
Mining Salt Tolerance SNP Loci and Prediction of Candidate Genes in the Rice Bud Stage by Genome-Wide Association Analysis


Abstract
:1. Introduction
2. Results
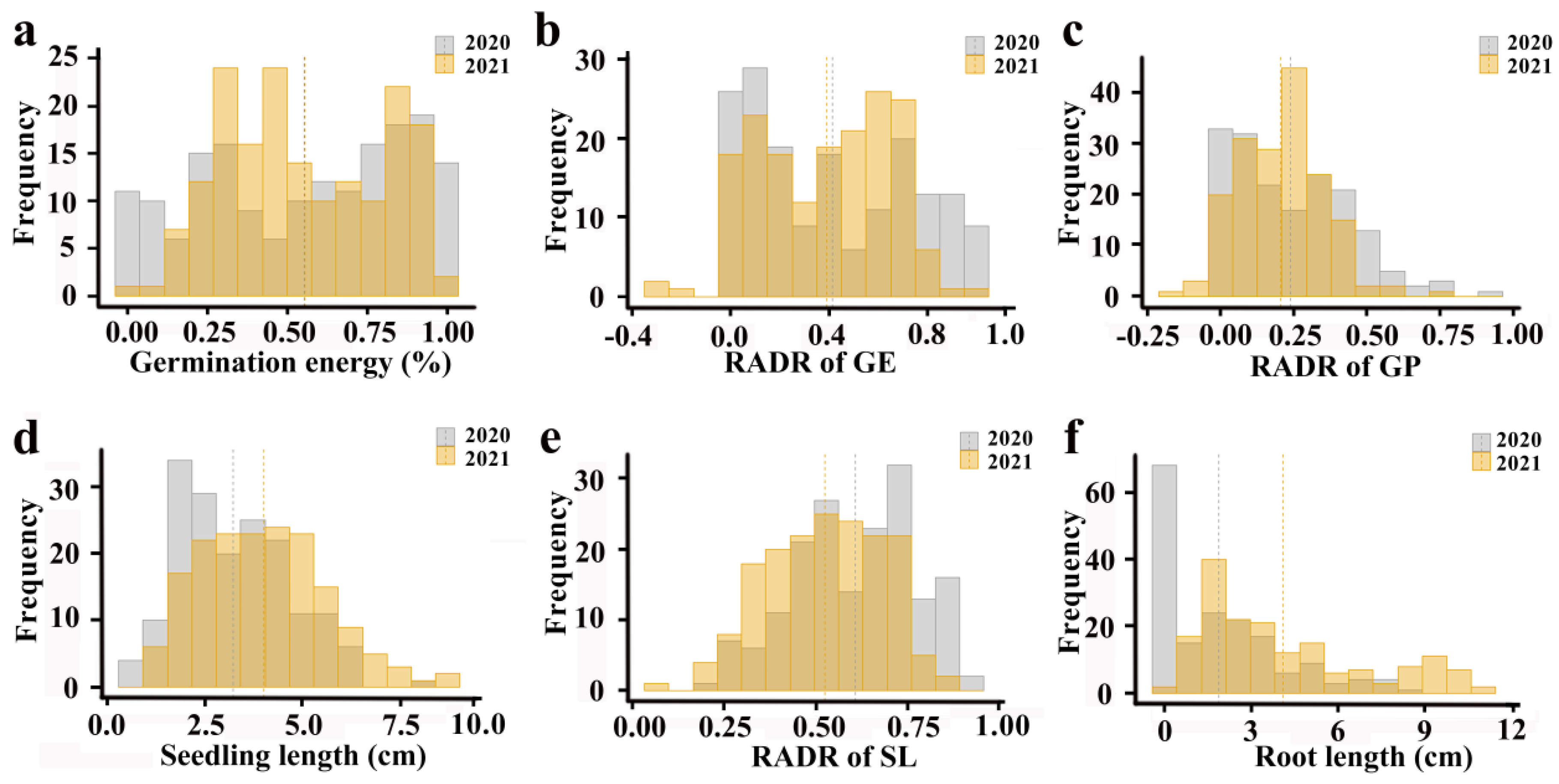
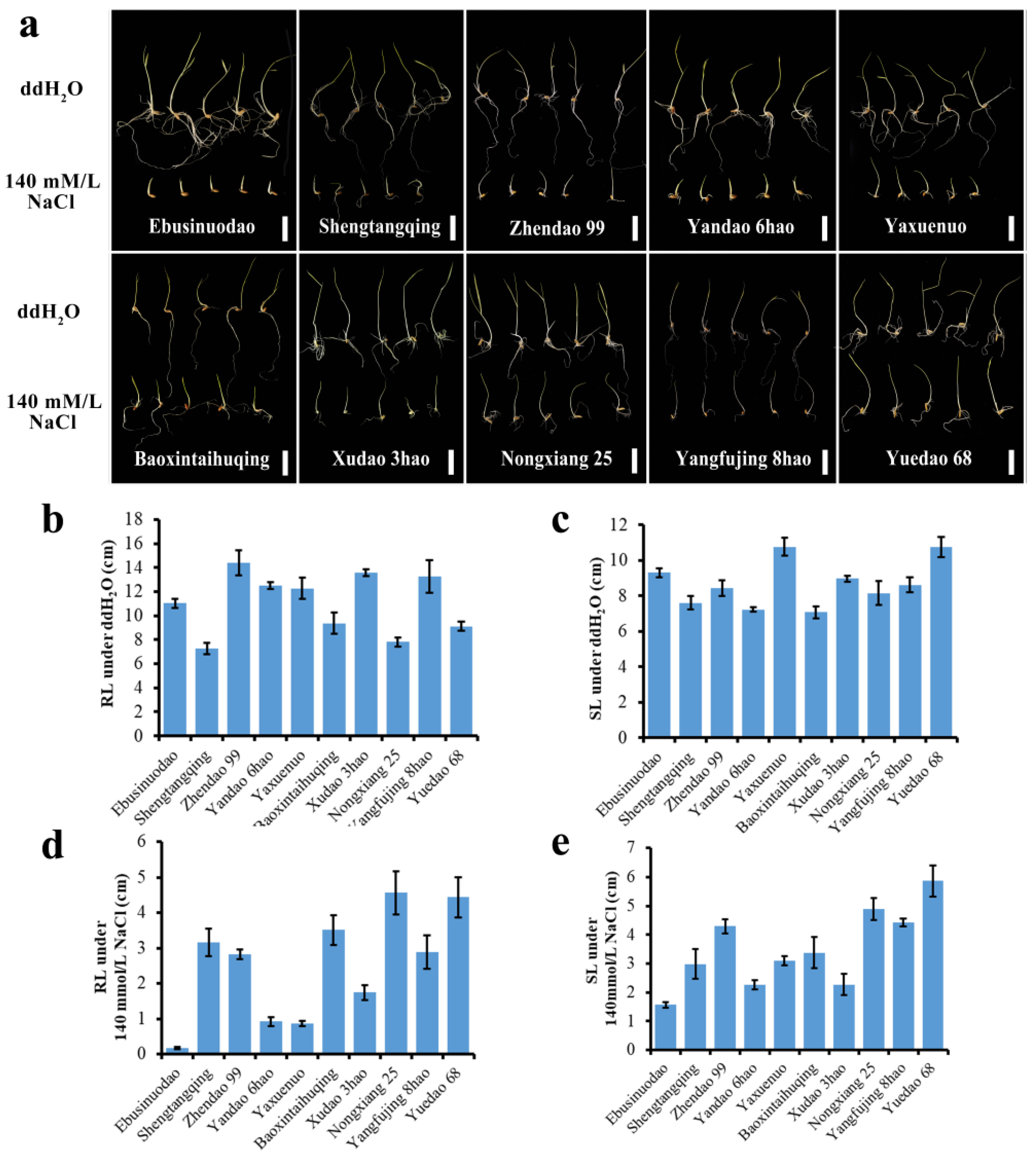
2.1. Abundant Phenotypic Variation in Salt Tolerance-Related Traits in 173 Accessions
2.2. Identification of 2 Subgroups of the 173 Accessions
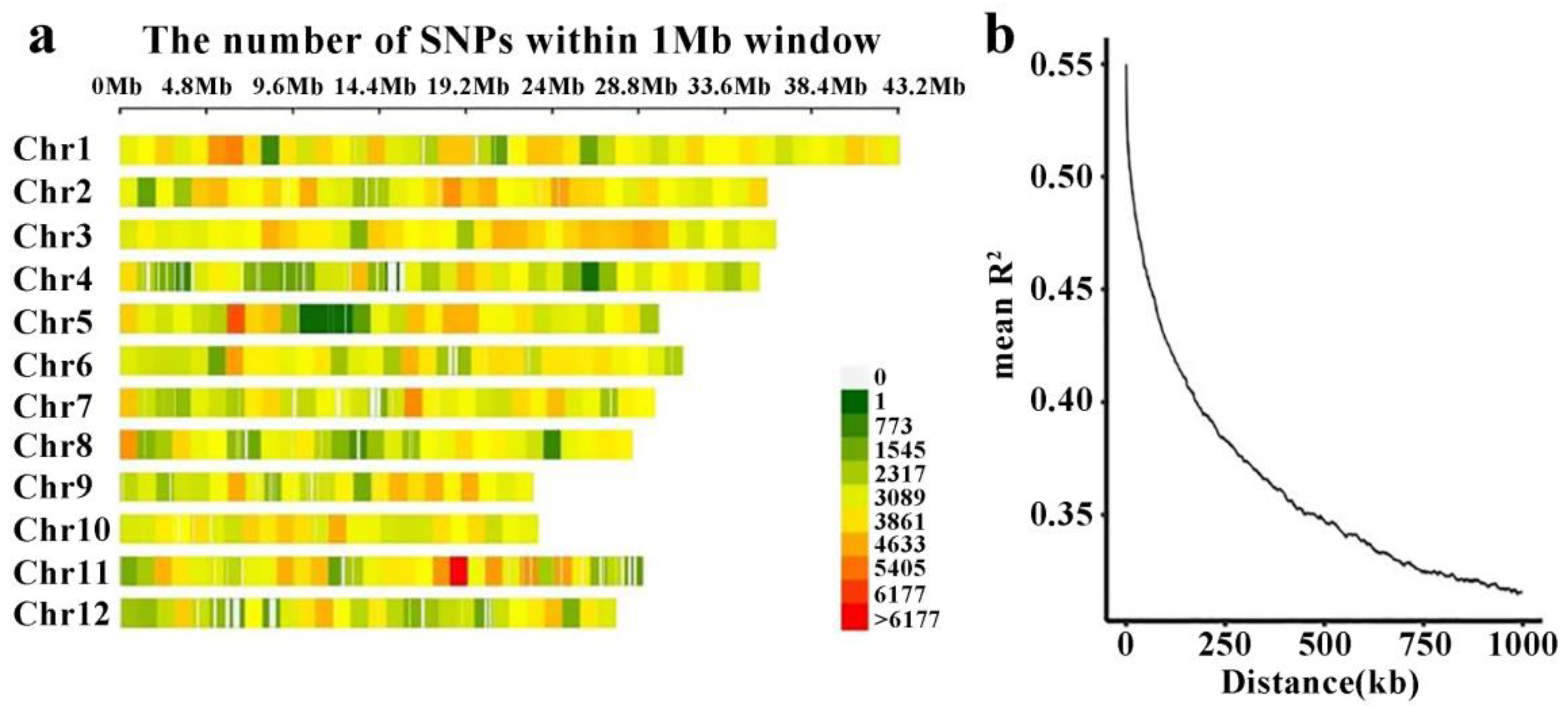
2.3. Linkage Disequilibrium
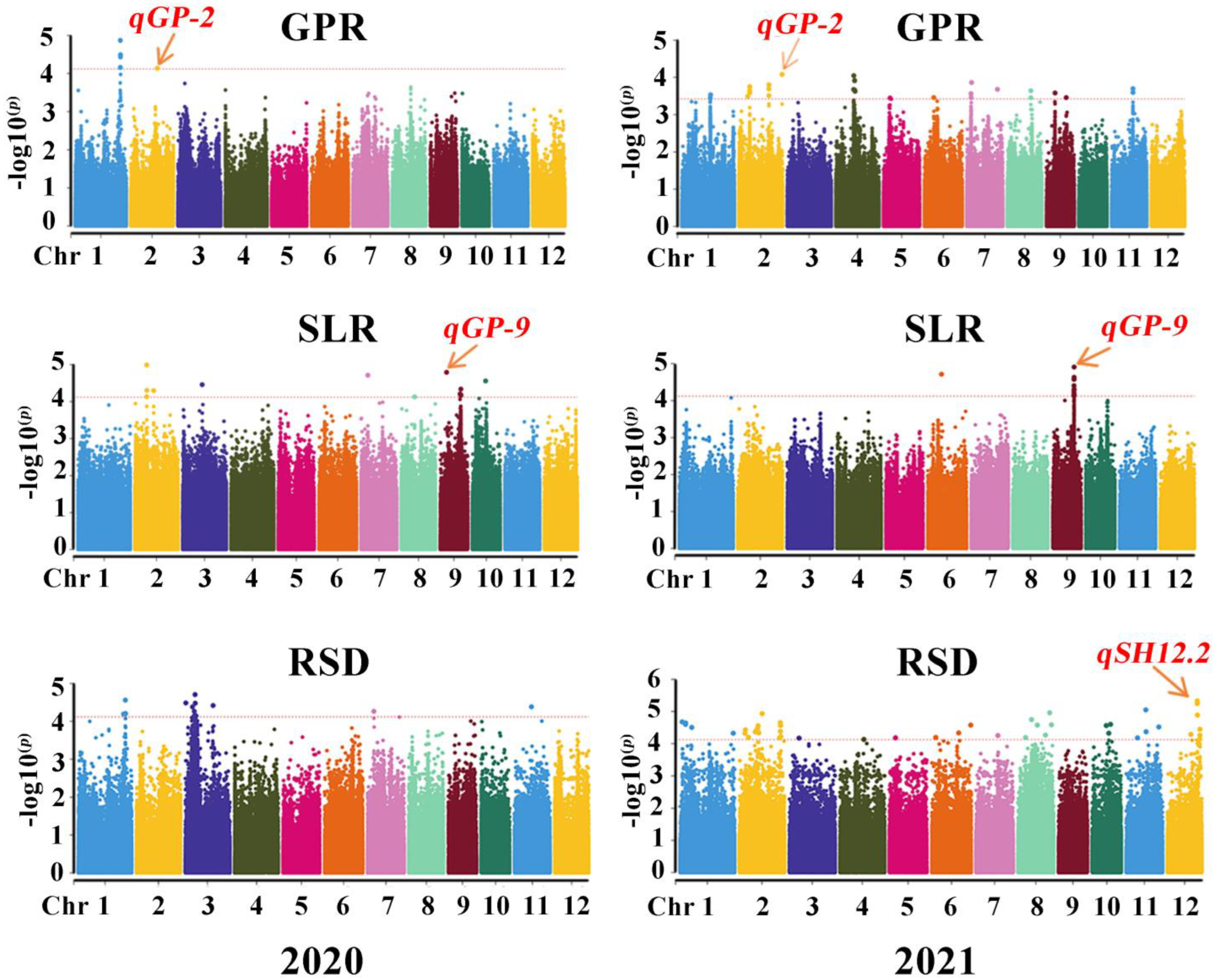
2.4. Twelve SNP Loci Related to Salt Tolerance Detected in Two Study Years
2.5. Prediction of Salt-Tolerant Candidate Genes at the Germination Stage
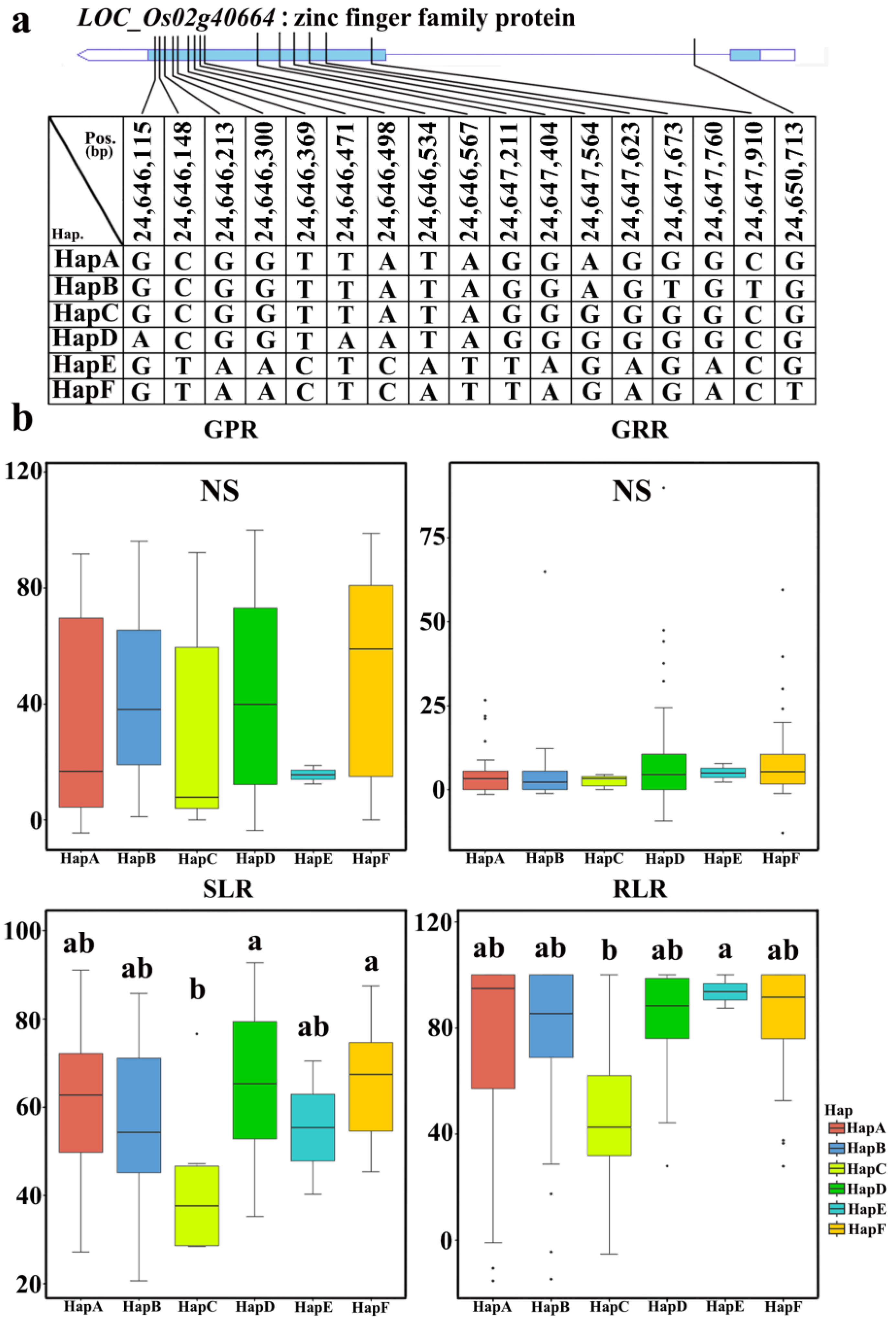
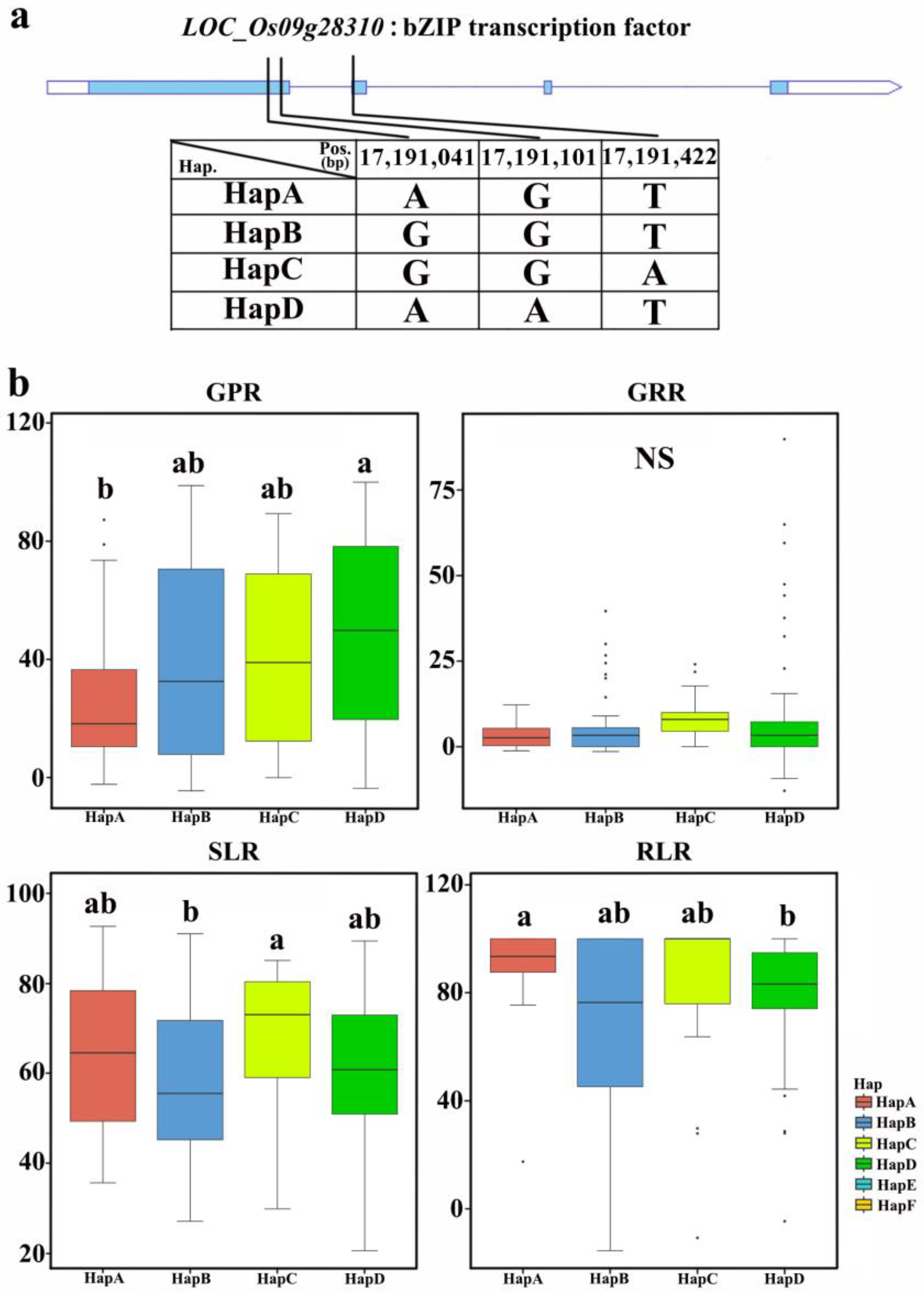
2.6. Elite Haplotype Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Field Planting and Management
4.3. Identification of Salt Tolerance
- GP (%) = (number of germinated grains on the 4th day/total number of grains) × 100%
- GR (%) = (number of germinated grains on the 10th day/total number of grains) × 100%
- GPR (%) = (control germination potential-treatment germination potential)/control germination potential × 100%
- GRR (%) = (control germination rate-treatment germination rate)/control germination rate × 100%
- SLR (%) = (control seedling length-treated seedling length)/control seedling length × 100%
- RLR (%) = (control root length-treated root length)/control root length × 100%
- RSD (%) = (GPR + GRR)/2
- CRS (%) = (SLR + RLR)/2
4.4. Genotype Identification
4.5. Phenotypic Data Analysis
4.6. Structure Analysis
4.7. Linkage Disequilibrium Analysis
4.8. Genome-Wide Association Analysis
4.9. Candidate Gene Prediction
4.10. Elite Haplotype Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreira, M.H.; They, N.H.; Rodrigues, L.R.; Alvarenga-Lucius, L.; Pita-Barbosa, A. Salty freshwater macrophytes: The effects of salinization in freshwaters upon non-halophyte aquatic plants. Sci. Total. Environ. 2023, 857, 159608. [Google Scholar] [CrossRef]
- Qin, H.; Li, Y.; Huang, R. Advances and Challenges in the Breeding of Salt-Tolerant Rice. Int. J. Mol. Sci. 2020, 21, 8385. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Dou, Y.; Xie, L.; Xie, X. Research progress on the genes related to salt tolerance in rice. Mol. Plant Breed. 2017, 15, 4411–4422. [Google Scholar] [CrossRef]
- Ma, C.; Ma, L.; Liu, T.; Zou, H.; Zhang, B.; Liu, Y. Research progress on saline land improvement technology. World For. Res. 2010, 23, 28–32. [Google Scholar] [CrossRef]
