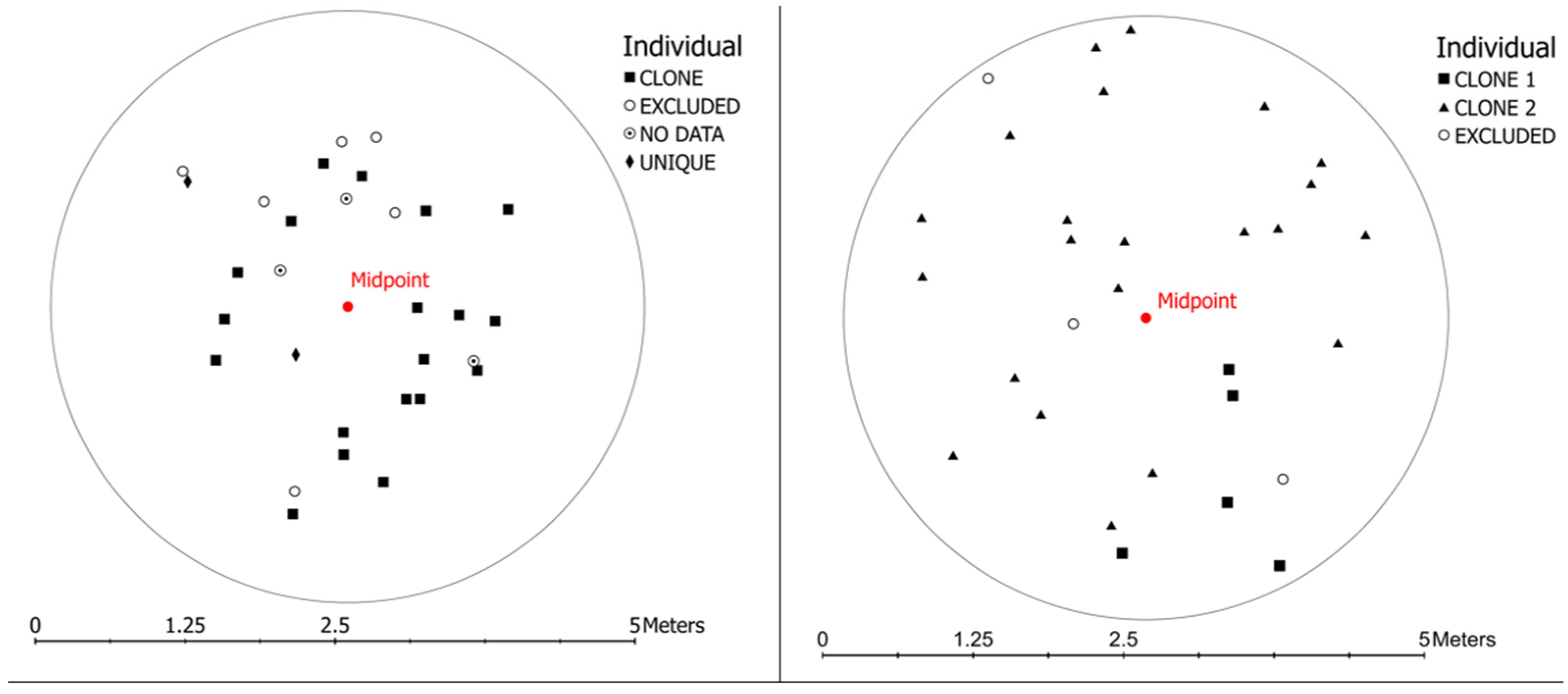
2.1. Spatial Analysis
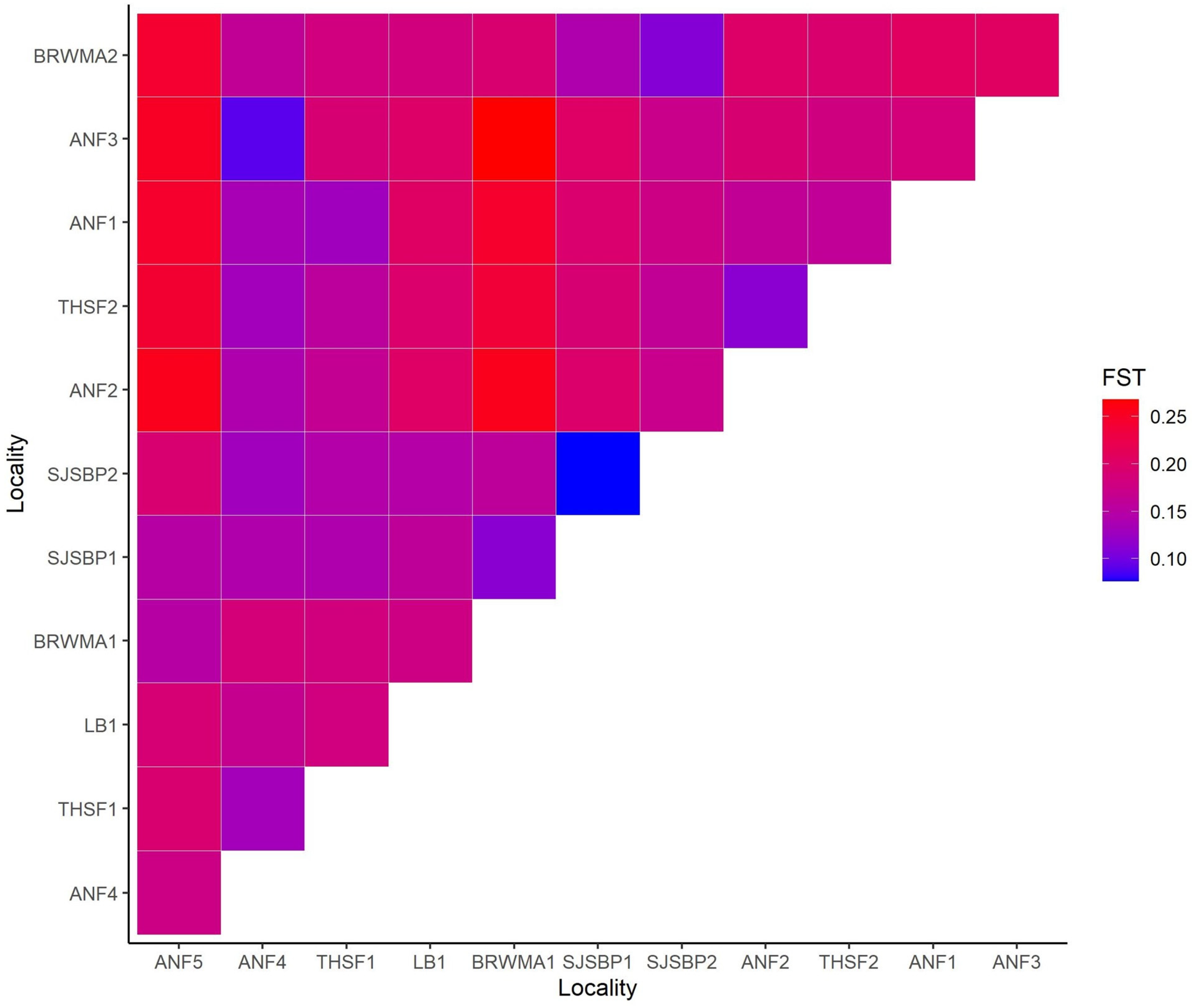
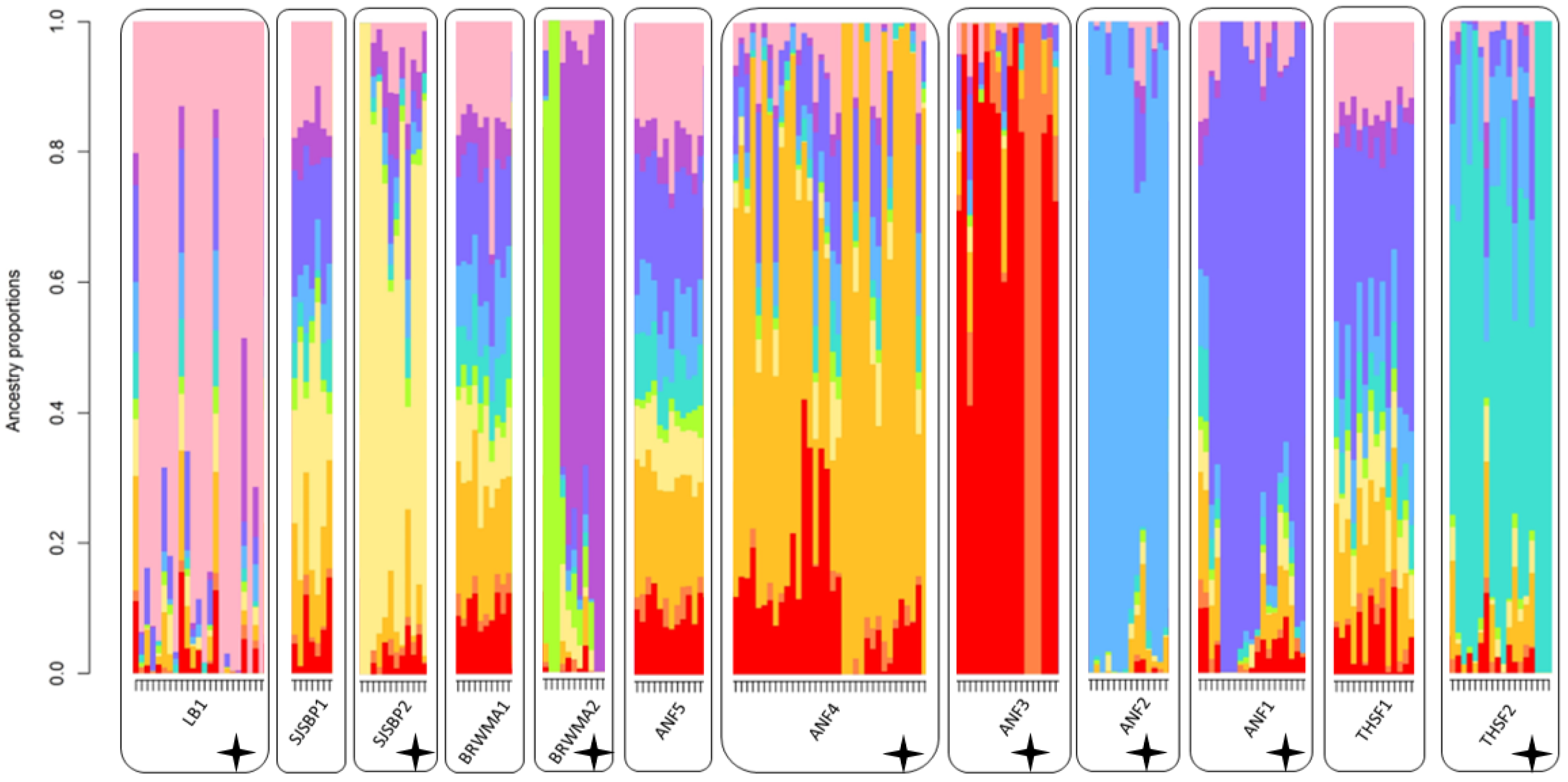
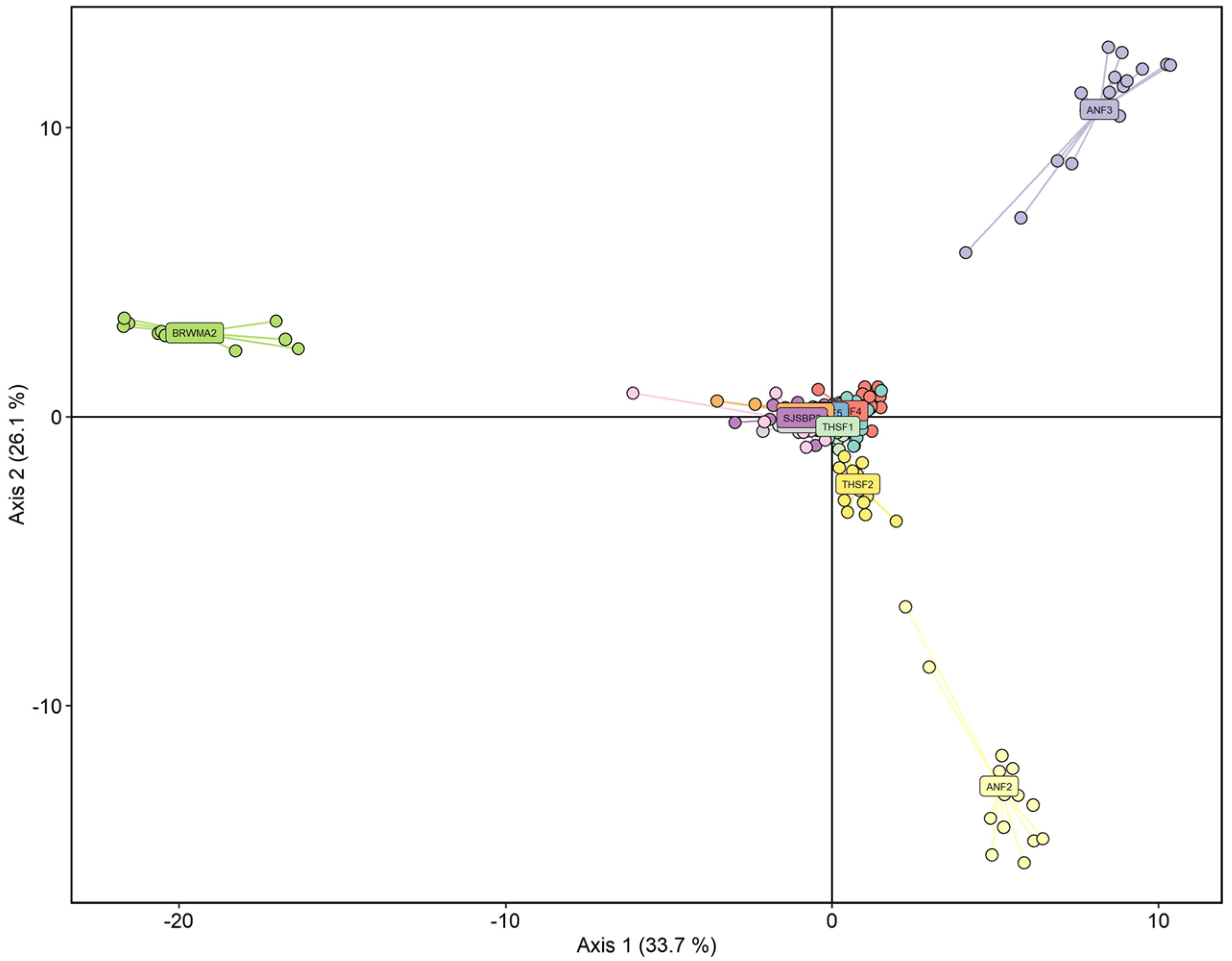
2.2. Population Structure and Diversity
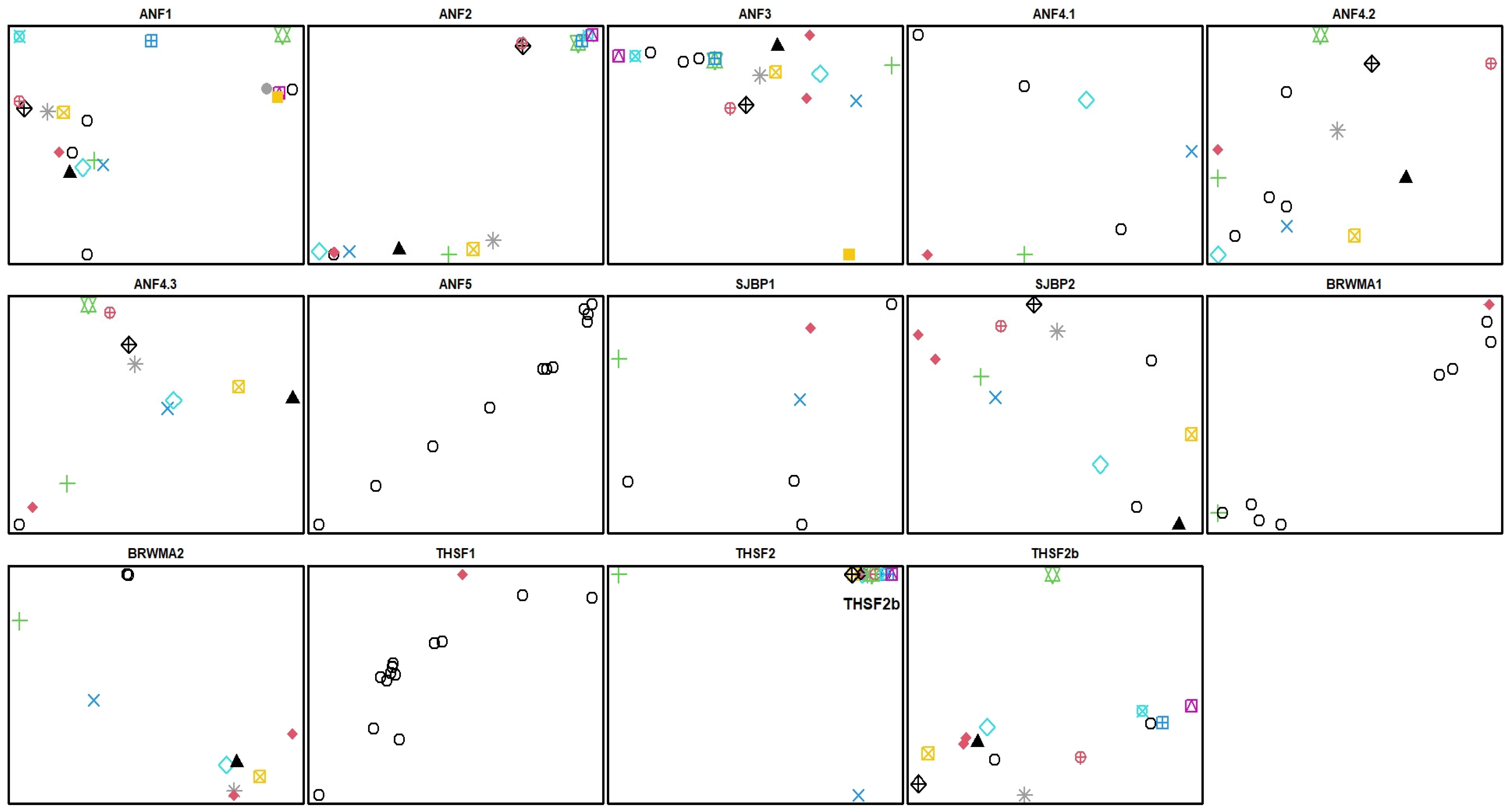
2.3. Clonality
3. Discussion
Implications for Conservation and Management
4. Materials and Methods
4.1. Study Area
4.2. Tissue Collection and Storage
4.3. DNA Isolation and Sequencing
4.4. Spatial Analysis
4.5. Genetic Diversity Assessment
4.6. Clonal Assignment and Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, R.; Inouye, B.; Johnson, M.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef]
