
New Records of Wood- and Bark-Inhabiting Nematodes from Woody Plants with a Description of Bursaphelenchus zvyagintsevi sp. n. (Aphelenchoididae: Parasitaphelenchinae) from Russia

Abstract
:1. Introduction
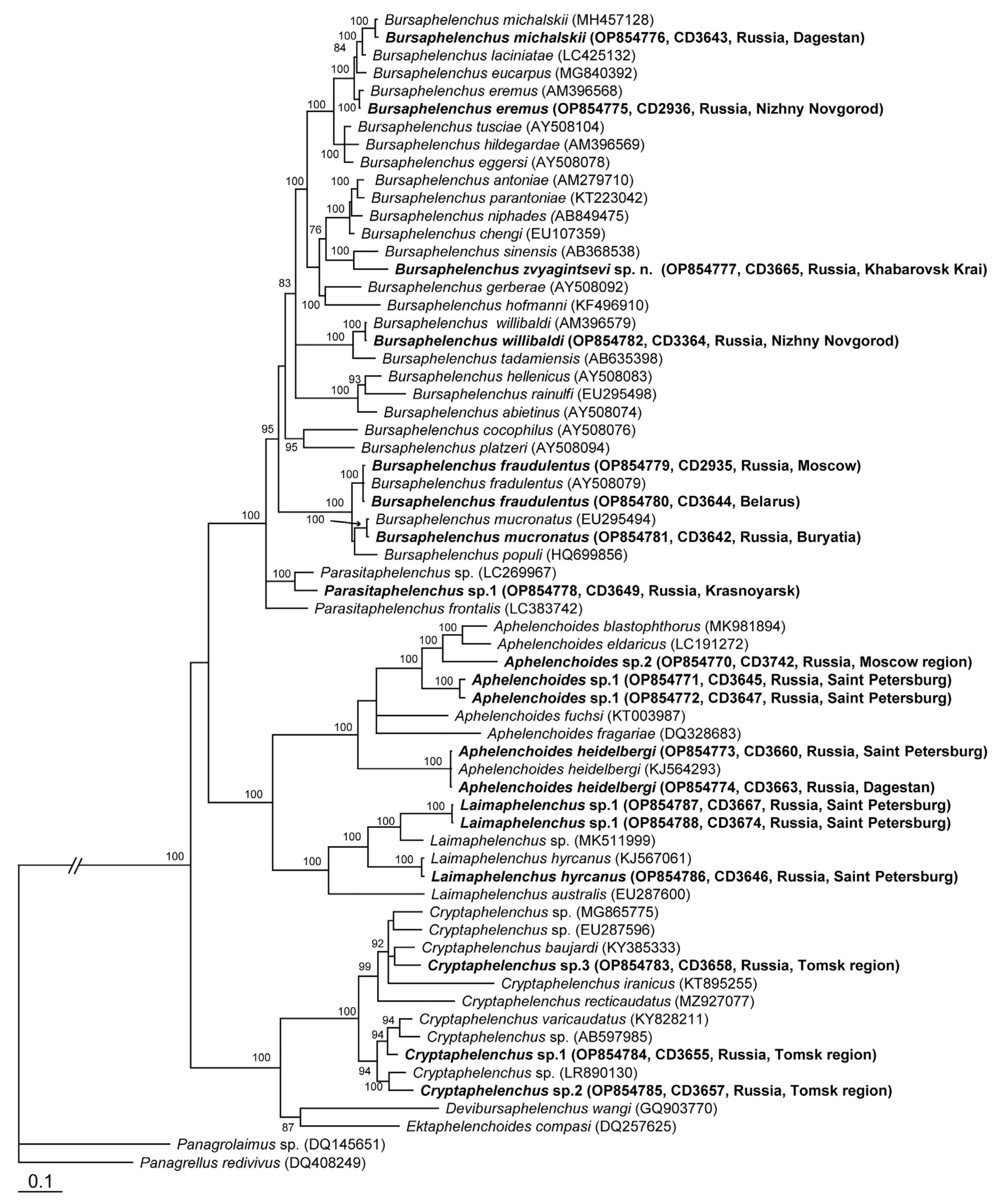
2. Results and Discussion
2.1. Species Identification and Ecological Grouping
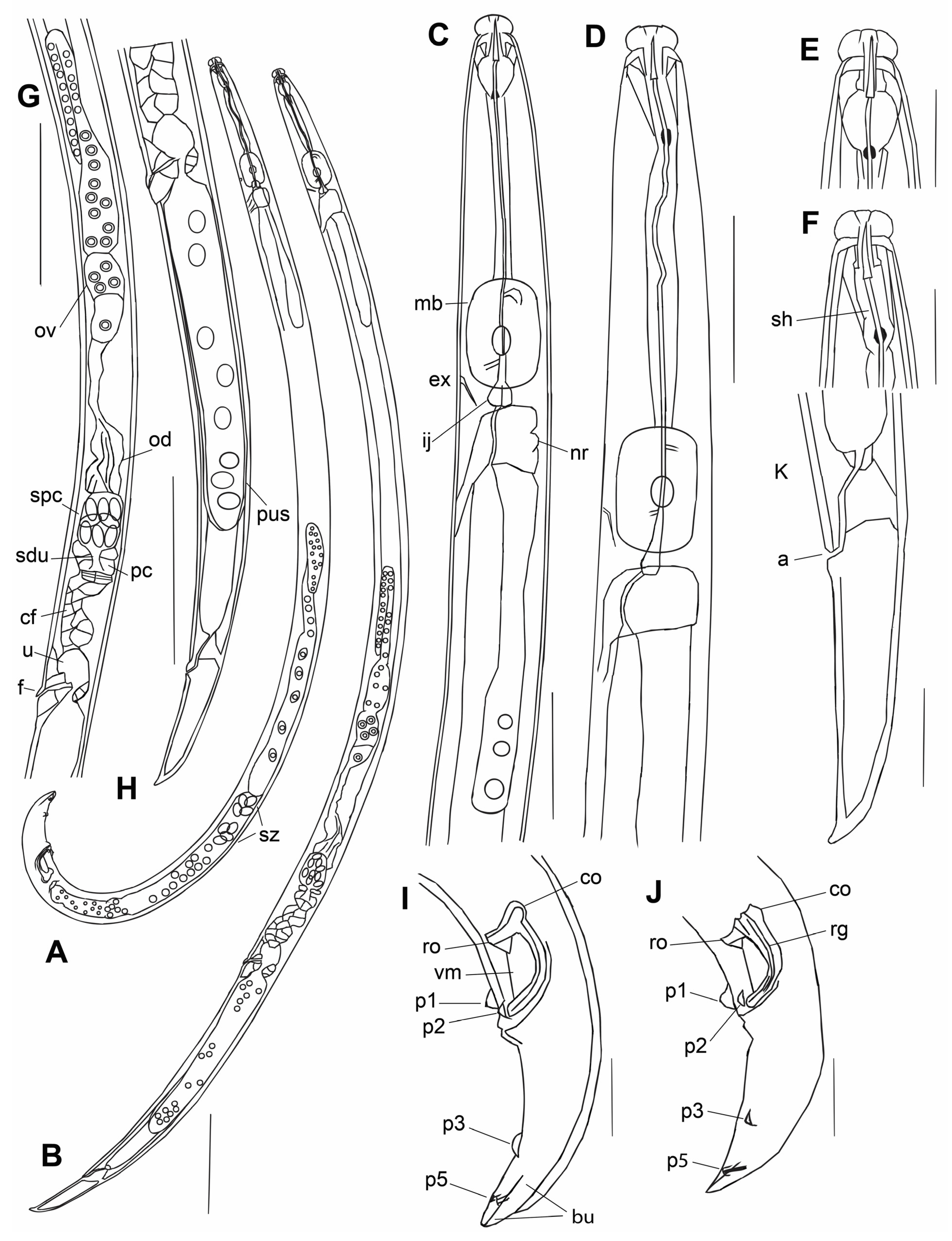
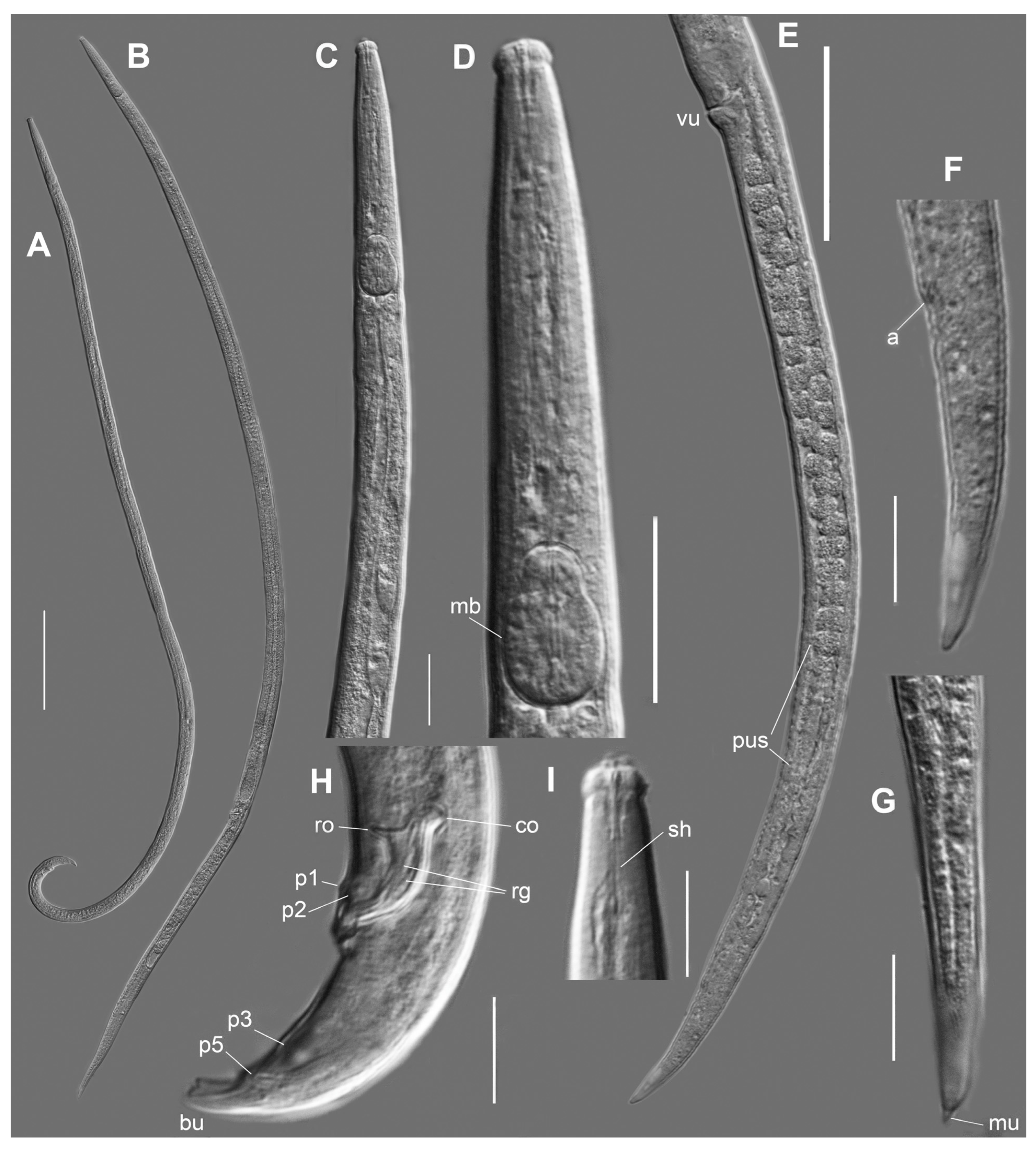
Bursaphelenchus zvyagintsevi sp. n.
2.2. Bursaphelenchus michalskii Tomalak and Filipiak, 2018
2.3. Characterization of Other Nematodes from Woody Plants
2.3.1. Aphelenchoides
2.3.2. Bursaphelenchus
2.3.3. Cryptaphelenchus
2.3.4. Deladenus
2.3.5. Diplogasteroides
2.3.6. Laimaphelenchus
2.3.7. Micoletzkya
2.3.8. Parasitaphelenchus
2.3.9. Parasitorhabditis
2.3.10. Tylenchid nematodes
2.4. General Discussion
3. Conclusions
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. Morphological Study
4.3. Molecular and Phylogenetic Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leoni, C.; Piancone, E.; Sasanelli, N.; Bruno, G.; Manzari, C.; Pesole, G.; Ceci, L.; Volpicella, M. Plant health and rhizosphere microbiome: Effects of the bionematicide Aphanocladium album in tomato plants infested by Meloidogyne javanica. Microorganisms 2020, 8, 1922. [Google Scholar] [CrossRef]
- Hao, X.; Wang, B.; Chen, J.; Wang, B.; Xu, J.; Jialiang, P.; Ma, L. Molecular characterization and functional analysis of multidrug resistance-associated genes of Pinewood nematode (Bursaphelenchus xylophilus) for nematicides. Pestic. Biochem. Physiol. 2021, 177, 104902. [Google Scholar] [CrossRef] [PubMed]
- Hough, J.; Howard, J.; Brown, S.; Portwood, D.; Kilby, P.; Dickman, M. Strategies for the production of dsRNA biocontrols as alternatives to chemical pesticides. Front. Bioeng. Biotechnol. 2022, 10, 980592. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.; Inacio, M.L.; Mota, M. The potential of Esteya spp. for the biocontrol of the pinewood nematode, Bursaphelenchus xylophilus. Microorganisms 2022, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Ryss, A.Y.; Polyanina, K.S.; Popovichev, B.G.; Subbotin, S.A. Description of Bursaphelenchus ulmophilus sp. n. (Nematoda: Parasitaphelenchinae) associated with Dutch elm disease of Ulmus glabra Huds. in the Russian North West. Nematology 2015, 17, 685–703. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Polyanina, K.S.; Popovichev, B.G.; Krivets, S.A.; Kerchev, I.A. Plant host range specificity of Bursaphelenchus mucronatus Mamiya et Enda, 1979 tested in the laboratory experiments. Parazitologiya 2018, 52, 32–40. [Google Scholar]
- Ryss, A.Y.; Polyanina, K.S.; Petrov, A.V.; Sazonov, A.A.; Mandelshtam, M.Y.; Subbotin, S.A. Report of Bursaphelenchus crenati Rühm, 1956 (Nematoda: Parasitaphelenchinae) from Belarus and Russia with a diagnostic key and phylogeny of the Sexdentati group. For. Pathol. 2019, 49, e12534. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Subbotin, S.A. Coevolution of wood-inhabiting nematodes of the genus Bursaphelenchus Fuchs, 1937 with their insect vectors and plant hosts. Zhurnal Obs. Biol. 2017, 78, 32–61. [Google Scholar]
- Lazarevskaya, S.L. On the nematode fauna of Acanthocinus aedilis (Cerambycidae). Helminthologia 1961, 3, 212–220. [Google Scholar]
- Lazarevskaya, S.L. Nematode fauna of Orthotomicus laricis and O. proximus (Coleoptera: Ipidae). Helminthologia 1963, 4, 254–265. [Google Scholar]