- Guo, F. Screening and Evaluation of Salt Tolerance Germplasm in Rice. Master’s Thesis, Yangzhou University, Yangzhou, China, 2018. Chapter 3. pp. 13–29. [Google Scholar]
- Shi, X.; Wang, H.; Song, J.; Lv, X.; Li, W.; Li, B.; Shi, J. Impact of saline soil improvement measures on salt content in the abandonment-reclamation process. Soil. Tillage Res. 2021, 208, 104867. [Google Scholar] [CrossRef]
- Zhu, H. Application of QTL technology in rice salt-tolerant breeding. Nong Jia Can Mou 2017, 547, 97. [Google Scholar]
- Mardani, Z.; Rabiei, B.; Sabouri, H.; Sabouri, A.; Virk, P. Identification of molecular markers linked to salt-tolerant genes at germination stage of rice. Plant Breeding 2014, 133, 196–202. [Google Scholar] [CrossRef]
- Jing, W.; Deng, P.; Cao, C.; Zhang, W. Fine mapping of qSKC-1, a major quantitative trait locus for shoot K(+) concentration, in rice seedlings grown under salt stress. Breed. Sci. 2017, 67, 286–295. [Google Scholar] [CrossRef]
- He, Y.; Yang, B.; He, Y.; Zhan, C.; Cheng, Y.; Zhang, J.; Zhang, H.; Cheng, J.; Wang, Z. A quantitative trait locus, qSE3, promotes seed germination and seedling establishment under salinity stress in rice. Plant J. 2019, 97, 1089–1104. [Google Scholar] [CrossRef]
- Lin, H.X.; Zhu, M.Z.; Yano, M.; Gao, J.P.; Liang, Z.W.; Su, W.A.; Hu, X.H.; Ren, Z.H.; Chao, D.Y. QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance. Theor. Appl. Genet. 2004, 108, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cheng, J.; Chen, Z.; Huang, J.; Bao, Y.; Wang, J.; Zhang, H. Identification of QTLs with main, epistatic and QTL x environment interaction effects for salt tolerance in rice seedlings under different salinity conditions. Theor. Appl. Genet. 2012, 125, 807–815. [Google Scholar] [CrossRef]
- Bimpong, I.K.; Manneh, B.; Diop, B.; Ghislain, K.; Sow, A.; Amoah, N.K.A.; Gregorio, G.; Singh, R.K.; Ortiz, R.; Wopereis, M. New quantitative trait loci for enhancing adaptation to salinity in rice from Hasawi, a Saudi landrace into three African cultivars at the reproductive stage. Euphytica 2014, 200, 45–60. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.; Liu, X.; Du, H.; Dai, M.; Zhou, D.X.; Yang, W.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Zhang, W.H. Research Progress on Gene Mapping and Cloning for Salt Tolerance and Variety Improvement for Salt Tolerance by Molecular Marker-Assisted Selection in Rice. Chin. J. Rice Sci. 2017, 31, 111–123. [Google Scholar] [CrossRef]