- Laikre, L. Genetic diversity is overlooked in international policy implementation. Conserv. Genet. 2010, 11, 349–354. [Google Scholar] [CrossRef]
- Cook, C.N.; Sgró, C.M. Aligning science and policy to achieve evolutionarily enlightened conservation. Conserv. Biol. 2016, 31, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Ralls, K.; Ballou, J.D.; Dudash, M.R.; Eldridge, M.D.B.; Fenster, C.B.; Lacy, R.C.; Sunnucks, P.; Frankham, R. Call for a paradigm shift in the genetic management of fragmented populations. Conserv. Lett. 2018, 11, e12412. [Google Scholar] [CrossRef]
- Holderegger, R.; Balkenhol, N.; Bolliger, J.; Engler, J.O.; Gugerli, F.; Hochkirch, A.; Nowak, C.; Segelbacher, G.; Widmer, A.; Zachos, F.E. Conservation genetics: Linking science with practice. Mol. Ecol. 2019, 28, 3848–3856. [Google Scholar] [CrossRef] [Green Version]
- Willi, Y.; Kristensen, T.N.; Sgro, C.M.; Weeks, A.R.; Orsted, M.; Hoffmann, A.A. Conservation genetics as a management tool: The five best-supported paradigms to assist the management of threatened species. Proc. Natl. Acad. Sci. USA 2022, 119, e2105076119. [Google Scholar] [CrossRef]