- Slankis, A. New nematode species Aphelenchoides macromucrons (Tylenchida) from bark beetle Ips typographus. Proc. Helminthol. Lab. AN SSSR 1967, 18, 279–282. [Google Scholar]
- Korentchenko, E.A. New species of nematodes from the family Aphelenchoididae, parasites of stem pests of the dahurian larch. Zool. Zhurnal 1980, 59, 1768–1780. [Google Scholar]
- Korentchenko, E.A. Cryptaphelenchus diversispicularis n. sp. (Tylenchida, Aphelenchoididae)—A new nematode of the bark beetle, Ips subelongatus (Coleoptera: Ipidae). Parazitologiya 1987, 21, 73–78. [Google Scholar]
- Kulinich, O.A. Aphelenchoides parasexlineatus sp. n. from seedlings of coniferous trees. Zool. Zhurnal 1984, 63, 1570–1572. [Google Scholar]
- Kulinich, O.A. Parasitic Nematodes of Conifers in Russia. Ph.D. Thesis, Institute of Parasitology of the Russian Academy of Scences, Moscow, Russia, 2005; pp. 1–24. [Google Scholar]
- Kulinich, O.A.; Arbuzova, E.N.; Chalkin, A.A.; Kozyreva, N.I.; Ryss, A.Y. Distribution of the pinewood nematode, Bursaphelenchus xylophilus, in the world and results of conifer forest surveys in the Russian Federation. In Materials of the II International Research-to-Practice Conference in Commemoration of 95th Anniversary of Professor Nikolai Ilyich Fedorov, and the 90th Anniversary of the Department of Forest Protection and Wood Science, Minsk, Russia, 30 November–4 December 2020; Zviagintsev, V.B., Siaredzich, M.O., Eds.; BSTU: Minsk, Belarus, 2020; pp. 263–267. [Google Scholar]
- Eroshenko, A.S.; Volkova, T.V. Plant Parasitic Nematodes of the South of the Russian Far East; DVO AN SSSR: Vladivostok, Russia, 1988; 140p. [Google Scholar]
- Gagarin, V.G. Nematode fauna in rotten wood of birch, aspen, and pine in Borok (Yaroslavl’ district, Central Russia). Zool. Zhurnal 1999, 78, 146–157. [Google Scholar]
- Eroshenko, A.S.; Kruglik, I.A. Aphelenchoides paramonovi sp. n. (Nematoda: Aphelenchoididae)—A new species of nematodes from the pine wood in Primorsky Krai. In Parazitocheskie Nematody Rastenii i Nasekomykh; Sonin, M.D., Ed.; Nauka: Moscow, Russia, 2004; Volume 1, pp. 46–49. [Google Scholar]
- Khusainov, R.V.; Rogozhin, E.A. Contribution to the study of the xylobiont nematodes of the order Tylenchida and Aphelenchida in Russia. In Nematodes of the Natural and Transformed Ecosystems; Karelian Scientific Center of the Russian Academy of Sciences: Petrozavodsk, Russia, 2011; pp. 101–103. [Google Scholar]
- Kerchev, I.A.; Krivets, S.A.; Ryss, A.Y. Associations of parasitic nematodes of Polygraphus proximus Blandf. (Coleoptera: Curculionidae: Scolytinae) in West Siberia. In Abstracts of the 12th International Symposium of the Russian Society of Nematologists “Nematodes and other Ecdysozoa under the Growing Ecological Footprint on Ecosystems”, Nizhny Novgorod, Russia, 31 July–6 August 2017; Zryanin, V.A., Ryss, A.Y., Eds.; Lobachevsky State University of Nizhni Novgorod: Nizhny Novgorod, Russia, 2017; pp. 40–41. [Google Scholar]
- Kerchev, I.; Kryukova, N.; Ryss, A.; Glupov, V. Complex of endoparasitic nematodes of four-eyed fir bark beetle Polygraphus proximus, and their impact on its immunity. In Proceedings of the 16th meeting of the IOBC-WPRS Working Group, Tbilisi, Georgia, 11–15 June 2017; p. 83. [Google Scholar]
- Ryss, A.Y.; Mokrousov, M.V. The analysis of climatic and biological parameters for the pest spread risk modelling of the wood nematode species Bursaphelenchus spp. and Devibursaphelenchus teratospicularis (Rhabditida: Aphelenchoidea). Parazitologiya 2014, 48, 188–294. [Google Scholar]