- Thomson, M.J.; de Ocampo, M.; Egdane, J.; Rahman, M.A.; Sajise, A.G.; Adorada, D.L.; Tumimbang-Raiz, E.; Blumwald, E.; Seraj, Z.I.; Singh, R.K.; et al. Characterizing the Saltol Quantitative Trait Locus for Salinity Tolerance in Rice. Rice 2010, 3, 148–160. [Google Scholar] [CrossRef]
- Ammar, M.H.M.; Pandit, A.; Singh, R.K.; Sameena, S.; Chauhan, M.S.; Singh, A.K.; Sharma, P.C.; Gaikwad, K.; Sharma, T.R.; Mohapatra, T.; et al. Mapping of QTLs Controlling Na+, K+ and Cl- lon Concentrations in Salt Tolerance Indica Rice Variety CSR27. J. Plant Biochem. Biotechnol. 2009, 18, 139–150. [Google Scholar] [CrossRef]
- Liu, K.; Wang, L.; Xu, Y.; Chen, N.; Ma, Q.; Li, F.; Chong, K. Overexpression of OsCOIN, a putative cold inducible zinc finger protein, increased tolerance to chilling, salt and drought, and enhanced proline level in rice. Planta 2007, 226, 1007–1016. [Google Scholar] [CrossRef]
- Asano, T.; Hakata, M.; Nakamura, H.; Aoki, N.; Komatsu, S.; Ichikawa, H.; Hirochika, H.; Ohsugi, R. Functional characterisation of OsCPK21, a calcium-dependent protein kinase that confers salt tolerance in rice. Plant Mol. Biol. 2011, 75, 179–191. [Google Scholar] [CrossRef]
- Lee, S.C.; Han, S.K.; Kim, S.R. Salt- and ABA-inducible OsGASR1 is involved in salt tolerance. J. Plant Biol. 2015, 58, 96–101. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, F.; Zhou, Y. The application of multi-locus GWAS for the detection of salt-tolerance loci in rice. Front. Plant Sci. 2018, 9, 1464. [Google Scholar] [CrossRef]
- Hu, H.; You, J.; Fang, Y.; Zhu, X.; Qi, Z.; Xiong, L. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2008, 67, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Choudhury, A.R.; Gupta, B.; Gupta, S.; Sengupta, D.N. An ABRE-binding factor, OSBZ8, is highly expressed in salt tolerant cultivars than in salt sensitive cultivars of indica rice. BMC Plant Biol. 2006, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, H.; Maruyama, K.; Kidokoro, S.; Ito, Y.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; Nakashima, K. The abiotic stress-responsive NAC-type transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in rice. Mol. Genet. Genom. 2010, 284, 173–183. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, B.G.; Kwon, T.R.; Jeong, M.J.; Park, S.R.; Lee, J.W.; Byun, M.O.; Kwon, H.B.; Matthews, B.F.; Hong, C.B.; et al. Overexpression of the mitogen-activated protein kinase gene OsMAPK33 enhances sensitivity to salt stress in rice (Oryza sativa L.). J. Biosci. 2011, 36, 139–151. [Google Scholar] [CrossRef]
