2.1.1. Paris yunnanensis Franch. Mem. Philom. Cent. (Paris) 24: 290. 1888. (Figure 1)
Paris yunnanensis Franch; C. H. Wright, Journ. Linn. Bot. 36:145. 1903.—Paris polyphylla Smith var. yunnanensis (Franch.) Hand.-Mazz., Symb. Sin. 7: 1216. 1936; Hara, Journ. Fac. Sci. Univ. Tokyo. Sect. 3, 10 (10): 154. 1069; Wang et Tang, Fl. Reip. Pop. Sin. 15: 95. 1978; H. Li, Bull. Bot. Res. Harbin 6 (1): 119. 1986; H. Li, The Genus Paris (Trilliaceae) 35. 1998; S. Y. Liang et V. G. Soukup, Fl. Chin. 24: 90. 2000.—Daiswa yunnanensis (Franch.) Takht., Brittonia 35 (3): 257. 1983; B. Mitchell, Plantsman 10 (3): 185. 1988. Type: China, Yunnan, Eryuan, 2000 m, J. M. Delavay 2227 (holotype P!).
Paris christii Lévl., Bull. Acad. Inter. Geogr. Bot. 12: 255. 1903. Type: China, Guizhou, Bodinier s. n. (holotype E!). (synonymized in [
9]).
Paris mercieri Lévl., Bull. Acad. Int. Géogr. Bot. 12: 255. 1903. Type: China, Guizhou, Guiyang, 17 July 1898, Bodinier 1635 (holotype E!). (synonymized in [
9]).
Paris franchetiana Lévl., Bull. Acad. Int. Géogr. Bot. 12: 255. 1903. Type: China, Guizhou, Bodinier 712 (holotype E!). (synonymized in [
9]).
Paris cavaleriei Lévl. et Vaniot, Nouv. Contrib. Liliac. Chine 24: 354. 1906. Type: China, Guizhou, Longli, 13 June 1902, Cavalerie 1310 (holotype E!). (synonymized in [
9]).
Paris gigas Lévl. et Vaniot, Nouv. Contrib. Liliac. Chine 24: 354. 1906. Type: China, Guizhou, Guiding, 23 November 1902, Cavalerie 729 (holotype E!). (synonymized in [
9]).
Paris pinfaensis Lévl., Repert. Spec. Nov. Regni Veg. 6: 265. 1909. Type: China, Guizhou, Guiding, June 1907, Cavalerie 2023 (holotype E!, isotype GH, K!). (synonymized in [
9]).
Paris aprica Lévl., Repert. Spec. Nov. Regni Veg. 6: 265. 1909. Type: China, Guizhou, Guiding, 25 June 1907, Cavalerie 3023 (holotype E!, isotype E!). (synonymized in [
9]).
Paris atrata Lévl., Repert. Spec. Nov. Regni Veg. 12: 536. 1913. Type: China, Yunnan, Ninglang, Maire s. n. (holotype E!). (synonymized in [
9]).
Paris polyphylla Smith var.
nana H. Li, Bull. Bot. Res. Harbin 6 (1): 123. 1986. Maity et al., J. Bot. Soc. Bengal 68: 117. 2014. Type: China, Sichuan, Yibin, 07 July 1977, Yibin Drug Inspection Institute Yi428 (KUN). (synonymized in [
2]).
Paris daliensis H. Li et V. G. Soukup, Act. Bot. Yunnanica, Supp. v: 15–16, Figure 3. 1992; H. Li, The Genus Paris (Trilliaceae) 32. 1998; S. Y. Liang et V. G. Soukup, Fl. Chin. 24: 89–90. 2000. Type: China, Yunnan, Dali, 2600 m, 06 Nov. 1986, H. Li & V. G. Soukup 1098 (holotype KUN, isotype CINC). (synonymized in [
2]).
Paris birmanica (Takht.) H. Li et H. Noltie., Edinb. J. Bot. 54 (3): 351–352. 1997; H. Li, The Genus Paris (Trilliaceae). 28. 1998.
Daiswa birmanica Takht., Brittonia 35 (3): 259, Figure 2. 1983; B. Mitchell, Plantsman 10 (3): 169. 1988; Type: Myanmar, Maymyo, 22 June 1913, Lace John Henry 6233 (holotype E!, isotype E!). (synonymized in [
2]).