- Frankham, R.; Bradshaw, C.J.A.; Brook, B.W. Genetics in conservation management: Revised guidelines for the 50/500 rules, Red List criteria and population viability analyses. Biol. Conserv. 2014, 170, 56–63. [Google Scholar] [CrossRef]
- Shafer, A.B.A.; Wolf, J.B.W.; Alves, P.C.; Bergstrom, L.; Bruford, M.W.; Brannstrom, I.; Colling, G.; Dalen, L.; Meester, L.D.; Ekblom, R.; et al. Genomics and the challenging translation into conservation practice. Trends Ecol. Evol. 2015, 30, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Jump, A.S.; Penuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Kramer, J.T.; Havens, K. Plant conservation genetics in a changing world. Trends Plant Sci. 2009, 14, 599–607. [Google Scholar] [CrossRef]
- Bragg, J.G.; Cuneo, P.; Sherieff, A.; Rossetto, M. Optimizing the genetic composition of a translocation population: Incorporating constraints and conflicting objectives. Mol. Ecol. Resour. 2020, 20, 54–65. [Google Scholar] [CrossRef]
- St. Clair, A.B.; Dunwiddie, P.W.; Fant, J.B.; Kaye, T.N.; Kramer, A.T. Mixing source populations increases genetic diversity of restored rare plant populations. Restor. Ecol. 2020, 28, 583–593. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service (USFWS). Scutellaria floridana (Florida skullcap) 5-Year Review: Summary and Evaluation; USFW: Panama City, FL, USA, 2019; p. 19.