- Diedhiou, C.; Golldack, D. Salt-dependent regulation of chloride channel transcripts in rice. Plant Sci. 2006, 170, 793–800. [Google Scholar] [CrossRef]
- Wei, H.; Wang, X.; He, Y.; Xu, H.; Wang, L. Clock component OsPRR73 positively regulates rice salt tolerance by modulating OsHKT2;1-mediated sodium homeostasis. EMBO J. 2021, 40, e105086. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Kobayashi, N.I.; Yamaji, N.; Yamamoto, H.; Okubo, K.; Ueno, H.; Costa, A.; Tanoi, K.; Matsumura, H.; Fujii-Kashino, M.; Horiuchi, T.; et al. OsHKT1;5 mediates Na+ exclusion in the vasculature to protect leaf blades and reproductive tissues from salt toxicity in rice. Plant J. 2017, 91, 657–670. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, W.; Shu, F.; He, Q.; Zhang, L.; Yang, Z.; Peng, Z.; Xie, Y.; Deng, H. Comprehensive evaluation of salt-alkali tolerance of rice germplasms at germination and seedling stages and analysis of salt-tolerant genes. Chin. J. Biotechnol. 2022, 38, 252–263. [Google Scholar] [CrossRef]
- Wang, W. Using Indica-Japonica Cross RIL Population to Loucate QTLs Related to Salt and Alkali Tolerance in Rice. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2020. Chapter 3. pp. 18–38. [Google Scholar] [CrossRef]
- Qi, D.; Zhang, S.; Cao, G.; Ruan, R.; Sun, M.; Zhang, Y.; Zhou, Q.; Han, L. Studies on Screening Methods for Alkaline Tolerance at Germination Period and Early Seedling Stage in Rice. J. Plant Genetic Resour. 2006, 7, 74–80. [Google Scholar] [CrossRef]
- Schmidt, R.; Schippers, J.H.; Mieulet, D.; Obata, T.; Fernie, A.R.; Guiderdoni, E.; Mueller-Roeber, B. MULTIPASS, a rice R2R3-type MYB transcription factor, regulates adaptive growth by integrating multiple hormonal pathways. Plant J. 2013, 76, 258–273. [Google Scholar] [CrossRef]
- Bakade, R.; Ingole, K.D.; Deshpande, S.; Pal, G.; Patil, S.S.; Bhattacharjee, S.; Prasannakumar, M.K.; Ramu, V.S. Comparative transcriptome analysis of rice resistant and susceptible genotypes to Xanthomonas oryzae pv. oryzae identifies novel genes to control bacterial leaf blight. Mol. Biotechnol. 2021, 63, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guo, Z.; Lv, Y.; Cen, X.; Ding, X.; Wu, H.; Li, X.; Huang, J.; Xiong, L. Genetic control of the root system in rice under normal and drought stress conditions by genome-wide association study. PLoS Genet. 2017, 13, e1006889. [Google Scholar] [CrossRef] [PubMed]