- Ryss, A.Y.; Polyanina, K.S. Nematode impact in the Dutch elm disease in NW Russia: Entomophilic dauers in Scolytus spp. and their adult stages in wood. In Proceedings of the Eleventh International Symposium of the Russian Society of Nematologists, Cheboksary, Russia, 6–11 July 2015. [Google Scholar]
- Ryss, A.Y.; Polyanina, K.S. Xylobiont nematodes parasitizing elm Ulmus glabra in parks of St. Petersburg, Russia. In Proceedings of the Eleventh International Symposium of the Russian Society of Nematologists, Cheboksary, Russia, 6–11 July 2015. [Google Scholar]
- Ryss, A.Y.; Polyanina, K.S. Life cycle and population dynamics of the Rhabditolaimus ulmi (Nematoda: Rhabditida: Diplogastridae) in vitro. Russ. J. Nematol. 2022, 30, 21–30. [Google Scholar]
- Ryss, A.Y.; Polyanina, K.S. Life cycle and population dynamics of Bursaphelenchus willibaldi (Nematoda: Rhabditida: Aphelenchoididae) in vitro. Nematology 2022, 24, 1–15. [Google Scholar] [CrossRef]
- Polyanina, K.S.; Ryss, A.Y. Parameters of ontogeny and population dynamics modeling of Panagrolaimus detritophagus (Nematoda: Rhabditida) in vitro. Proc. Zool. Inst. RAS 2021, 325, 91–98. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Polyanina, K.S.; Álvarez-Ortega, S.; Subbotin, S.A. Morphology, development stages and phylogeny of the Rhabditolaimus ulmi (Nematoda: Diplogastridae), a phoront of the bark beetle Scolytus multistriatus from the elm Ulmus glabra Huds. in Northwest Russia. J. Nematol. 2021, 53, e2021–e2025. [Google Scholar] [CrossRef]
- Polyanina, K.S.; Mandelshtam, M.Y.; Ryss, A.Y. Brief review of the associations of xylobiont nematodes with bark beetles (Coleoptera, Curculionidae: Scolytinae). Entomol. Rev. 2019, 99, 598–614. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Parker, C.; Álvarez-Ortega, S.; Nadler, S.A.; Subbotin, S.A. Bursaphelenchus juglandis n. sp. (Nematoda: Aphelenchoididae), an associate of walnut twig beetle, Pityophthorus juglandis, the vector of thousand cankers disease. Nematology 2021, 23, 171–200. [Google Scholar] [CrossRef]
- Palmisano, A.M.; Ambrogioni, L.; Tomiczek, C.H.; Brandstetter, M. Bursaphelenchus sinensis sp. n. and B. thailandae Braasch et Braasch-Bidasak in packaging wood from China. Nematol. Mediterr. 2004, 32, 57–65. [Google Scholar]
- Giblin-Davis, R.M.; Kanzaki, N.; Ye, W.; Center, B.J.; Thomas, K.W. Morphology and systematics of Bursaphelenchus gerberae n. sp. (Nematoda: Parasitaphelenchidae), a rare associate of the palm weevil, Rhynchophorus palmarum in Trinidad. Zootaxa 2006, 1189, 39–53. [Google Scholar] [CrossRef]
- Fang, Y.; Zhuo, K.; Zhao, J. Description of Bursaphelenchus aberrans n. sp. (Nematoda: Parasitaphelenchidae) isolated from pine wood in Guangdong Province, China. Nematology 2002, 4, 791–794. [Google Scholar] [CrossRef]
- Braasch, H.; Schmutzenhofer, H. Bursaphelenchus abietinus sp. n. (Nematoda: Parasitaphelenchidae) associated with fir bark beetles (Pityokteines spp.) from declining trees in Austria. Russ. J. Nematol. 2000, 8, 1–6. [Google Scholar]
- Skarmoutsos, G.; Braasch, H.; Michalopoulou, H. Bursaphelenchus hellenicus sp. n. (Nematoda, Aphelenchoididae) from Greek pine wood. Nematologica 1998, 44, 623–629. [Google Scholar] [CrossRef]
- Massey, C.L. Biology and Taxonomy of Nematode Parasites and Associates of Bark Beetles in the United States; Agricultural Handbook No. 446; United States Department of Agriculture: Washington, DC, USA, 1974; pp. 1–233.