- Molano-Flores, B.; Coons, J.; Annis, J.; O’Brien, J.; Feist, M.; Koontz, J.; Maruzak, J.; Menglekoch, J. Seed Ecology and Population Genetics Studies for Pinguicula ionantha (Godfrey’s Butterwort) and Scutellaria floridana (Florida skullcap); Illinois Natural History Survey: Champaign, IL, USA, 2014; p. 27. [Google Scholar]
- Ye, X.-H.; Yu, F.-H.; Dong, M. A trade-off between guerrilla and phalanx growth forms in Leymus secalinus under different nutrient supplies. Ann. Bot. 2006, 98, 187–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Witte, L.C.; Stocklin, J. Longevity of clonal plants: Why it matters and how to measure it. Ann. Bot. 2010, 106, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Tepedino, V.J. Overestimating population sizes of rare clonal plants. Conserv. Biol. 2012, 26, 945–947. [Google Scholar] [CrossRef] [PubMed]
- Balloux, F.; Lehmann, L.; de Meeus, T. The population genetics of clonal and partially clonal diploids. Genetics 2003, 164, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Evolution in Mendelian populations. Genetics 1930, 16, 97–169. [Google Scholar] [CrossRef] [PubMed]
- U.S. Fish and Wildlife Service. Recovery Plan for Four Plants of the Lower Apalachicola Region, Florida: Euphorbia Telephioides (Telephus Spurge), Macbridea Alba (White Birds-in-a-Nest), Pinguicula ionantha (Godfrey’s Butterwort), and Scutellaria floridana (Florida skullcap); Fish and Wildlife Service: Panama City, FL, USA, 1994; p. 32. Available online: https://ecos.fws.gov/ecp/species/2240 (accessed on 14 March 2022).
- Kyrkjeeide, M.O.; Westergaard, K.B.; Kleven, O.; Evju, M.; Endrestol, A.; Brandrud, M.K.; Stabbetorp, O. Conserving on the edge: Genetic variation and structure in northern populations of the endangered plant Dracocephalum ruyschiana L. (Lamiaceae). Conserv. Genet. 2020, 21, 707–718. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.; Huang, Y.; Zhao, H.; Wu, J.; Qi, Z.; Wei, Y. Conservation genomics of wild red sage (Salvia miltiorrhiza) and its endangered relatives in China: Population structure and interspecific relationships revealed from 2b-RAD data. Front. Genet. 2021, 12, 688323. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Elam, D.R. Population genetic consequences of small population size: Implications for plant conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]
- Aguilar, R.; Quesada, M.; Ashworth, L.; Herrerias-Diego, Y.; Lobo, J.A. Genetic consequences of habitat fragmentation in plant populations: Susceptible signals in plant traits and methodological approaches. Mol. Ecol. 2019, 17, 5177–5188. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-Marin, M.; Dorken, M.E.; Barrett, S.C. The ecological and evolutionary consequences of clonality for plant mating. Annu. Rev. Ecol. Evol. S. 2010, 41, 193–213. [Google Scholar] [CrossRef] [Green Version]
- Delmotte, F.; Leterme, N.; Gauthier, J.P.; Rispe, C.; Simon, J.C. Genetic architecture of sexual and asexual populations of the aphid Rhopalosiphum padi based on allozyme and microsatellite markers. Mol. Ecol. 2002, 11, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Lovet, D.L. Population dynamics and local specialization in a clonal perennial (Ranunculus repens) 1. The dynamics of ramets in contrasting habitats. J. Ecol. 1981, 69, 743–755. [Google Scholar] [CrossRef]