- Baoxiang, W.; Yan, L.; Yifeng, W.; Jingfang, L.; Zhiguang, S.; Ming, C.; Yungao, X.; Bo, X.; Bo, Y.; Jian, L.; et al. OsbZIP72 is involved in transcriptional gene-regulation pathway of abscisic acid signal transduction by activating rice high-affinity potassium transporter OsHKT1;1. Rice Sci. 2021, 28, 257–267. [Google Scholar] [CrossRef]
- Jin, L.; Lu, Y.; Xiao, P.; Sun, M.; Corke, H.; Bao, J. Genetic diversity and population structure of a diverse set of rice germplasm for association mapping. Theor. Appl. Genet. 2010, 121, 475–487. [Google Scholar] [CrossRef]
- Ding, X.; Wang, L.; Zhang, Y.; Li, D.; Gao, Y.; Wei, W.; Wang, L.; Zhang, X. Genetic variation and associated mapping for traits related to plant height constitutions in core collections of sesame (Sesamum indicum L.). Chin. J. Oil Crop. Sci. 2013, 35, 262–270. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mendioro, M.S.; Diaz, G.Q.; Gregorio, G.B.; Singh, R.K. Mapping quantitative trait loci associated with yield and yield components under reproductive stage salinity stress in rice (Oryza sativa L.). J. Genet. 2013, 92, 433–443. [Google Scholar] [CrossRef]
- Lin, J.; Zhang, Y.; Chen, H.; Zhu, X.; Zhang, S.; Wang, Y.; Fang, X. Mapping QTLs for salt tolerance at seeding stage of landrace rice (Oryza sativa L.). Agric. Biotechnol. 2020, 9, 12–14, 25. [Google Scholar]
- Qiu, X.; Yuan, Z.; Liu, H.; Xiang, X.; Yang, L.; He, W.; Du, B.; Ye, G.; Xu, J.; Xing, D.; et al. Identification of salt tolerance-improving quantitative trait loci alleles from a salt-susceptible rice breeding line by introgression breeding. Plant Breed. 2015, 134, 653–660. [Google Scholar] [CrossRef]
- Chrispeels, H.E.; Oettinger, H.; Janvier, N.; Tague, B.W. AtZFP1, encoding Arabidopsis thaliana C2H2 zinc-finger protein 1, is expressed downstream of photomorphogenic activation. Plant Mol. Biol. 2000, 42, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wang, H.; Yin, X.; Wang, F.; Hu, S.; Liu, W.; Chen, L.; Dai, X.; Liang, M. Identification of Salt Tolerance Related Candidate Genes in ‘Sea Rice 86’ at the Seedling and Reproductive Stages Using QTL-Seq and BSA-Seq. Genes 2023, 14, 458. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, J.; Suo, Y.; Liu, H.; Wang, J.; Zheng, H.; Sun, X.; Zou, D. QTL Mapping and QTL × Environment Interaction Analysis of Salt and Alkali Tolerance-Related Traits in Rice (Oryza sativa L.). Scientia Agricultura Sinica 2017, 50, 1747–1762. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Sang, T.; Lu, B.R.; Hong, D.Y. Phylogeny of rice genomes with emphasis on origins of allotetraploid species. Proc. Natl. Acad. Sci. USA 1999, 96, 14400–14405. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhang, M.; Niu, X.; Wang, C.; Xu, Q.; Feng, Y.; Wang, S.; Yuan, X.; Yu, H.; Wang, Y.; et al. Uncovering novel loci for mesocotyl elongation and shoot length in indica rice through genome-wide association mapping. Planta 2016, 243, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Mather, K.A.; Caicedo, A.L.; Polato, N.R.; Olsen, K.M.; McCouch, S.; Purugganan, M.D. The extent of linkage disequilibrium in rice (Oryza sativa L.). Genetics 2007, 177, 2223–2232. [Google Scholar] [CrossRef]
- Zhang, L.S.; Ma, C.R.; Ji, Q.; Wang, Y.F. Genome-wide identification, classification and expression analyses of SET domain gene family in Arabidopsis and rice. Yi Chuan 2009, 31, 186–198. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Bland, J.M.; Altman, D.G. Multiple significance tests: The Bonferroni method. BMJ 1995, 310, 170. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, Z.L.; Zou, X.; Huang, J.; Ruas, P.; Thompson, D.; Shen, Q.J. Annotations and functional analyses of the rice WRKY gene superfamily reveal positive and negative regulators of abscisic acid signaling in aleurone cells. Plant Physiol. 2005, 137, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Peng, Y.; Guo, Z. Constitutive expression of pathogen-inducible OsWRKY31 enhances disease resistance and affects root growth and auxin response in transgenic rice plants. Cell Res. 2008, 18, 508–521. [Google Scholar] [CrossRef]
- Giri, J.; Vij, S.; Dansana, P.K.; Tyagi, A.K. Rice A20/AN1 zinc-finger containing stress-associated proteins (SAP1/11) and a receptor-like cytoplasmic kinase (OsRLCK253) interact via A20 zinc-finger and confer abiotic stress tolerance in transgenic Arabidopsis plants. New Phytol. 2011, 191, 721–732. [Google Scholar] [CrossRef]
- Wang, R.; Jing, W.; Xiao, L.; Jin, Y.; Shen, L.; Zhang, W. The Rice High-Affinity Potassium Transporter1;1 Is Involved in Salt Tolerance and Regulated by an MYB-Type Transcription Factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014, 84, 19–36. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | Year | Mean ± SE | Range | CV | HB2 |
|---|---|---|---|---|---|
| GP | 2020 | 0.55 ± 0.024 | 0.00~1.0 | 57.18% | 83.89% |
| 2021 | 0.56 ± 0.019 | 0.02~0.97 | 44.14% | ||