Paris polyphylla Smith var.
emeiensis X. H. Yin, H. Zhang & D. Xue, Acta Phytotaxon. Sin. 45 (6): 822–827. 2007. Type: China, Sichuan, Mt. Emei, 1900 m, 27 April 2006, Yin XH et al. 06121 (holotype SZ, isotype, CDBI). (synonymized in [
2]).
Perennial herb, (20–)27.7–121.5(–130) cm tall. Rhizome thick, cylindrical, conical, to C-shaped, 4–19 cm long, 1.5–7(–10) cm in diam., glabrous, with 2–12 scars on the flowering state. Aerial stems 15–105 cm long, 0.5–1.8 cm in diam., purple to green, or green at the upper part, glabrous to remotely short spiny. Leaves 5–11; petiole 1–6.5(–8) cm long, 1.5–4 mm in diam., purple above, glabrous; lamina 7.7–25 × 4.3–10.5(–15) cm, obovate, obovate, to oblanceolate, base cuneate, apex acute, width/length ratio 0.29–0.71: lateral veins two pairs. Flower solitary, terminal; pedicel (2.5–)5.3–24(–30) cm long, 1.5–6 mm in diam., green to purple with green at the upper part, glabrous; Sepals (3–)4–8(–10), leaf-like, 4.2–14.5 × 1.3–5 cm, lanceolate to elliptic, width/length ratio 0.17–0.44, base cuneate to obtuse, apex acute, green, glabrous; petals (3–)5–8(–10), (1.2–)3–7.5(–8.5) cm × 0.5–2(–2.5) mm, linear, protruding above the sepal, yellow to green: upper part (0.8–)1.5–2(–2.5) cm × (0.5–)1–2(–3) mm, yellow, petals/sepal length ratio (0.19–)0.33–0.89; Stamen (8–)10–22(–25), stamens/petals number ratio (1.83–)2–3.2(–4.4): filaments (1–)2–6(–7) mm long, (0.3–)1(–2) mm in diam.: anthers 7–13(–17) mm long; free portion of connective up to (0.2–)0.5–1.5 × (0.2–)1–1.5 mm, green to yellow to brown, glabrous, anther/filament length ratio (1.43–)2–6. Pistil 1: style with an enlarged base: Stylar base purplish red to orange: Ovary 4–8(–10) angled, single-locular with parietal placentation: stigma (3–)4–7(–15), (2–)3–8(–12) mm long, strongly curved, orange to red. Fruits dehiscent capsule, angular, 4–8(–10) angled; immature fruit up to 27 mm long, up to 40 mm in diam., green to dark green, yellow at ovary base, glabrous. Seed white covered by red to orange: sarcotesta red to orange.
Phenology. Flowering April–August. Fruiting July–February.
Distribution. Myanmar (Mandalay and Shan). China (Chongqing, Guangxi, Guizhou, Sichuan, Tibet, Yunnan). Thailand (Northern).
Additional Specimens Examined. THAILAND. NORTHERN. Chiang Mai.
[Chiang Dao Distr., Doi Chaing Dao Wildlife Sanctuary, 1400 m alt., 6 February 2012, V. Chamchumroon et al. V.C. 1764 (BKF141913); ibid., 1200 m alt., 6 December 1959, T. Smitinand & E.C. abbe 6213 (BKF24723); ibid., 1020 m alt., 23 June 1998, Khantchai 192 (BKF24021); ibid., 7 May 1999, PS. Plernchit 1205 (BKF15756); ibid., 1400–1500 m alt., 16 December 1983, N. Fukuoka & M. Ito T-35234 (BKF109875, L3915025); ibid., 22 December 1931, K. Larsen et al. 2919 (L.1460262); ibid., 22/12/1931, Put s. n. (BK25671). Chom Thong Distr., Bann Angka Noi, Doi Inthanon National Park, 1160 m alt., 4 August 2003, TH. Wongprasert & S. Khaoiam 038–63 (BKF157006); ibid., 900 m alt., 26 June 1978, C. Phengkai et al., 4118 (BKF); Doi Inthanon National Park, Mae Pan waterfall, 1100 m alt., 16 October 1979, T. Shimizu et al. 18975 (BKF76677); Doi Inthanon, 1020 m alt., 22 July 1988, M.N. Tamura T-60181 (BKF94753); ibid., 1150 m alt, 22 October 1999, Th. Wongprasert s. n. (BKF128662). Pha Mon, 2 January 1981, Somkid 115 (BKF110099), Doi Inthanon National Park, Mae Klang Luang Village, 1615 m alt, 23 May 2011, Pavlos Georgiadis 615 (L4345332), Mae Soi Ridge, Mae Soi Subdistr., Ban Bah Gluay (Meo village), 16 June 1991, JF. Maxwell 91-551 (L3896551); ibid., 8 May 1992, J.F. Maxwell 92-189 (L3893265); Doi Saket Distr., Thep Sadet, 1206 m alt, 16 July 2020, S. Ruchisansakun et al. PJDSK63008 (QBG); S. Ruchisansakun et al. PJDSK63010 (QBG); S. Ruchisansakun et al. PJDSK63013 (QBG); S. Ruchisansakun et al. PJDSK63014 (QBG); S. Ruchisansakun et al. PJDSK63015 (QBG); S. Ruchisansakun et al. PJDSK63018 (QBG); S. Ruchisansakun et al. PJDSK63020 (QBG); S. Ruchisansakun et al. PJDSK63021 (QBG); S. Ruchisansakun et al. PJDSK63025 (QBG); S. Ruchisansakun et al. PJDSK63026 (QBG); S. Ruchisansakun et al. PJDSK63027 (QBG); S. Ruchisansakun et al. PJDSK63028 (QBG); S. Ruchisansakun et al. PJDSK63030 (QBG); S. Ruchisansakun et al. PJDSK63032 (QBG); S. Ruchisansakun et al. PJDSK63035 (QBG); S. Ruchisansakun et al. PJDSK63037 (QBG); S. Ruchisansakun et al. PJDSK63040 (QBG); S. Ruchisansakun et al. PJDSK63044 (QBG); Chai Prakan Distr., Wiang Pha, 800 m alt., 11 November 2021, S. Ruchisansakun et al. PW641111 (TR64) (QBG); Fang Distr., Doi Fa Hom Pok NP, 19 December 2010, P. Phonsena et al. 6673 (L3809213); Hod Distr., Mai Muang Nao Arboretum, Ban Mae Sanam Mai, Baw Salee Subdistr., 15 May 2001, W. Sankamethawee 175 (L3809523); Mae Chaem Distr., Mae Suek, Ban Pha La Pi, 1541 m alt., 18 June 2022, S. Ruchisansakun et al. PW650618 (QBG); S. Ruchisansakun et al. PW650619 (QBG); Mae Taeng Distr., Pa Pae, 14 June 2020, S. Ruchisansakun et al. PJMT63008 (QBG); PJMT63010 (QBG); PJMT63012 (QBG); PJMT63013 (QBG); PJMT63017 (QBG); PJMT63018 (QBG); Mae Rim Distr., Pong Yaeng, Queen Sirikit Botanical Garden, 15 July 2020, S. Ruchisansakun et al. PJHL63003 (QBG); PJHL63004 (QBG); PJHL63005 (QBG); PJHL63006 (QBG); PJHL63007 (QBG), PJHL63008 (QBG); PJHL63020 (QBG); PJHL63021 (QBG); PJHL63022 (QBG); Queen Sirikit Botanical Garden, 28 September 2005, Tillich 5045 (BKF163774); Mae Rim Distr., Mae Sa valley, 1000 m alt. 17 May 1974, J.K. Jackson 6052 (BKF58733); Mae Rim Distr., Saluang, Phraphutthabat Si Roi, 5 August 