- Braasch, H.; Burgermeister, W. Bursaphelenchus rainulfi sp. n. (Nematoda: Parasitaphelenchidae), first record of the genus Bursaphelenchus Fuchs, 1937 from Malaysia. Nematology 2002, 4, 971–978. [Google Scholar] [CrossRef]
- Thong, C.H.S.; Webster, J.M. Nematode parasites and associates of Dendroctonus spp. and Trypodendron lineatum (Coleoptera: Scolytidae), with a description of Bursaphelenchus varicauda n. sp. J. Nematol. 1983, 15, 312–318. [Google Scholar] [PubMed]
- Penas, A.C.; Metge, K.; Mota, M.; Valadas, V. Bursaphelenchus antoniae sp. n. (Nematoda: Parasitaphelenchidae) associated with Hylobius sp. from Pinus pinaster in Portugal. Nematology 2006, 8, 659–669. [Google Scholar] [CrossRef]
- Carta, L.K.; Wick, R.L. First report of Bursaphelenchus antoniae from Pinus strobus in the U.S. J. Nematol. 2018, 50, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Trinh, P.Q.; Waeyenberge, L.; Moens, M. Bursaphelenchus chengi sp. n. (Nematoda: Parasitaphelenchidae) isolated at Nanjing, China, in packaging wood from Taiwan. Nematology 2008, 10, 335–346. [Google Scholar] [CrossRef]
- Tanaka, S.E.; Tanaka, R.; Akiba, M.; Aikawa, T.; Maehara, N.; Takeuchi, Y.; Kanzaki, N. Bursaphelenchus niphades n. sp. (Tylenchina: Aphelenchoididae) amensally associated with Niphades variegatus (Roelofs) (Coleoptera: Curculionidae). Nematology 2014, 16, 259–281. [Google Scholar] [CrossRef]
- Maria, M.; Fang, Y.; He, J.; Gu, J.F.; Li, H.M. Bursaphelenchus parantoniae n. sp. (Tylenchina: Aphelenchoididae) found in packaging wood from Belgium. Nematology 2015, 17, 1141–1152. [Google Scholar] [CrossRef]
- Kanzaki, N.; Okabe, K.; Kobori, Y. Bursaphelenchus sakishimanus n. sp. (Tylenchomorpha: Aphelenchoididae) isolated from a stag beetle, Dorcus titanus sakishimanus Nomura (Coleoptera: Lucanidae), on Ishigaki Island, Japan. Nematology 2015, 17, 531–542. [Google Scholar] [CrossRef]
- Tomalak, M.; Filipiak, A. Bursaphelenchus michalskii sp. n. (Nematoda: Aphelenchoididae), a nematode associate of the large elm bark beetle, Scolytus scolytus Fabr. (Coleoptera: Curculionidae), in Dutch elm disease-affected elm, Ulmus laevis Pall. Nematology 2019, 21, 301–318. [Google Scholar] [CrossRef]
- Zhao, Z.Q.; Davies, K.A.; Riley, I.T.; Nobbs, J.M. Laimaphelenchus heidelbergi sp. nov.(Nematoda: Aphelenchina) from Victoria, Australia, and emendment of the diagnosis of the genus. Trans. R. Soc. S. Aust. 2007, 131, 182–191. [Google Scholar] [CrossRef]
- Maleita, C.; Costa, S.R.; Abrantes, I. First report of Laimaphelenchus heidelbergi (Nematoda: Aphelenchoididae) in Europe. For. Pathol. 2014, 45, 76–81. [Google Scholar] [CrossRef]
- Carta, L.K.; Li, S.; Skantar, A.M.; Newcombe, G. Morphological and molecular characterization of two Aphelenchoides endophytic in poplar leaves. J. Nematol. 2016, 48, 28–33. [Google Scholar] [CrossRef]
- Hunt, D.J. Aphelenchida, Longidoridae and Trichodoridae: Their Systematics and Bionomics; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Kruglik, I.A. Distribution of nematodes inside trunk and branches of the dead 200 years old pine Pinus koraiensis. In Proceedings of the Fifth English Language International Symposium of the Russian Society of Nematologists, Vladivostok, Russia, 13–17 July 2003. [Google Scholar]
- Kruglik, I.A. Fauna of xylobiont nematodes of the pine Pinus densiflora from Primorye. In Proceedings of the Fifth English Language International Symposium of the Russian Society of Nematologists, Vladivostok, Russia, 13–17 July 2003. [Google Scholar]
- Gu, J.; Wang, J.L.; Braasch, H.; Burgermeister, W.; Zheng, J. Description of a new subspecies of Bursaphelenchus africanus (Nematoda: Aphelenchoididae) found in packaging wood from Russia. Nematology 2012, 14, 27–37. [Google Scholar] [CrossRef]
- Braasch, H.; Tomiczek, C.; Metge, K.; Hoyer, U.; Burgermeister, W.; Wulfert, I.; Schonfeld, U. Records of Bursaphelenchus spp. (Nematoda, Parasitaphelenchidae) in coniferous timber imported from the Asian part of Russia. For. Pathol. 2001, 31, 129–140. [Google Scholar] [CrossRef]
- Kolosova, N.V. Bursaphelenchus eroshenkii sp. n. (Nematoda: Aphelenchoididae) from the Russian Far East, with a key to some species of Bursaphelenchus Fuchs, 1937. Russ. J. Nematol. 1998, 6, 161–164. [Google Scholar]
- Kruglik, I.A.; Eroshenko, A.S. Bursaphelenchus fuchsi sp. n. (Nematoda: Bursaphelenchidae)—New nematode species from wood of pine Pinus koraiensis, Primorsky Territory. In Paraziticheskie Nematody Rastenii i Nasekomykh; Sonin, M.D., Ed.; Nauka: Moscow, Russia, 2004; pp. 96–99. [Google Scholar]
- Magnusson, C.; Kulinich, O.A. A taxonomic appraisal of original description, morphology and status of Bursaphelenchus kolymensis Korentchenko, 1980 (Aphelenchida: Aphelenchoididae). Russ. J. Nematol. 1996, 4, 155–161. [Google Scholar]
- Braasch, H. First detection of Bursaphelenchus mucronatus Mamiya & Enda, 1979 in Germany and its presence in wood imported from USSR, with further details on the description of the species. Arch. Phytopathol. Pflanzenschutz Berl. 1991, 27, 209–218. [Google Scholar]