| GR | 2020 | 0.91 ± 0.47 | 0.10~1.00 | 15.27% | 52.27% |
| 2021 | 0.93 ± 0.072 | 0.33~1.00 | 10.26% | ||
| SL | 2020 | 3.32 ± 0.11 | 0.69~8.22 | 43.39% | 85.44% |
| 2021 | 4.00 ± 0.12 | 1.03~8.77 | 39.41% | ||
| RL | 2020 | 1.87 ± 0.16 | 0.00~8.63 | 80.50% | 89.05% |
| 2021 | 4.08 ± 0.22 | 0.36~11.03 | 70.57% | ||
| GPR | 2020 | 0.41 ± 0.025 | −0.04~1.00 | 78.41% | 58.61% |
| 2021 | 0.39 ± 0.020 | −0.30~0.98 | 67.72% | ||
| GRR | 2020 | 0.07 ± 0.01 | −0.13~0.90 | 117.30% | 68.35% |
| 2021 | 0.01 ± 0.01 | −0.57~0.44 | 124.30% | ||
| SLR | 2020 | 0.61 ± 0.01 | 0.21~0.93 | 26.87% | 66.47% |
| 2021 | 0.53 ± 0.01 | 0.07~0.83 | 29.52% | ||
| RLR | 2020 | 0.78 ± 0.02 | −0.15~1.00 | 35.18% | 80.51% |
| 2021 | 0.61 ± 0.02 | −0.65~0.97 | 49.05% | ||
| RSD | 2020 | 0.24 ± 0.02 | −0.03~0.95 | 81.19% | 46.26% |
| 2021 | 0.21 ± 0.01 | −0.14~0.73 | 67.60% | ||
| CRS | 2020 | 0.7 ± 0.02 | 0.07~0.96 | 29.32% | 66.87% |
| 2021 | 0.56 ± 0.02 | −0.20~0.88 | 37.29% |
| Traits | GP | GPR | GR | GRR | SL | SLR | RL | RLR | RSD | CRS |
|---|---|---|---|---|---|---|---|---|---|---|
| GP | 1 | |||||||||
| GPR | −0.840 ** | 1 | ||||||||
| GR | 0.366 ** | −0.309 ** | 1 | |||||||
| GRR | −0.261 * | 0.321 ** | −0.696 ** | 1 | ||||||
| SL | 0.004 | −0.023 | 0.201 | −0.179 | 1 | |||||
| SLR | −0.008 | 0.050 | −0.240 | 0.219 | −0.841 ** | 1 | ||||
| RL | −0.166 | 0.180 | 0.105 | −0.143 | 0.740 ** | −0.722 ** | 1 | |||
| RLR | 0.188 | −0.182 | −0.057 | 0.145 | −0.648 ** | 0.662 ** | −0.887 ** | 1 | ||
| RSD | −0.818 ** | 0.868 ** | −0.570 ** | 0.538 ** | −0.065 | 0.079 | 0.095 | −0.105 | 1 | |
| CRS | 0.129 | −0.110 | −0.121 | 0.187 | −0.777 ** | 0.836 ** | −0.901 ** | 0.961 ** | −0.043 | 1 |
| Trait | QTLs | Chr. | Position | p Value | R2 (%) | Year | Cloned Genes | Reference |
|---|---|---|---|---|---|---|---|---|
| GPR | qGPR1-1 | 1 | 38,365,940 | 1.07 × 10−4 | 11.3 | 2020 | ||
| qGPR1-2 | 1 | 38,309,966 | 3.33 × 10−5 | 12.8 | 2020 | |||
| qGPR2-1 | 2 | 24,709,239 | 7.28 × 10−5 | 11.8 | 2020 | |||
| 24,709,239 | 2.04 × 10−4 | 10.4 | 2021 | |||||
| qGPR2-2 | 2 | 35,432,245 | 8.33 × 10−5 | 11.6 | 2021 | OsMPS | Schmidt et al. [33] | |
| qGPR6 | 6 | 7,793,840 | 1.58 × 10−4 | 10.8 | 2021 | |||
| qGPR7 | 7 | 3,448,084 | 2.09 × 10−4 | 10.4 | 2021 | |||
| SLR | qSLR2 | 2 | 9,878,862 | 5.04 × 10−5 | 10.1 | 2020 | ||
| qSLR9 | 9 | 17,238,271 | 4.58 × 10−5 | 10.2 | 2020 | |||
| 17,238,271 | 6.99 × 10−5 | 11.6 | 2021 | |||||
| RSD | qRSD1 | 1 | 38,309,966 | 6.36 × 10−5 | 12.0 | 2020 | ||