2020, S. Ruchisansakun et al. PJPPB63012 (QBG); PJPPB63013 (QBG); PJPPB63014 (QBG); Mae Wang Distr., Mae Win, Ban Pah Kluai, 1110 m alt, 20 June 2022, S. Ruchisansakun et al. PW650620 (QBG); Mueang Chiang Mai Distr., Mueang Chiang Mai Distr., Doi Pui, ca. 1500 m alt., 14 October 1979, T. Shimizu et al. 18637 (BKF76676); Doi Suthep, 25 July 1991, Soradet SS. 159 (BKF4372); ibid., 24 April 1977, Sin Boonchu 1860 (BKF112159); ibid., 5 December 1987, J.F. Maxwell 87-1542 (L1460259, L1460260); ibid., 3 May 1958, T. Sorensen 3229 (L1460261); Suthep-Pui National Park, 1300–1500 m alt., 22 October 1988, C. Phengklai et al. 6541 (BKF91009); Payap, Doi Buak Ha, 1150 m alt., 29 November 1965, E. Hennipman 3165 (BKF39737, L1449986, L1449987); Samoeng Distr., Maerim-Samueng road, Maeo village, 900 m alt., 28 December 1987, R. Pooma 53 (BKF99839)]. Chiang Rai [Mae Fa Luang Distr., Mae Salong Nok, 1207 m alt., 22 April 2022, P. Umpunjun et al. PW650422 (QBG), Mueang Chiang Rai Distr., Huai Chomphu, Saroj Ruchisansakun, et al. PW641021 (QBG); Mae Sai Distr., Wat Phra Thard Doi thung, 1315 m, 22 May 2008, V. Chamchumroon et al. 1593 (BKF158522); Mae Suai Distr., Mae Suai, K. Bunchuai & B. Nimanong 1473 (BKF36501); Doi Wa Wee, 1300 m alt., 1994, Y. Prasorksantisatang s. n. (BK62091); Ban San Sa At., 30 July 1967, Prayad 952 (BK40805); Phan Distr., Wiang Bah Pao, Doi Luang National Park, 1300–1400 m alt., 28 October 1997, J.F. Maxwell 97-1263 (BKF121813, L3896511); ibid., 1200 m alt, 24 May 1998, J.F. Maxwell 98-557 (BKF121738, L3896507)]. Lampang [Hang Chat Distr., Khun Tarn, 10 February 1976, Put 125 (BKF16254); Mae Tha Distr., Doi Khun Tan, 1000–1370 m, 27 December 1984, H. Koyama & C. Phengklai T-39059 (BKF110946); ibid., H. Koyama & C. Phengklai T-39059 (BKF110946); Mueang Pan Distr., Mueang Pan, Khun Chae National Park, 1427 m alt., 6 August 2020, S. Ruchisansakun et al. PJCS63002 (QBG); PJCS63004 (QBG); Chae Son, 1210 m alt, 9 November 2021, S. Ruchisansakun et al. PW641109 (QBG)]. Lamphun [Mae Tha Distr., Tha Pla Duk, 17 June 2022, S. Ruchisansakun MW650607 (QBG); Khun Tan, 1200 m alt., 2 November 1975, T. Smitinand 12080 (BKF63463); ibid., 1050 m alt., 5 June 1993, J.F. Maxwell 93-578 (BKF152332, L3808810)]. Mae Hong Son [Mae Hong Son, Khun Yuam Distr., Mae U Kho, 1240 m alt., 10 November 2021, S. Ruchisansakun et al. PW641110 (QBG); ibid., 1360 m alt., 19 June 2022, S. Ruchisansakun et al. PW641110 (QBG); 26 June 1998, Chusie KY392 (QBG47309)]. Phayao [1510 m alt, 6 May 2015, M. Norsaengsri 12354 (QBG82970); 1054 m alt., 31 June 2016, N. Muangyen 1053 (QBG91043)].
Vernacular name. ตีนฮุ้งดอย สัตตฤาษี จ้วงนางแฮะ (Chiang Mai) ว่านตีนฮุ้ง (Lampang).
Karyotype. 2n = 2x = 10 = 6m + 4t, 6m + 4st, or 4m + 2sm + 4st [
10].