- Kulinich, O.A.; Kruglik, A.; Eroshenko, A.; Kolosova, N. Ocurrence and distribution of the nematode Bursaphelenchus mucronatus in the Russian Far East. Russ. J. Nematol. 1994, 2, 113–119. [Google Scholar]
- Wang, X.; Fang, Y.; Maria, M.; Gu, J.; Ge, J. Description of Bursaphelenchus rockyi n. sp. (Nematoda: Aphelenchoididae) in peat moss from Russia. Nematology 2019, 21, 253–265. [Google Scholar] [CrossRef]
- Kruglik, I.A. Xylobiont Nematodes of Pines in Primorsky Territory. Ph.D. Thesis, Biologo-Pochvennyi Institut, Far East Scientific Center of the Russian Academy of Sciences, Vladivostok, Russia, 2003. [Google Scholar]
- Khusainov, R.V. Diversity and Ecology of Xylobionts Aphelenchidae (Nematoda: Aphelenchina) of Various Ecosystems of Tula Region. In Bulletin of Tula State University. Interregional Scientific Conference “Studying and Preserving Biodiversity of the Tula Region and Neighboring Regions of the Russian Federation”, Dedicated to the 120th Anniversary of G. N. Likhachev. 20–22 November 2019 Tula; Shiryaev, K.A., Ed.; TulSU Publishing House: Maharashtra, India, 2019; pp. 45–47. [Google Scholar]
- Yu, Q.; Gu, J.; Ye, W.; Li, R.; He, J. Deladenus posteroporus n. sp. (Nematoda: Neotylenchidae) isolated from packaging wood from Canada and white pine (Pinus monticola) Lumber from the United States and Intercepted in Ningbo, China. J. Nematol. 2017, 49, 168–176. [Google Scholar] [CrossRef]
- Miraeiz, E.; Heydari, R.; Golhasan, B. A new and a known species of Deladenus Thorne, 1941 (Nematoda: Neotylenchidae) from Iran, with an updated species checklist of the genus. Acta Zool. Bulg. 2017, 69, 307–316. [Google Scholar]
- Chizhov, V.N.; Butorina, N.N.; Tabolin, S.B.; Limantseva, L.A.; Subbotin, S.A. Entomoparasitic Nematodes of the Orders Tylenchida and Aphelenchida; KMK Publishers: Moscow, Russia, 2019. [Google Scholar]
- Blinova, S.L.; Korentchenko, E.A. Phaenopsitylenchus laricis gen. n., sp. n. (Nematoda: Phaenopsitylenchidae fam. n.—A parasite of Phaenops guttulata, and notes to the taxonomy of the nematode superfamily Sphaerularioidea. Proc. Helminthol. Lab. AN SSSR 1986, 34, 14–23. [Google Scholar]
- Kanzaki, N.; Sakamoto, H.; Maehara, N. Diplogasteroides nix n. sp. (Nematoda: Diplogastridae), a cryptic species related to D. andrassyi, isolated from Monochamus urussovi (Coleoptera: Cerambycidae) from Hokkaido, Japan, with remarks on body surface structures. Nematology 2016, 18, 753–773. [Google Scholar] [CrossRef]
- Vosilite, B.S. A new species of nematode, Diplogasteroides sexdentati n. sp. and notes on biology of the ectoparasitic nematodes of Ips sexdentatus. In Gel’minty Nasekomykh; Sonin, M.D., Ed.; Nauka: Moscow, Russia, 1980; pp. 21–28. [Google Scholar]
- Miraeiz, E.; Heydari, R.; Maafi, Z.T.; Bert, W. Laimaphelenchus hyrcanus n. sp. (Nematoda: Aphelenchoididae), a new species from northern Iran. Zootaxa 2015, 3915, 591–600. [Google Scholar] [CrossRef]
- Oro, V. Description of Laimaphelenchus belgradiensis sp. nov. (Nematoda: Aphelenchoididae) and its phylogenetic and system- atic position within Aphelenchoidoidea. Eur. J. Plant Pathol. 2015, 142, 13–23. [Google Scholar] [CrossRef]
- Maleita, C.M.N.; Abrantes, I.M.O. On the species status of Laimaphelenchus hyrcanus and L. belgradiensis. For. Pathol. 2018, 48, e12425. [Google Scholar] [CrossRef]
- Eroshenko, A.S.; Volkova, T.V. Plant Parasitic Nematodes of the Far East of Russia; Dal’nauka: Vladivostok, Russia, 2005; pp. 1–226. [Google Scholar]
- Rogozhin, E.A.; Chizhov, V.N.; Ryss, A.Y.; Nasonova, L.V.; Kulinich, O.A. Fauna of xylobiont nematodes in the Central region of the Non-Black-Soil zone of the Russian Federation. In Proceedings of the VII International Nematological Symposium, Petrazavodsk, Russia, 9–14 July 2007; Karelian Scentific Center of the Russian Academy of Scences: Petrazavodsk, Russia, 2007; pp. 80–81. [Google Scholar]
- Ryss, A.Y. Classification, Morphology and Evolution of the Plant Parasitic Nematodes of the Families Pratylenchidae (Order Tylenchida), Aphelenchoididae and Parasitaphelenchidae (Order Aphelenchida). Ph.D. Thesis, Zoological Institute, Russian Academy of Sciences, St. Petersburg, Russia, 2008; pp. 1–50. [Google Scholar]
- Susoy, V.; Kanzaki, N.; Herrmann, M. Description of the bark beetle associated nematodes Micoletzkya masseyi n. sp. and M. japonica n. sp. (Nematoda: Diplogastridae). Nematology 2013, 15, 213–231. [Google Scholar] [CrossRef]
- Kanzaki, N.; Ekino, T.; Ide, T.; Masuya, H.; Degawa, Y. Three new species of parasitaphelenchids, Parasitaphelenchus frontalis n. sp., P. costati n. sp. and Bursaphelenchus hirsutae n. sp. (Nematoda: Aphelenchoididae), isolated from bark beetles from Japan. Nematology 2018, 20, 957–1005. [Google Scholar] [CrossRef]
- Carta, L.K.; Bauchan, G.; Hsu, C.Y.; Yuceer, C. Description of Parasitorhabditis frontali n. sp. (Nemata: Rhabditida) from Dendroctonus frontalis Zimmermann (Coleoptera: Scolytidae). J. Nematol. 2010, 42, 46–54. [Google Scholar]