| qRSD2 | 2 | 4,515,468 | 6.03 × 10−5 | 11.7 | 2021 | |||
| qRSD3 | 3 | 5,919,379 | 3.82 × 10−5 | 12.6 | 2020 | |||
| qRSD12 | 12 | 23,873,291 | 5.53 × 10−5 | 11.9 | 2021 |
| QTL | Chr. | Gene ID | MSU ID | Feature Notes |
|---|---|---|---|---|
| 2 | Os02g0619600 | LOC_Os02g40664 | zinc finger family protein, putative, expressed | |
| 2 | Os02g0620100 | LOC_Os02g40680 | mis12 protein, expressed | |
| 2 | Os02g0621100 | LOC_Os02g40770 | SET domain containing protein, expressed | |
| qGPR2 | 2 | Os02g0621300 | LOC_Os02g40784 | WAX2, putative, expressed |
| 2 | Os02g0621500 | LOC_Os02g40810 | Zinc finger, ZZ type domain containing protein, expressed | |
| 2 | Os02g0622100 | LOC_Os02g40860 | CK1_CaseinKinase_1.5—CK1 includes the casein kinase 1 kinases, expressed | |
| 2 | Os02g0622400 | LOC_Os02g40890 | GLTP domain containing protein, putative, expressed | |
| 2 | Os02g0622500 | LOC_Os02g40900 | RNA recognition motif containing protein, putative, expressed | |
| 9 | Os09g0456100 | LOC_Os09g28300 | remorin C-terminal domain containing protein, putative, expressed | |
| 9 | Os09g0456200 | LOC_Os09g28310 | bZIP transcription factor, putative, expressed | |
| 9 | Os09g0456700 | LOC_Os09g28340 | expressed protein | |
| qSLR9 | 9 | Os09g0456900 | LOC_Os09g28370 | retrotransposon protein, putative, unclassified, expressed |
| 9 | Os09g0458000 | LOC_Os09g28450 | paramyosin, putative, expressed | |
| 9 | Os09g0458100 | LOC_Os09g28460 | xyloglucan fucosyltransferase, putative, expressed | |
| 9 | Os09g0458400 | LOC_Os09g28480 | expressed protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Zhou, Z.; Xiong, M.; Du, M.; Lin, X.; Liu, C.; Lu, M.; Liu, Z.; Chang, Y.; Liu, E. Mining Salt Tolerance SNP Loci and Prediction of Candidate Genes in the Rice Bud Stage by Genome-Wide Association Analysis. Plants 2023, 12, 2163. https://doi.org/10.3390/plants12112163
Wang R, Zhou Z, Xiong M, Du M, Lin X, Liu C, Lu M, Liu Z, Chang Y, Liu E. Mining Salt Tolerance SNP Loci and Prediction of Candidate Genes in the Rice Bud Stage by Genome-Wide Association Analysis. Plants. 2023; 12(11):2163. https://doi.org/10.3390/plants12112163
Chicago/Turabian StyleWang, Rui, Zhenzhen Zhou, Mengyuan Xiong, Mingyu Du, Xingxing Lin, Cuiping Liu, Mingwei Lu, Zhengbo Liu, Yinping Chang, and Erbao Liu. 2023. "Mining Salt Tolerance SNP Loci and Prediction of Candidate Genes in the Rice Bud Stage by Genome-Wide Association Analysis" Plants 12, no. 11: 2163. https://doi.org/10.3390/plants12112163