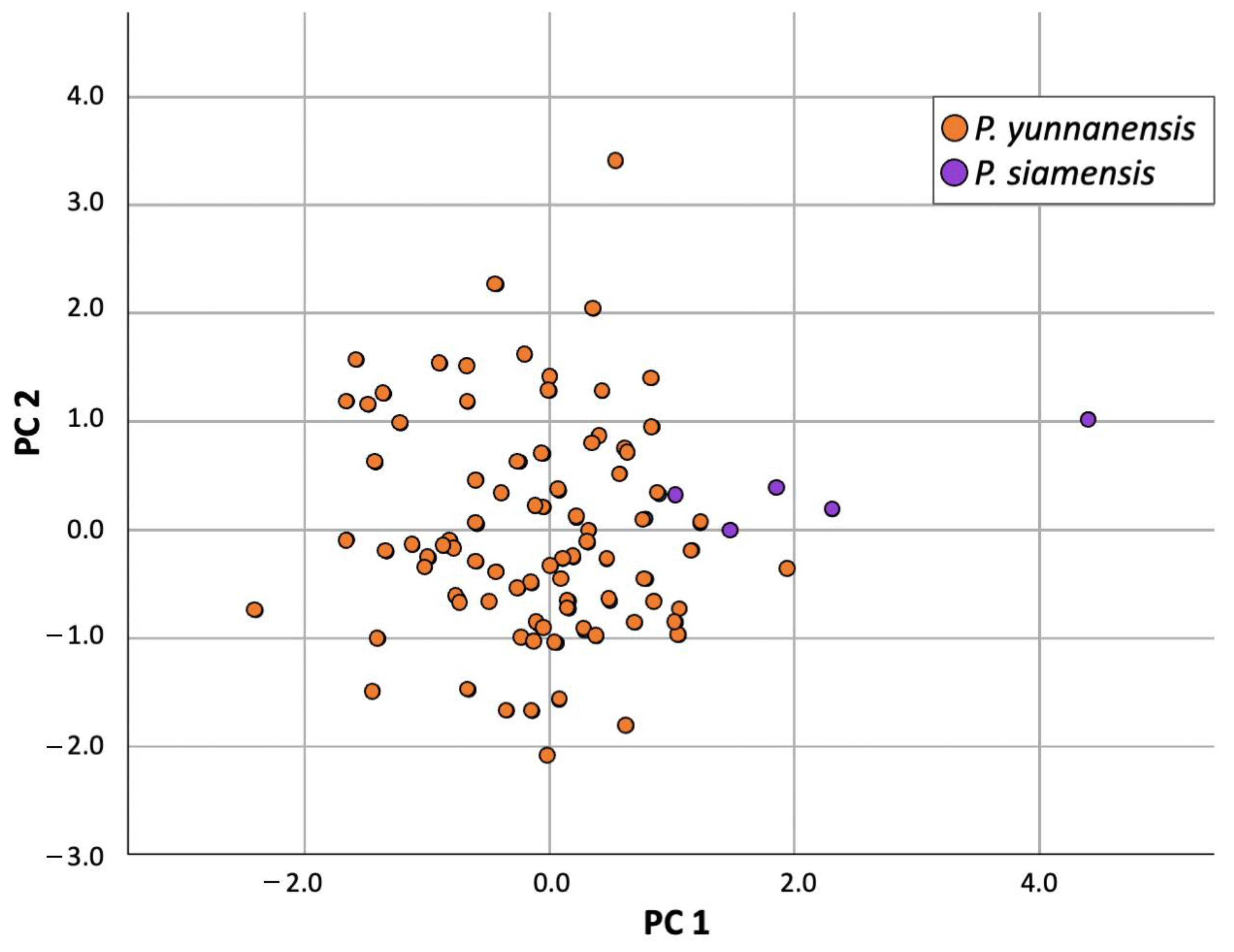
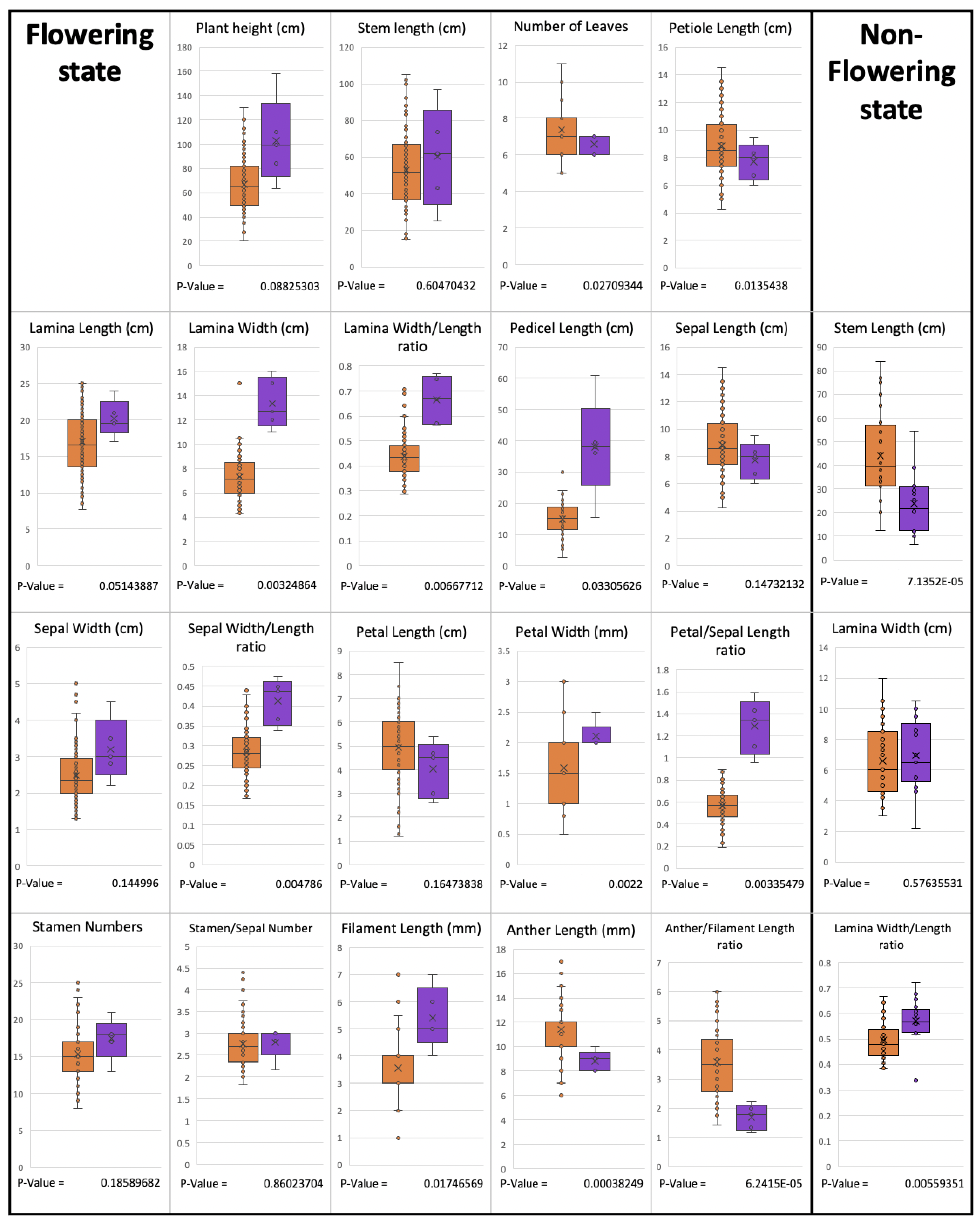
Note. Paris yunnanensis is a cryptic species with very high variation in its morphology. The population in Thailand differs from
P. yunnanensis in China [
8] in many characters (as shown in
Table 1), such as having petals above sepals (versus petals below sepals) and petals that are shorter than sepals (versus longer than or as long as sepals). The plant from the locality of
J.F. Maxwell 98-557, which was identified as