- Blinova, S.L.; Gurando, E.V. Parasitorhabdidtis fuchsi sp. n. (Nematoda: Rhabditidae—A parasite of the Blastophagus minor Hartig. Vestn. Zool. 1974, 2, 50–55. [Google Scholar]
- Skrjabin, K.I.; Shikhobalova, N.P.; Sobolev, A.A.; Paramonov, A.A.; Sudarikov, V.E. Camallanata, Rhabditata, Tylenchata, Trichocephalata, Dioctophymata and the Distribution of the Parasitic Nematodes in Their Hosts; Nauka: Moscow, Russia, 1954; 827p. [Google Scholar]
- Andrássy, I. Klasse Nematoda (Ordnungen Monhysterida, Desmoscolecida, Araeolaimida, Chromadorida, Rhabditida); Akademie-Verlag: Berlin, Germany, 1984; 509p. [Google Scholar]
- Blinova, S.L.; Vosilite, B.S. Study of individual and geographic variability of the nematode Parasitorhabditis sexdentati (Rhabditidae), parasite of the weevil Ips sexdentatus. Tr. Gel’mintologicheskoi Lab. AN SSSR 1974, 24, 25–29. [Google Scholar]
- Lodge, D.M.; Simonin, P.W.; Burgiel, S.W.; Keller, R.P.; Bossenbroek, J.M.; Jerde, C.L.; Kramer, A.M.; Rutherford, E.S.; Barnes, M.A.; Wittmann, M.E.; et al. Risk analysis and bioeconomics of invasive species to inform policy and management. Annu. Rev. Environ. Resour. 2016, 41, 453–488. [Google Scholar] [CrossRef]
- Mota, M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Jones, J.T.; Moens, M.; Mota, M.; Li, H.; Kikuchi, T. Bursaphelenchus xylophilus: Opportunities in comparative genomics and molecular host-parasite interactions. Mol. Plant Pathol. 2008, 9, 357–368. [Google Scholar] [CrossRef]
- Mallez, S.; Castagnone, C.; Lombaert, E.; Castagnone-Sereno, P.; Guillemaud, T. Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysis. bioRxiv 2021, 452326. [Google Scholar] [CrossRef]
- Kanzaki, N.; Ekino, T.; Giblin-Davis, R.M. Feeding dimorphism in a mycophagous nematode, Bursaphelenchus sinensis. Sci. Rep. 2019, 9, 13956. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, N.; Futai, K. Isolation of Bursaphelenchus sinensis (Nematoda: Parasitaphelenchidae) from dead Japanese black pine, Pinus thunbergii Parl. in Japan. J. Nematode Morphol. Syst. 2007, 9, 129–136. [Google Scholar]
- Han, H.; Kim, H. First Report of Bursaphelenchus sinensis on Pinus densiflora in South Korea. Plant Dis. 2020, 104, 596. [Google Scholar] [CrossRef]
- Qin, F.N.; Pan, C.S. Studies on pine-parasitic nematodes in Fujian. I. Species of Bursaphelenchus genus on pine stem. J. Xiamen Univ. (Nat. Sci.) 2005, 44, 723–728. [Google Scholar]
- Braasch, H.; Braasch-Bidasak, R. First record of the genus Bursaphelenchus Fuchs, 1937 in Thailand and description of B. thailandae sp. n. (Nematoda: Parasitaphelenchidae). Nematology 2002, 4, 853–863. [Google Scholar] [CrossRef]
- Tomiczek, C.; Braasch, H.; Burgermeister, W.; Metge, K.; Hoyer, U.; Brandstetter, M. Identification of Bursaphelenchus spp. isolated from Chinese packaging wood imported to Austria. Nematology 2003, 5, 573–581. [Google Scholar] [CrossRef]
- Kanzaki, N.; Giblin-Davis, R.M. Diversity and plant pathogenicity of Bursaphelenchus and related nematodes in relation to their vector bionomics. Curr. For. Rep. 2018, 4, 85–100. [Google Scholar] [CrossRef]
- Kanzaki, N.; Masuya, H.; Ichihara, Y.; Maehara, N.; Aikawa, T.; Ekino, T.; Ide, T. Bursaphelenchus carpini n. sp., B. laciniatae n. sp. and B. cryphali okhotskensis n. subsp. (Nematoda: Aphelenchoididae) isolated from Japan. Nematology 2019, 21, 361–388. [Google Scholar] [CrossRef]
- Rühm, W. Die Nematoden der Ipiden. Parasitol. Schr. 1956, 6, 1–435. [Google Scholar]
- Kurashvili, B.E.; Kakulia, G.A.; Devdariani, T.G. Parasitic Nematodes of the Bark-Beetles in Georgia; Metsniereba: Tbilisi, Georgia, 1980. [Google Scholar]
- Kubatova, A.; Novotny, D.; Prasil, K.; Mrácek, Z. The nematophagous hyphomycete Esteya vermicola found in the Czech Republic. Czech Mycol. 2000, 52, 227–235. [Google Scholar] [CrossRef]
- Schonfeld, U.; Braasch, H.; Burgermeister, W. Bursaphelenchus spp. (Nematoda: Parasitaphelenchidae) in wood chips from sawmills in Brandenburg and description of Bursaphelenchus willibaldi sp. n. Russ. J. Nematol. 2006, 14, 119–126. [Google Scholar]
- Calin, M.; Vieira, P.; Costache, C.; Braasch, H.; Gu, J.; Wang, J.; Mota, M. Survey of the genus Bursaphelenchus Fuchs, 1937 (Nematoda: Aphelenchoididae) in Romania. EPPO Bull. 2013, 43, 144–151. [Google Scholar] [CrossRef]
- Miraeiz, E.; Heydari, R.; Aliramaji, F.; Pourjam, E.; Pedram, M.; Atighi, M.R. New morphometric ranges for Bursaphelenchus willibaldi Schönfeld et al. 2006 (Nematoda: Parasitaphelenchidae) from Iran. For. Pathol. 2016, 46, 29–34. [Google Scholar] [CrossRef]
- Ryss, A.; Vieira, P.; Mota, M.; Kulinich, O. A synopsis of the genus Bursaphelenchus Fuchs, 1937 (Aphelenchida: Parasitaphelenchidae) with keys to species. Nematology 2005, 7, 393–458. [Google Scholar] [CrossRef]
- Ryss, A.Y. The simplest “field” methods for extraction of nematodes from plants, wood, insects and soil with additional description how to keep extracted nematodes alive for a long time. Parazitologiya 2017, 51, 57–67. [Google Scholar]