- Olmstead, R.G. Biological and historical factors influencing genetic diversity in the Scutellaria angustifolia complex (Labiatae). Evolution 1990, 44, 54–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M. Cleistogamy in Scutellaria indica (Labiatae): Effective mating system and population genetic structure. Mol. Ecol. 1999, 88, 1285–1295. [Google Scholar] [CrossRef]
- Maguire, A.J.; Menges, E.S. Post-fire growth strategies of resprouting Florida scrub vegetation. Fire Ecol. 2011, 7, 12–25. [Google Scholar] [CrossRef]
- Brewer, J.S.; Platt, W. Effects of fire season and soil fertility on clonal growth in a pyrophilic forb, Pityopsis graminifolia (Asteraceae). Am. J. Bot. 1994, 817, 805–814. [Google Scholar] [CrossRef]
- Hartnett, D.C. Effects of Fire on Clonal Growth and Dynamics of Pityopsis graminifolia (Asteraceae). Am. J. Bot. 1987, 74, 1737–1743. [Google Scholar] [CrossRef]
- Slapcinsky, J.L.; Gordon, D.R.; Menges, E.S. Responses of rare plant species to fire in Florida’s pyrogenic communities. Nat. Area. J. 2010, 30, 4–19. [Google Scholar] [CrossRef]
- Ames, G.M.; Anderson, S.M.; Wright, J.P. Multiple environmental drivers structure plant traits at the community level in a pyrogenic ecosystem. Funct. Ecol. 2016, 30, 789–798. [Google Scholar] [CrossRef]
- Negrón-Ortiz, V.; Kaeser, M. Timing and patterns of size, reproduction, and seed germination in Florida endemic Euphorbia telephioides (Euphorbiaceae): Management and conservation implications. Nat. Area. J. 2020, 40, 262–272. [Google Scholar] [CrossRef]
- NOAA National Centers for Environmental Information (NCEI) U.S. Climate Normals 2020: Annual/Seasonal Climate Normals 2006–2020. Available online: https://is.gd/IsUcIp (accessed on 14 March 2022).
- NatureServe. NatureServe Network Biodiversity Location Data; NatureServe: Arlington, VA, USA, 2022. [Google Scholar]
- U.S. Forest Service (USFS). Forest Plan: National Forests in Florida; USFS: Washington, DC, USA, 1999.
- Florida Department of Agriculture and Consumer Services & Florida Forest Service. Ten-Year Land Management Plan for the Tate’s Hell State Forest; Florida Department of Agriculture and Consumer Services & Florida Forest Service: Tallahassee, FL, USA, 2019.
- Florida Fish and Wildlife Conservation Commission. A Management Plan for Box-R Wildlife Management Area; Florida Fish and Wildlife Conservation Commission: Tallahassee, FL, USA, 2016. [Google Scholar]
- Florida Department of Environmental Protection. St. Joseph Bay State Buffer Preserve Management Plan; Florida Department of Environmental Protection: Tallahassee, FL, USA, 2016.
- Bureau of Land Management. Lathrop Bayou. Final Habitat Management Plan Environmental Assessment; Bureau of Land Management: Jackson, MS, USA, 2003.
- Kim, C.S.; Lee, C.H.; Shin, J.S.; Chung, Y.S.; Hyung, N.I. A simple and rapid method for isolation of high-quality DNA from fruit trees and conifers using PVP. Nucleic Acids Res. 1997, 25, 1085–1086. [Google Scholar] [CrossRef] [PubMed]
- Catchen, J.; Hohenlohe, P.; Bassham, S.; Amores, A.; Cresko, W. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Paris, J.R.; Stevens, J.R.; Catchen, J.M. Lost in parameter space: A road map for STACKS. Methods Ecol. Evol. 2017, 8, 1360–1373. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J.; Jombart, T. Hierfstat: Estimation and Tests of Hierarchical F-Statistics. 2022. Available online: https://www.r-project.org (accessed on 1 May 2022).