P. cabangensis [
2], was also proven to be
P. yunnanensis.
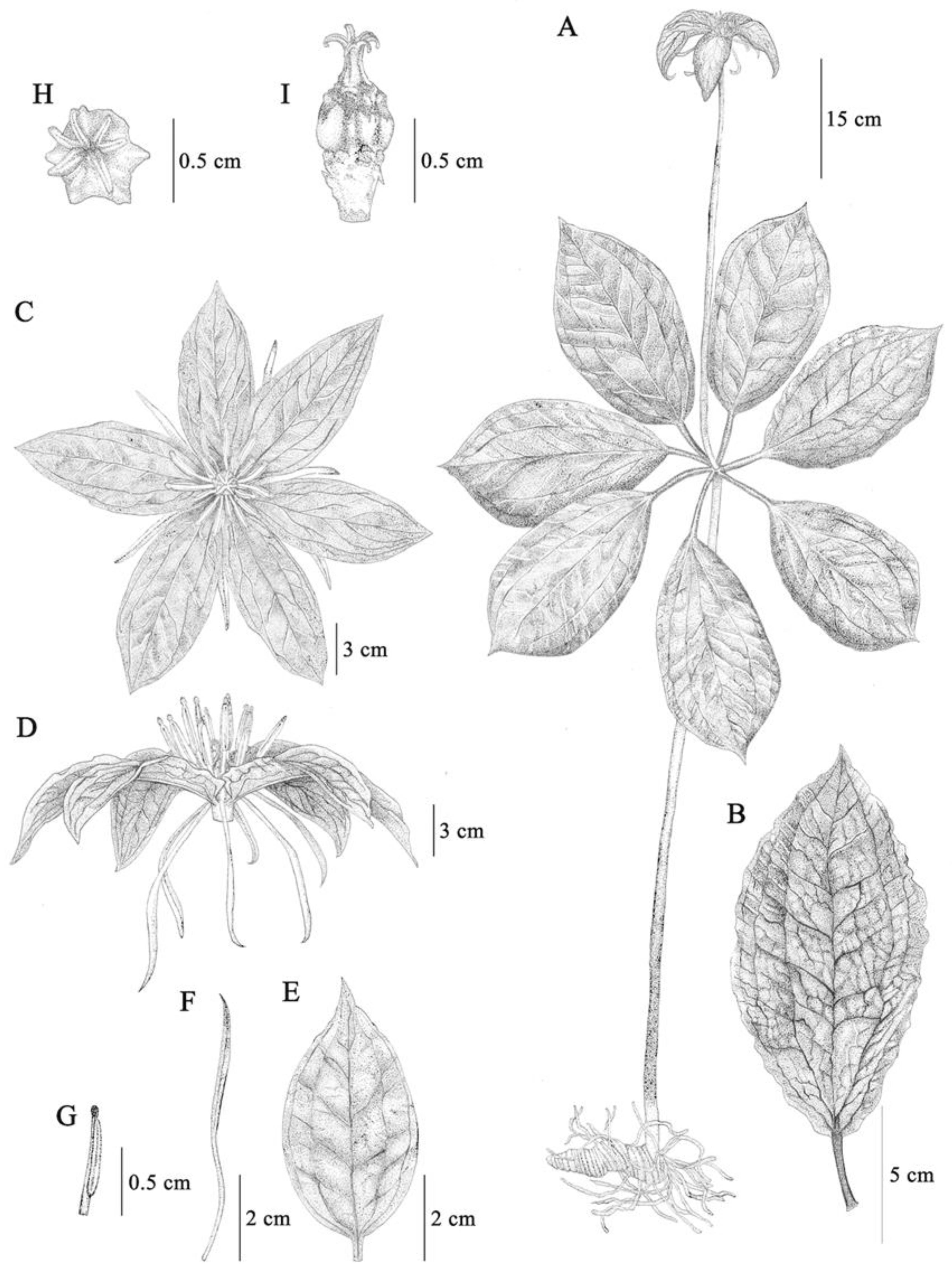
Figure 1.
Paris yunnanensis Franch. (A) Habit, (B) Leaf, (C) Flower (front view), (D) Flower (lateral view), (E) Sepal, (F) Petal, (G) Stamen, (H) Pistil (front view), (I) Pistil (lateral view), (J) Fruit.
Figure 1.
Paris yunnanensis Franch. (A) Habit, (B) Leaf, (C) Flower (front view), (D) Flower (lateral view), (E) Sepal, (F) Petal, (G) Stamen, (H) Pistil (front view), (I) Pistil (lateral view), (J) Fruit.
2.1.2. Paris siamensis Ruchis. (Figure 2 and Figure 3)
Paris siamensis is similar to P. liiana but differs in having petals below sepals (versus petals above sepals), stamen number (2.2–)2.8–3 times petal number (versus stamen number 2 times petal number), and anther length 8–10 mm long (versus anther 15–40 mm long). Type: Thailand, Northern, Nan, Mae Charim Distr., Nong Daeng, 18°48′04.3″ N 101°12′47.0″ E, 1216 m alt, 21 April 2022, P. Umpunjun et al. PW650421 (holotype QBG!, isotypes QBG!).