- Ryss, A.Y.; Polyanina, K.S. The development and population dynamics of Aphelenchoides sp. from Quercus robur (Nematoda: Rhabditida: Aphelenchoididae) in vitro. Russ. J. Nematol. 2022, 31, 1–15. [Google Scholar] [CrossRef]
- Ryss, A.Y. The most simple methods to detect the wood-inhabiting nematodes and the laboratory cultivation. Proc. St. Petersburg For. Tech. Acad. 2015, 2011, 287–295. [Google Scholar]
- Ryss, A.Y. A Simple express technique to process nematodes for collection slide mounts. J. Nematol. 2017, 49, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, S.A. Molecular identification of nematodes using polymerase chain reaction (PCR). In Techniques for Work with Plant and Soil Nematodes; Perry, R.N., Hunt, D.J., Subbotin, S.A., Eds.; CAB International: Wallingford, UK, 2021; pp. 218–239. [Google Scholar]
- Launay, C.; Félix, M.A.; Dieng, J.; Delattre, M. Diversification and hybrid incompatibility in auto-pseudogamous species of Mesorhabditis nematodes. BMC Evol. Biol. 2020, 20, 105. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, S.A. Phylogenetic analysis of DNA sequence data. In Techniques for Work with Plant and Soil Nema-Todes; Perry, R.N., Hunt, D.J., Subbotin, S.A., Eds.; CAB International: Wallingford, UK, 2021; pp. 265–282. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample Code | Location | GPS Coordinates | Associated Plant/Insect | GenBank Accession Number |
|---|---|---|---|---|---|
| Aphelenchoides heidelbergi | CD366 | Saint Petersburg, Bol’shoy Prospekt Vasil’yevskogo Ostrova 25 | 59.939251; 30.281787 | Quercus robur/Scolytus sp. | OP854773 |
| A. heidelbergi | CD3663 | Republic of Dagestan, Samurskiy Les | 41.844639; 48.544833 | Elaeagnus latifolia/Scolytus jaroschewskii | OP854774 |
| Aphelenchoides sp.1 | CD3645 | Saint Petersburg, Bol’shoy Prospekt Vasil’yevskogo Ostrova | 59.939251; 30.281787 | Q. robur/Scolytus sp. | OP854771 |
| Aphelenchoides sp.1 | CD3647 | Saint Petersburg, Isaakiyevskiy Skver | 59.932579; 0.307305 | Q. robur/Scolytus sp. | OP854772 |
| Aphelenchoides sp.2 | CD3742 | Nizhny Novgorod | - | Betula sp./- | OP854770 |
| Bursaphelenchuseremus | CD2936 | Nizhny Novgorod, Strigino | 56.198483; 43.761133 | Q. robur/Scolytus intricatus; Plagiontus sp. | OP854775 |
| B. fraudulentus | CD2935 | Moscow, Main Botanical Garden of the Russian Academy of Sciences | 55.841033; 37.607117 | Q. robur/Cerambycidae | OP854779 |
| B. fraudulentus | CD3644 | Belarus, Gomel Region | 52.478967; 29.419450 | Q. robur/Cerambycidae | OP854780 |
| B. michalskii | CD3643 | Republic of Dagestan, Samurskiy Les | 41.844639; 48.544833 | Ulmus minor/S. jaroschewskii | OP854776 |
| B. mucronatus | CD3642 | Buryatia, Baikal Nature Reserve | 51.415833; 104.821794 | Abies sibirica/Monochamus galloprovincialis | OP854781 |
| B. willibaldi | CD3664 | Nizhny Novgorod, Botanical Garden of Lobachevsky University | 56.254267; 44.006583 | Q. robur/S. intricatus | OP854782 |
| B. zvyagintsevi sp. n. | CD3665 | Khabarovsk Krai, Arboretum of the Far Eastern Research Institute of Forestry | 48.463611; 135.084444 | Fraxinus mandshurica/Hylesinus laticollis | OP854777 |
| Cryptaphelenchus sp.1 | CD3655 | Tomsk region, Luchanovo- Ipatovsky Settlement Cedar Forest | 56.333228; 85.050120 | Pinus sibirica/Ips sexdentatus | OP854784 |
| Cryptaphelenchus sp.2 | CD3657 | Tomsk region, Malinovka | 56.696938; 85.362820 | A. sibirica/Polygraphus proximus | OP854785 |
| Cryptaphelenchus sp.3 | CD3658 | Tomsk region, Zavarzino | 56.472507; 85.092819 | A. sibirica/P. proximus | OP854783 |
| Deladenus posteroporus | CD3652 | Magadan region, Omsukchansky District | 61.180106; 153.900808 | Larix sibirica/Scolytinae | OP854789 |
| Deladenus posteroporus | CD3668 | Magadan region, Omsukchansky District | 61.180106; 153.900808 | L. sibirica/Scolytinae | OP854790 |
| Diplogasteroides nix | CD3673 | Magadan region, Omsukchansky District | 61.180106; 153.900808 | L. sibirica/Scolytinae | OP854796 |
| Laimaphelenchus hyrcanus | CD3646 | Saint Petersburg, Isaakiyevskiy Skver | 59.932579; 30.307305 | Q. robur/Scolytus sp. | OP854786 |
| Laimaphelenchus sp.1 | CD3667 | Saint Petersburg, Russian Museum, Mikhailovsky Garden | 59.940949; 30.332522 | A. platanoides/Scolytinae | OP854787 |
| Laimaphelenchus sp.1 | CD3674 | Saint Petersburg, Russian Museum, Mikhailovsky Garden | 59.939623; 30.329235 | Tilia cordata/Scolytus sp. | OP854788 |
| Micoletzkya sp.1 | CD3672 | Buryatia, Baikal Nature Reserve | 51.563500; 105.394778 | A. sibirica/P. proximus | OP854795 |
| Parasitaphelenchus sp.1 | CD3649 | Krasnoyarsk | 56.103451; 92.959464 | A. sibirica/Ips sp. | OP854778 |
| Parasitorhabditis sp.1 | CD3671 | Buryatia, Baikal Nature Reserve | 51.563500; 105.394778 | A. sibirica/P. proximus | OP854794 |
| Sphaerulariid nematode | CD3653 | Saint Petersburg, Ekaterinhof Park | 59.902938; 30.267495 | Lindera angustifolia/Scolytinae | OP854791 |
| Tylenchid nematode sp.1 | CD3666 | Saint Petersburg, Russian Museum, Mikhailovsky Garden | 59.940617; 30.332185 | T. cordata/Scolytus sp. | OP854793 |