- Pembleton, L.W.; Cogan, N.O.I.; Forster, J.W. StAMPP: An R package for calculation of genetic differentiation and structure of mixed-ploidy level populations. Mol. Ecol. Res. 2013, 13, 946–952. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Frichot, E.; Francois, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Dorken, M.E.; Eckert, C.G. Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae). J. Ecol. 2001, 89, 339–350. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Population | Sites | N | HE | HO | FIS | Population Divergence | Unique Ancestry | Genetic Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| Bay | LB1 | 1 | 23 | 0.142 | 0.125 | 0.113 | Moderate | Yes | Moderate | |
| Franklin | ANF1 | 1 | 19 | 0.125 | 0.106 | 0.137 | Moderate | Yes | Moderate | |
| Franklin | ANF2 | 1 | 14 | 0.128 | 0.127 | 0.024 | Very High | Yes | High | |
| Franklin | BRWMA1 | 1 | 10 | 0.139 | 0.04 | 0.555 | Moderate | No | Low | |
| Franklin | BRWMA2 | 1 | 11 | 0.136 | 0.139 | −0.01 | Very High | Yes | High | |
| Franklin | THSF1 | 1 | 14 | 0.131 | 0.081 | 0.297 | Moderate | No | Low | |
| Franklin | THSF2 | 2 | 18 | 0.131 | 0.13 | 0.025 | High | Yes | High | |
| Gulf | SJSBP1 | 2 | 7 | 0.145 | 0.090 | 0.268 | Moderate | No | Low | |
| Gulf | SJSBP2 | 1 | 12 | 0.145 | 0.136 | 0.060 | Moderate | Yes | Moderate | |
| Liberty | ANF3 | 1 | 18 | 0.126 | 0.132 | −0.02 | Very High | Yes | High | |
| Liberty | ANF4 | 3 | 34 | 0.138 | 0.116 | 0.154 | Low | Yes | Moderate | |
| Liberty | ANF5 | 2 | 11 | 0.124 | 0.044 | 0.504 | Moderate | No | Low | |
| Total: | 17 | 191 | Mean: | 0.134 | 0.106 | 0.176 |
| Population | N | Clonal Diversity Threshold | Number of Unique Clones | Number of Ramets per Unique Clone | CR |
|---|---|---|---|---|---|
| ANF1 | 19 | 0.01 | 16 | 4, 1 (15) | 0.83 |
| ANF2 | 14 | n/a | 14 | 1 (14) | 1.00 |
| ANF3 | 18 | 0.02 | 15 | 3, 2, 1 (13) | 0.82 |
| ANF4.1 | 7 | 0.01 | 5 | 3, 1 (4) | 0.67 |
| ANF4.2 | 15 | 0.01 | 11 | 5, 1 (10) | 0.71 |
| ANF4.3 | 11 | n/a | 11 | 1 (11) | 1.00 |
| ANF5 | 11 | 0.02 | 1 | 11(1) | 0 |
| BRWMA1 | 10 | 0.002 | 3 | 8, 1 (2) | 0.22 |
| BRWMA2 | 11 | 0.02 | 8 | 3, 2, 1 (6) | 0.70 |
| LB1 | 23 | 0.02 | 3 | 18, 3, 2 | 0.09 |
| SJSBP1 | 7 | 0.01 | 4 | 4, 1 (3) | 0.50 |
| SJSBP2 | 12 | 0.03 | 10 | 2, 2, 1 (8) | 0.82 |
| THSF1 | 14 | 0.03 | 2 | 13, 1 | 0.08 |
| THSF2 | 18 | 0.01 | 14 | 3, 2, 2, 1 (11) | 0.76 |
| Total, or Average ± standard error | 190 | 117 | 1.7 ± 0.23 | 0.59 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanko, G.R.; Vogel, M.T.; Negrón-Ortiz, V.; Moore, R.C. High Prevalence of Clonal Reproduction and Low Genetic Diversity in Scutellaria floridana, a Federally Threatened Florida-Endemic Mint. Plants 2023, 12, 919. https://doi.org/10.3390/plants12040919
Hanko GR, Vogel MT, Negrón-Ortiz V, Moore RC. High Prevalence of Clonal Reproduction and Low Genetic Diversity in Scutellaria floridana, a Federally Threatened Florida-Endemic Mint. Plants. 2023; 12(4):919. https://doi.org/10.3390/plants12040919
Chicago/Turabian StyleHanko, Gina Renee, Maria Therese Vogel, Vivian Negrón-Ortiz, and Richard C. Moore. 2023. "High Prevalence of Clonal Reproduction and Low Genetic Diversity in Scutellaria floridana, a Federally Threatened Florida-Endemic Mint" Plants 12, no. 4: 919. https://doi.org/10.3390/plants12040919