Perennial herb, 63.5–110(–158) cm tall. Rhizome thick, cylindrical, conical, to C-shaped, 9.5–15(–28.5) cm long, 2.5–4 cm in diam., glabrous, with 5–7 scars on the flowering state. Aerial stems (25–)–43–97 cm long, (0.85–)1–1.5 cm in diam., purple to green, or green at the upper part, glabrous to remotely short spiny. Leaves 6–7; petiole (4.5–)6–9 cm long, 3–4 mm in diam., purple above, glabrous; lamina 17–24 × 11–16 cm, ovate, elliptic, to obovate, base cuneate, to round, apex acute, width/length ratio 0.56–0.77: lateral veins two pairs. Flower solitary, terminal; pedicel 15.5–40(–61) cm long, 3–5 mm in diam., green to purple with green at the upper part, glabrous; Sepals 6–7, leaf-like, (6–)6.7–9.5 × 2.2–4.5 cm, ovate to narrowly ovate, width/length ratio 0.34–4.7, base obtuse to stipitate, apex acute, green, glabrous; petals 6–7, 2.6–5.4 cm × 1–3 mm, linear, protruding under the sepal, yellow to green: upper part 2.3–2.5 cm × 2–2.5 mm, petals/sepal length ratio (0.27–)0.65–0.75; Stamen (13–)17–21, stamens/petals number ratio (2.2–)2.8–3: filaments 4–7 mm long, ca. 1 mm in diam.: anthers 8–10 mm long: free portion of connective 1–1.5 × ca. 1 mm, yellow to brown, glabrous, anther/filament length ratio (1.14–)1.8–2.25. Pistil 1: style with an enlarged base: Stylar base purple to orange: Ovary 6–7 angled, single-locular with parietal placentation: stigma 6–7, ca 4 mm long, strongly curved, orange to red. Fruits dehiscent capsule, angular, 6–7 angled; immature fruit up to 9 mm long, up to 13 mm in diam., green to dark green, yellow at ovary base, glabrous. Seed white covered by red to orange: sarcotesta red to orange.
Phenology. Flowering April–August. Fruiting July–February.
Distribution. To date, endemic to Nan province, Thailand (may be found in Laos), 1050–1216 m alt.
Vernacular name. Tin Hung Doi Siam (ตีนฮุ้งดอยสยาม), Tin Hung Doi (ตีนฮุ้งดอย).
Additional Specimens Examined. THAILAND. NORTHERN. Nan.
[Bo Kluea Distr., Dong Phaya, 20 April 2022, P. Umpunjun et al. PW650420 (QBG);
Mae Charim Distr., Nong Daeng, 1216 m alt.; ibid., 750 m alt. 26 December 2016, V. Nguanchoo 1016 (QBG102719); Pua Distr., Phu Kha, Doi Phu Kha National Park, 1050 m alt, 19 October 2021, S. Ruchisansakun et al. PW641019 (QBG); ibid., 1100 m alt., R. Pooma 1098 (BKF102154); ibid., 1600 m alt., 10 April 1991, R. Pooma 467 (BKF97579); ibid., 1200 m alt., 2 July 1999, P. Srisanga et al. 837 (QBG14625); ibid., 1700 m alt., 11 December 2002, P. Srisanga 2657 (QBG23648); ibid., 1280 m alt., 19 June 2010, K. Srithi 491 (QBG65084)].
Karyotype. 2n = 2x = 10 = 6m + 4t [
10].
Note.
Paris siamensis is similar to
P.
liiana,
P.
yunnanensis (population in China), and
P.
chinensis. It differs from
P.
liana, as mentioned in the species diagnosis. Differences compared to other species are shown in
Table 1.
Figure 2.
Paris siamensis Ruchis. (A) Habit, (B) Leaf, (C) Flower (front view), (D) Flower (lateral view), (E) Sepal, (F) Petal, (G) Stamen, (H) Pistil (front view), (I) Pistil (lateral view).
Figure 2.
Paris siamensis Ruchis. (A) Habit, (B) Leaf, (C) Flower (front view), (D) Flower (lateral view), (E) Sepal, (F) Petal, (G) Stamen, (H) Pistil (front view), (I) Pistil (lateral view).
Figure 3.
Paris siamensis Ruchis. (A) Plant, (B) Leaf, (C) Flower (front view), (D) Flower (lateral view), (E) Sepal, (F) Petal, (G) Stamen, (H) Pistil (front view), (I) Pistil (lateral view). (Drawn by Jeerapach Monthanom).
Figure 3.
Paris siamensis Ruchis. (A) Plant, (B) Leaf, (C) Flower (front view), (D) Flower (lateral view), (E) Sepal, (F) Petal, (G) Stamen, (H) Pistil (front view), (I) Pistil (lateral view). (Drawn by Jeerapach Monthanom).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}