| Tylenchid nematode sp.2 | CD3648 | Saint Petersburg, Russian Museum, Mikhailovsky Garden | 59.939513; 30.328864 | U. laevis/S. multistriatus | OP854792 |
| Species | Bursaphelenchus zvyagintsevi sp. n. | Bursaphelenchus michalskii Dagestan Population | ||||
|---|---|---|---|---|---|---|
| Character | Holotype Male | Male | Female | Male | Female | |
| n | 1 | 20 | 20 | 20 | 20 | |
| L | 562 | 545 ± 65 (470–676) | 566 ± 47 (484–625) | 901 ± 119 (699–1052) | 1035 ± 164 (817–1277) | |
| a | 34.1 | 35.2 ± 2.4 (33.1–40.2) | 32.1 ± 2.6 (28.0–36.7) | 64.4 ± 7.0 (53.8–73.7) | 56.3 ± 6.3 (50.2–65.5) | |
| b | 9.9 | 9.4 ± 1.5 (8.3–12.8) | 9.0 ± 0.9 (7.9–10.3) | 13.9 ± 1.2 (11.7–15.4) | 15.5 ± 1.7 (13.0–18.2) | |
| b’ | 4.7 | 4.3 ± 0.7 (3.8–5.7) | 4.3 ± 0.4 (3.7–4.7) | 5.9 ± 0.6 (5.0–6.9) | 6.3 ± 0.7 (5.4–7.3) | |
| c | 17.6 | 19.1 ± 2.1 (17.1–22.5) | 18.7 ± 1.2 (17.3–20.4) | 31.1 ± 3.3 (28.0–36.3) | 23.7 ± 2.2 (19.2–25.5) | |
| c’ | 2.3 | 2.0 ± 0.2 (1.7–2.3) | 3.5 ± 0.4 (3.1–4.0) | 2.3 ± 0.2 (2.1–2.6) | 4.6 ± 0.4 (4.1–5.1) | |
| V (%) | - | - | 73 ± 2 (70–76) | - | 71.4 ± 0.9 (70.1–72.7) | |
| Stylet | 14 | 13.7 ± 1.1 (12–15) | 13.6 ± 0.7 (12–15) | 13 ± 1 (11–14) | 14.0 ± 1.3 (12.0–15.5) | |
| Stylet cone/stylet (%) | 43 | 47 ± 5 (39–54) | 49 ± 4 (43–56) | 50 ± 4 (45–56) | 52 ± 5 (45–59) | |
| Cephalic region diam. | 6.5 | 5.5 ± 0.9 (4.0–6.5) | 6.1 ± 0.6 (5.0–7.0) | 6 ± 0.5 (5–6.5) | 6.5 ± 0.5 (6–7) | |
| Cephalic region height | 3.0 | 2.8 ± 0.4 (2.5–3.5) | 2.9 ± 0.5 (2.0–3.5) | 3.1 ± 0.3 (2.5–3.5) | 3.3 ± 0.3 (3–3.5) | |
| Median bulb length (L) | 16 | 14.8 ± 0.9 (13–16) | 16.6 ± 1.4 (15.0–19.0) | 14.3 ± 1.4 (13.0–16.5) | 14 ± 2.9 (11–18) | |
| Median bulb diam. (D) | 10 | 9.8 ± 0.7 (9–11) | 11.0 ± 0.9 (10.0–12.0) | 8.5 ± 1.5 (6.0–10.0) | 9.4 ± 1.0 (8–11) | |
| Median bulb L/D | 1.6 | 1.5 ± 0.2 (1.2–1.7) | 1.5 ± 0.1 (1.4–1.7) | 1.7 ± 0.3 (1.4–2.3) | 1.5 ± 0.2 (1.2–1.7) | |
| Excretory pore from anterior | 58 | 66 ± 5 (58–71) | 73 ± 5 (65–80) | 71 ± 10 (57–84) | 80 ± 7 (71–89) | |
| Nerve ring posterior border from anterior | 78 | 74 ± 4 (70–78) | 78 ± 3 (72–83) | 81 ± 7 (74–92) | 85 ± 9 (74–99) | |
| Pharynx | 57 | 58.3 ± 3.4 (53–62) | 63 ± 4 (57–71) | 65 ± 5 (57–73) | 67 ± 6 (59–76) | |
| Anterior to gland lobe end | 120 | 127.9 ± 7.7 (119–137) | 131 ± 4 (124–138) | 154 ± 14 (135–173) | 163 ± 22 (141–192) | |
| Gland lobe | 63 | 69.6 ± 4.7 (63–76) | 68 ± 6 (57–75) | 89 ± 11 (75–103) | 97 ± 16 (81–118) | |
| Gland lobe/body diam. at median bulb | 4.3 | 5.1 ± 0.6 (4.1–5.8) | 4.7 ± 0.6 (3.8–5.4) | 7.1 ± 0.8 (5.9–8.8) | 7.1 ± 0.9 (6.0–8.4) | |
| Body diameter (BW) | 16.5 | 15 ± 1 (13–17) | 18 ± 2 (15–21) | 14.1 ± 1.9 (12.0–16.5) | 18.4 ± 2.5 (13.5–21) | |
| PUS | - | - | 82 ± 10 (62–98) | - | 179 ± 20 (140–200) | |
| Vulva–anus distance | - | - | 123 ± 14 (105–144) | - | 251 ± 36 (208–298) | |
| Vagina | - | - | 11 ± 1 (10–13) | - | 11.6 ± 1.3 (9.5–12.5) | |
| PUS/BW at vulva | - | - | 4.7 ± 0.8 (3.6–5.8) | - | 9.8 ± 0.6 (9.1–10.6) | |
| PUS/vulva–anus distance (%) | - | - | 67 ± 9 (53–83) | - | 72 ± 7 (63–81) | |
| Tail | 32 | 29 ± 3 (25–32) | 30 ± 2 (27–33) | 29 ± 3 (24–34) | 44 ± 5 (36–50) | |
| Tail diam. at anus | 14 | 14 ± 1 (13–16) | 9 ± 1 (8–10) | 13 ± 1 (12–15) | 9 ± 1 (8.0–10.5) | |
| Annuli (width of 10 at mid-body) | 8.0 | 8.0+0.9 (7.0–9.5) | 8.8 ± 0.8 (7.5–10.0) | 10.9+2.4 (8.0–14.0) | 11.1+2.0 (8.0–14.5) | |
| Spicule length (midline arc) | 16.5 | 16.5 ± 0.8 (15–17.5) | - | 13.7 ± 1.3 (12.4–15.5) | - | |
| Spicule length (dorsal lamina arc) | 19 | 19.1 ± 0.7 (18–20) | - | 17.6 ± 1.0 (16–19) | - | |
| Spicule length (ventral lamina arc) | 17 | 15.3 ± 1.4 (13.7–17) | - | 12.2 ± 1.9 (10–15) | - | |
| Distance between ends of condylus and rostrum | 6.5 | 6.5 ± 0.8 (5–7.5) | - | 7.7 ± 0.9 (6.3–8.8) | - | |
| Spicule width posterior to rostrum (lateral view) | 2.5 | 2.6 ± 0.3 (2–3) | - | 3.5 ± 0.4 (2.8–4.2) | - | |
| Angle between lines: along capitulum (condylus–rostrum) and extending spicule end (dorsal intersection) | 13 | 17.4 ± 3.1 (13–23)° | - | 6.0 ± 2.2 (3–9)° | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryss, A.Y.; Subbotin, S.A. New Records of Wood- and Bark-Inhabiting Nematodes from Woody Plants with a Description of Bursaphelenchus zvyagintsevi sp. n. (Aphelenchoididae: Parasitaphelenchinae) from Russia. Plants 2023, 12, 382. https://doi.org/10.3390/plants12020382
Ryss AY, Subbotin SA. New Records of Wood- and Bark-Inhabiting Nematodes from Woody Plants with a Description of Bursaphelenchus zvyagintsevi sp. n. (Aphelenchoididae: Parasitaphelenchinae) from Russia. Plants. 2023; 12(2):382. https://doi.org/10.3390/plants12020382
Chicago/Turabian StyleRyss, Alexander Y., and Sergei A. Subbotin. 2023. "New Records of Wood- and Bark-Inhabiting Nematodes from Woody Plants with a Description of Bursaphelenchus zvyagintsevi sp. n. (Aphelenchoididae: Parasitaphelenchinae) from Russia" Plants 12, no. 2: 382. https://doi.org/10.3390/plants12020382